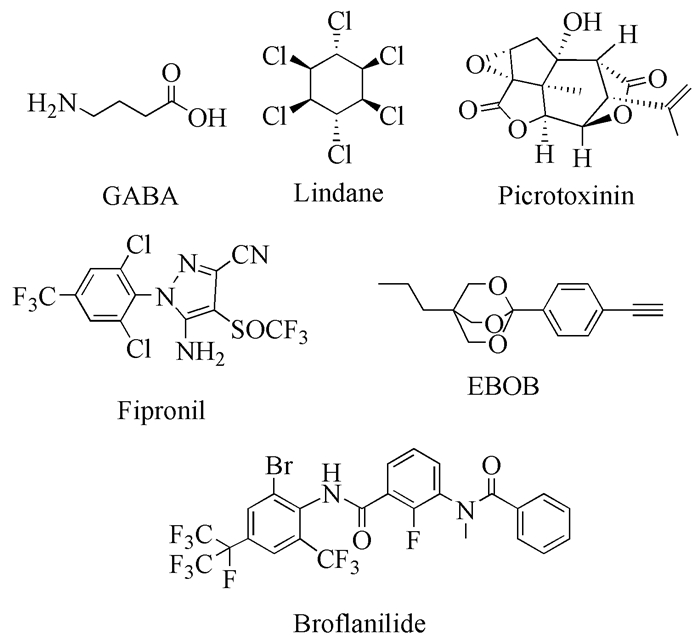
图式 1.
GABA与典型的GABA受体非竞争性拮抗剂
Scheme 1.
GABA and representative NCAs of GABA receptors

γ-氨基丁酸(γ-aminobutyric acid,GABA)是动物体内一类重要的神经递质,主要存在于中枢神经系统中,由谷氨酸经体内酶代谢转化而来[1~4]。作为一种抑制性神经递质,GABA由神经细胞突触前膜释放,经突触间隙到达突触后膜,并与后膜上的GABA受体发生特异性结合,促使细胞膜上的氯离子通道开放,膜外的氯离子进入细胞内,产生超极化现象,从而抑制神经细胞兴奋,发挥生理作用。
GABA门控氯离子通道作为一种离子型GABA受体(若无特殊说明,本文中涉及的GABA受体均为离子型GABA受体)是一类重要的杀虫剂靶标[4]。该离子通道由5个亚基组成,每个亚基主要由一个较长胞外结构域和4个跨膜域(Transmembrane Domain,TMD)构成,每个亚基的第二个跨膜区(TMD2)构成氯离子通道的内壁[5]。研究表明,GABA受体拮抗剂在哺乳动物和昆虫之间具有选择性,这为设计和开发安全、高效的杀虫剂提供了理论依据[6]。该受体拮抗剂又分为非竞争性拮抗剂(Noncompetitive antagonist,NCA)和竞争性拮抗剂(Competitive antagonist,CA)两类。这两类拮抗剂的主要区别在于其结合位点的不同,从而导致调节受体的功能有一定差异[3]。
目前已上市靶向GABA受体的农用杀虫剂主要为该类受体的NCA。传统的GABA受体NCA(图式 1)主要有多氯环烷烃类(如Lindane)、内酯环己醇类(如Picrotoxinin)、芳基吡唑类(如Fipronil)和双环磷酸酯类(如EBOB)等[7]。一般来说,NCA直接作用GABA受体的氯离子通道,非竞争性地阻滞神经细胞氯离子的内流,干扰昆虫中枢神经系统的正常功能,使神经兴奋不能够正常的抑制,从而导致昆虫死亡。传统NCA主要作用于受体的TMD2跨膜段,通过变构调节的作用方式阻碍氯离子通道的开启[7, 67];CA则作用于受体细胞膜外区域的GABA识别位点。两者的作用机制和作用位点有明显的区别[4]。
传统NCA类杀虫剂因毒性富集及对环境和其他生物的潜在隐患[5, 8],大部分已禁止在农业中使用[9, 10]。针对传统NCA的缺点和不足,近年来研究人员开发了两类新型GABA受体NCA:异噁唑啉类[11](Isoxazolines,如氟雷拉纳)和间二酰胺类[8](meta-Diamides,如Broflanilide,图式 1)。研究表明,这两类新型NCA的作用位点和机制有别于传统的NCA。此外,研究发现GABA受体的CA也具有一定的杀虫活性[11~15],但目前其作为杀虫剂的研究仍处于起步阶段。本文主要对近年来作用于GABA受体的新型异噁唑啉类杀寄生虫剂的研究进展进行综述,为这类杀虫剂的研发提供一定参考和思路。
杂环化合物是农药合成和研发过程中的重要中间体或化学实体[16~18]。噁唑或异噁唑是一类有价值的杂环化合物,具有独特的化学结构和生物活性,其衍生物很多被开发为农化产品[16]。异噁唑啉类化合物是一类重要的五元杂环化合物,从结构上异噁唑啉主要分为:2-异噁唑啉和4-异噁唑啉(图式 2);其中,4-异噁唑啉的稳定性较差,因此文献中无特殊说明,异噁唑啉一般指2-异噁唑啉。异噁唑啉类化合物不仅在有机合成领域占据比较重要的地位,同时在消炎[18]、抗癌[19]、杀菌及除草[24, 25]等方面也有很好的应用价值,部分异噁唑啉类化合物已被开发成为农药[21~23]。
在研究邻甲酰氨基苯甲酰胺的过程中,研究者意外发现了一类具有杀虫活性的异噁唑啉类化合物[26]。其中,氟雷拉纳(Fluralaner)是最具代表性的化合物,其可通过阻断GABA和谷氨酸门控氯离子通道产生杀虫或杀螨活性[27]。氟雷拉纳化学名称为4-[5-(3, 5-二氯苯基)-5-(三氟甲基)-4, 5-二氢-异噁唑-3-基]-2-甲基-N-[2-氧代-2-(2, 2, 2-三氟乙基氨基)乙基]-苯甲酰胺(图式 3),研发代号为A1443[28],具有一对对映异构体[29, 30]。研究表明,其S构型(A664)杀虫活性远高于R构型(A663)[31, 32]。
2004年,日本日产化学工业株式会社Mita等首先发现了广谱杀虫剂氟雷拉纳,于2005年获国际专利[11]。2010年,Ozoe等[45]报道了氟雷拉纳的对昆虫GABA受体的离体电生理活性及其杀寄生虫活性。2013年,Garcia-Reynaga等[5, 8, 11, 30]研究了氟雷拉纳及异噁唑啉类衍生物在GABA受体的作用位点和机制。2014年,美国FDA批准氟雷拉纳口服咀嚼片(商品名:Bravector)上市,主要用来治疗猫犬等动物体外寄生虫[33~40]。2017年11月,氟雷拉纳外用滴剂(商品名:一锭除)在中国台湾上市销售[41]。2019年7月,默沙东动物保健宣布,其犬类抗寄生虫口服药物贝卫多(氟雷拉纳咀嚼片)获批在中国上市[42],主要用于治疗犬体表的跳蚤和蜱虫等感染,还可辅助治疗因跳蚤引起的过敏性皮炎[29, 43, 44]。
氟雷拉纳对犬蚤(Ctrnocephalides canis)、猫蚤(Ctenocephalides felis)有驱杀作用[38, 45~47],对成年跳蚤起效快,还可阻止跳蚤产卵,从而破坏跳蚤的生命周期。此外,氟雷拉纳还具有广谱的杀蜱活性,对篦子硬蜱(Ixodes ricinus)、六角硬蜱(Ixodes hexagonus)、肩突硬蜱(Ixodes scapularis)、全环硬蜱(Ixodes holocylus)等有较理想的灭杀作用[48~51]。与传统的杀寄生虫剂相比,氟雷拉纳具有杀虫速度快[52, 53]、药效时间长[53]和毒副作用小[54]等特点。作为异噁唑啉类体外抗寄生虫剂的首款药剂,氟雷拉纳对跳蚤和螨虫的药效可持续7个月,能达到治疗与长期预防的目的[54, 55]。此外,氟雷拉纳对蚊虫的杀灭活性也引起了研发人员的兴趣,Jiang等[56]报道了氟雷拉纳对黑斑蚊(Aedes aegypti)幼蚊的杀灭活性是氟虫腈的10~13倍,但对成年蚊的效果低于氟虫腈,这也为预防蚊虫提供了新的思路。
Sheng等[29, 51, 57, 59]发现,氟雷拉纳对斜纹夜蛾(Spodoptera litura)、二化螟(Chilo suppressalis)、二斑叶螨(Tetranychus urticae)、灰飞虱(Laodelphax striatellus)等常见的农业害虫有明显的杀虫作用。与传统NCA类杀虫剂氟虫腈相比,氟雷拉纳表现出高效的杀虫效果且不存在交互抗性[46]。Leviticus等[60]比较了氟雷拉纳和三种主要的杀螨剂对二斑叶螨的杀虫活性,结果表明氟雷拉纳的活性仅低于阿维菌素,但远高于哒螨灵和噻螨酮[44, 61, 62]。Sun等[58]发现氟雷拉纳对小菜蛾(Pultella xylostella)和黏虫(mythimna seperata)均具有较高的杀虫活性,且优于传统药剂四氯虫酰胺。虽然目前氟雷拉纳对农业害虫的杀虫活性均来自实验室研究,但特殊的作用靶点和高效的杀虫活性使其在农用方面仍具有很好的潜在应用价值,有望成为新型农业杀虫剂。
氟雷拉纳作为昆虫GABA受体的新型NCA,其主要作用机理是通过减弱昆虫神经突触后膜的GABA门控氯离子通道的通透性,快速介导抑制突触传递,导致昆虫的死亡[37, 63~65]。Gassel等[28, 30]发现,氟虫腈对昆虫GABA受体亲和力比氟雷拉纳低,且氟雷拉纳与苯基吡唑类、有机氯类等传统杀虫剂的作用位点不同。Ozoe等[28, 29]发现,氟雷拉纳可与二化螟突变型(A2'S)GABA受体结合,而该突变位点能够影响狄氏剂等传统NCA的抑制作用,这表明氟雷拉纳的作用位点可能有别于传统NCA的TMD2中的2'位点。氟雷拉纳还可作用于谷氨酸门控氯离子通道[31],Nakata等[68]研究发现,当家蝇(Musca domestica)谷氨酸门控氯离子通道TMD3的315位亮氨酸突变为苯丙氨酸时,氟雷拉纳对该通道的活性显著提高,这一发现可为研发多靶标杀寄生虫剂提供新的思路和参考。氟雷拉纳与GABA受体的相互作用的模式尚不十分明确,仍需进一步研究。
Pfister等[70]对氟雷拉纳的体内药代动力学研究发现,其主要通过肝脏代谢排出,几乎不经过肾脏代谢,安全性较好,对肝脏无损害。Kilp等[69]发现犬在口服氟雷拉纳后可快速吸收,24h内可以达到最大血浆浓度,且在血浆中能够存在12周。犬对外用的氟雷拉纳吸收缓慢,这可能与其在生物体内的代谢方式有关。
在治疗跳蚤、蜱虫和螨虫等动物常见的寄生虫的同时,氟雷拉纳对哺乳动物如大鼠[71, 72]、猫[73]、犬[74, 75]等和禽类如鸟[76]均未表现出明显的毒副作用,且还能治疗寄生虫引起的皮肤病。Huang等[77, 78]研究证实氟雷拉纳对蜜蜂毒性较高(LD50为0.13μg/头),这表明氟雷拉纳在需要蜜蜂等传粉媒介的农业种植应用领域会受到一定限制[79],但是对自花授粉类作物无较大影响[80]。Jia等[81]研究了氟雷拉纳对水生生物的毒性,结果表明,其对鲤鱼(Cyprinus carpio)和斑马鱼(Danio rerio)毒性较低,而对溞类(Daphnia magna)等毒性很高。针对其他水生生物的研究报道较少,因此在水产养殖行业应慎用氟雷拉纳。2017年,美国FDA发现含氟雷拉纳成分的口服药可能会有导致犬类神经失调的副作用[82, 86, 87]。
阿福拉纳(Afoxolaner)化学名为4-[5-[3-氯-5-(三氟甲基)苯基]-4, 5-二氢-5-(三氟甲基)-3-异噁唑基]-N-[2-氧代-2-(2, 2, 2-三氟乙基)氨基]-乙基]-1-萘甲酰胺(图式 4),由法国梅里亚公司于2014年研发,是一种新型的动物体外抗寄生虫剂,具有起效快速、药效时间长、安全性高、口服方便等特点[90~92, 95],能够治疗猫蚤等寄生虫,并且可以预防跳蚤和蜱虫再次寄生[1, 91~94, 99]。2017年8月,勃林格殷格翰公司研发的犬用口服驱虫药尼可信[88, 89](NexGard,通用名:阿福拉纳咀嚼片)在中国上市;2018年8月,一种广谱杀虫剂组合咀嚼片超可信(NexGard Spectra)在中国上市[88],其主要成分中包含阿福拉纳和米尔贝肟。米尔贝肟是一种有效治疗多种犬消化道线虫和预防心丝虫感染的大环内脂类药物[91, 96~98],该药剂不仅可以治疗跳蚤、蜱虫等体外寄生虫,同样也可以驱除和预防心丝虫等体内常见寄生虫[45, 83~85, 88, 96]。
Lahm等[100]发现异噁唑啉4位的三氟甲基与3,5-二取代的芳基和羧酰胺基是该类化合物产生活性的药效基团,于是进一步设计并合成了4-偶氮苯基异噁唑啉类衍生物,代表化合物为CPD I(图式 4),此化合物具有较高杀虫活性,而阿福拉纳是在CPD I的基础上进行结构修饰而来。Shoop等[91, 83, 99]发现,CPD I能有效抑制美洲大蠊(Periplaneta americana)神经元中GABA引起的电流,IC50值为10.8nmol/L,但对烟碱乙酰胆碱受体无显著活性[91],与阿福拉纳测试结果相似。阿福拉纳对在非洲爪蟾卵母细胞中表达的果蝇(Drosophila melanogaster)野生型GABA受体(RDL[66],Resistance to Dieldrin)的IC50值为3.7nmol/L。Cordova等[100, 101]进一步测试了阿福拉纳对果蝇野生型和A302S突变GABA受体[8, 71, 102]的活性,结果表明,果蝇A302S突变GABA受体对狄氏剂和氟虫腈表现出较强或中等强度的抗性,而对阿福拉纳无明显抗性,故阿福拉纳可能是以不同于狄氏剂和氟虫腈的结合模式与昆虫GABA受体发生相互作用[91]。
Shoop等[91, 125]研究表明,阿福拉纳可与血浆蛋白高度结合,在吸血节肢动物的进食过程中可被吸收,因此对跳蚤等有很强的抑制作用。Letendre等[91, 103, 104]确定犬类口服剂量为2.5mg/kg时,阿福拉纳血浆浓度可达到最大。犬在使用阿福拉纳2h后开始杀灭跳蚤[90, 91, 93, 94],8h后,杀虫效率可达到100%,且对猫蚤的治疗效果相似[89, 105~107]。Dryden等[108, 109]比较了阿福拉纳和氟雷拉纳在控制跳蚤数量、减少瘙痒和减少自然感染的犬的皮肤病变方面的活性,发现两者之间的差距较小[93, 106]。在跳蚤繁殖试验中发现,阿福拉纳能够导致跳蚤性别发生转变,因此可以抑制跳蚤的繁殖,减少寄主体表跳蚤的数量[108~110]。阿福拉纳可驱杀多种常见蜱虫[84, 92, 95, 96],如美洲犬蜱(Dermacentor variabilis)、篦子硬蜱和网纹革蜱(Dermacentor reticulatus)等[153, 155, 158, 162, 163],并可在24h内杀灭90%以上感染的蜱虫,杀虫效果持续1个月以上[94, 104, 97]。Beugnet等[45, 93, 105, 106]比较了阿福拉纳与吡虫啉/莫西丁对犬体外杀螨的疗效,结果表明,阿福拉纳螨虫数减少率均高于传统药物吡虫啉/莫西丁[106]。阿福拉纳对蠕形螨(Demodex)清除率可达到99%以上[92, 110, 111],对耳螨(Otodectes cynotis)的清除率达98.5%[112];阿福拉纳对其他螨虫如犬疥螨(Sarcoptes scabiei)、毛囊虫(Demodicosis)等同样具有良好的治疗效果[91, 99, 113]。相比其他异噁唑啉杀虫剂,阿福拉纳还对黑斑蚊、锥蝽(Triatoma infestans)有杀灭作用[99, 105, 114]。基于阿福拉纳公认的安全性和功效特性以及高度可接受的口服剂型,其已被全球50多个国家的兽医和宠物主人广泛接受,并为数百万只犬提供了保护。
药代动力学研究表明,阿福拉纳分子量低且具有疏水性,LogP值为5.5,在动物生理pH范围内可保持电中性[97, 102~104];同时,阿福拉纳在体内可通过被动扩散穿过细胞膜。Carithers等[94, 104, 112]测试了尼可信和超可信两种制剂,在给药后2~4 h内,阿福拉纳的血浆最高浓度分别可达到1655ng/mL和1822ng/mL;尼可信阿福拉纳和超可信阿福拉纳的平均生物利用度分别为73.9%和88.3%。阿福拉纳在成年犬体内的半衰期为12.8d左右,在肝脏通过细胞色素P450酶形成的氧化产物而代谢清除,但清除率较低[115, 116]。研究表明,阿福拉纳对昆虫GABA受体的选择性较高[29, 117],对哺乳动物较安全,因此阿福拉纳在正常给药剂量下对动物安全有效[118, 119]。
2009年,Zoetis公司从3000多种异噁唑啉类化合物中筛选出了一种新型的口服、外源性广谱杀寄生虫剂—沙罗拉纳(Sarolaner,图式 5)[120, 121],其化学名称为1-[5'-[(5S)-5-(3,5-二氯-4-氟苯基)]-5-(三氟甲基)-4, 5-二氢异噁唑-3-基]-3'-H-螺-[(氮杂环丁烷)-3, 1'-(2)-苯并呋喃-1-基]-2-(甲基磺酰基)-乙酮,研发代号为PF-6450567[122, 123]。沙罗拉纳主要用于犬蜱虫、螨虫的预防和治疗(商品名:SimparicaTM)。与其他几类异噁唑啉杀虫剂相比,该化合物的化学结构有明显的区别,含有一个独特的螺氮化二苯并呋喃的结构,这种结构在传统的杀寄生虫剂中未见报道;同时分子末端存在磺酸甲酯结构,增加了分子的极性表面积。这些结构可以增加药物在体内的药代动力学特性,达到快速杀灭寄生虫的目的[120~123]。

沙罗拉纳对跳蚤的LC80值为0.1μg/mL,对蜱虫的LC100值为0.003μg/mL[174],与阿福拉纳和氟雷拉纳等相比,效果更好[122~125]。沙罗拉纳是一种广谱杀寄生虫剂,可防治多种蜱虫(表 1)。沙罗拉纳对犬类的最佳口服剂量为2.5mg/kg,可在3~4 h内发挥药效,并持续35d以上[119, 124, 126~130]。
 下载:
导出CSV
下载:
导出CSV
| 蜱虫种类 | 拉丁学名 | 文献 |
| 肩突硬蜱 | Ixodes scapularis | [132] |
| 篦子硬蜱 | Ixodes ricinus | [119, 120, 124] |
| 六角硬蜱 | Ixodes hexagonus | [119, 133] |
| 全沟硬蜱 | Ixodes pesuleatus | [132] |
| 全环硬蜱 | Ixodes holocyclus | [119] |
| 网纹革蜱 | Dermacentor reticulatus | [119, 126, 137] |
| 斑点钝眼蜱 | Amblyomma maculatum | [131] |
| 美洲钝眼蜱 | Amblyomma americanum | [134] |
| 变异革蜱 | Dermacentor variabilis | [124] |
| 血红扇头蜱 | Rhipicephalus sanguineus | [119, 124] |
Geurden等[119, 124, 131]测试了沙罗拉纳对常见的四种蜱虫(网纹革蜱、多毛硬蜱、篦子硬蜱、血红扇头蜱)的杀虫活性,结果表明杀虫率均大于99%,且此疗效均优于多杀霉素和氟虫腈,同时可大幅度防止再次遭受感染。沙罗拉纳和Advantix(吡虫啉和氯菊酯)对网纹革蜱的杀虫速度相比较,沙罗拉纳在服用后可以快速杀死所有感染的蜱虫,而Advantix的疗效较慢[132~135]。对于螨虫的治疗,口服剂量为2.0mg/kg时,蠕形螨和耳螨的数量均可减少98%以上,同时螨虫引起的临床症状也有所改善。因此,沙罗拉纳可以作为全身性杀螨剂,预防由莱姆病螺旋体(Borrelia burgdorferi)、犬埃里希氏菌(Ehrlichia canis)、犬巴贝斯虫(Babesia canis)等病原体传播的疾病[122, 123, 127, 135]。另一方面,Yonetake等[136]将沙罗拉纳与塞拉霉素联用,所开发的外用制剂可用于预防猫的心脏蛔虫疾病,这为治疗寄生虫感染提供了新思路。
电生理研究证实,沙罗拉纳可抑制猫蚤CfRDL-A285和CfRDL-S285受体的GABA诱导电流,IC50值分别为135和136 nmol/L,阿福拉纳对这两种受体的IC50值分别为539和412 nmol/L,而狄氏剂对CfRDL-S285受体(IC50 > 3300nmol/L)显示出较弱的拮抗作用,这表明沙罗拉纳对猫蚤GABA受体的抑制作用不受耐药基因突变的影响,其结合方式与传统抑制剂可能不同。此外,与人GABA受体相比,沙罗拉纳对跳蚤等昆虫GABA受体的选择性更强(约244倍)[120],故沙罗拉纳对哺乳动物相对安全无毒。
Cherni等[136, 137]对美国不同地区的479只犬在服用沙罗拉纳后进行了安全性评估。大多数犬观察到的临床症状与过敏和皮炎相一致,或为一般犬类中常见的偶发症状。经沙罗拉纳治疗的犬出现的异常临床症状主要包括红斑(3.5%)、瘙痒(3.5%)、呕吐(3.2%)、耳漏(2.5%)、嗜睡(2.5%)和咳嗽(2.2%)等;而接受多杀霉素治疗的犬中,这些症状的发生率更高。为确定沙罗拉纳与其他药物间的耐受性,研究人员在犬接受其他药物治疗(如狂犬病疫苗等)后让其服用沙罗拉纳,结果表明沙罗拉纳对这些药物均表现出较好的耐受性,无其他严重副作用[118, 138],因此宠物使用其他药物的情况下仍可服用沙罗拉纳。
新型杀寄生虫剂洛替拉纳(Lotilaner)(商品名:CredelioTM,图式 5)由礼来公司于2017年开发[7, 138],用于快速治疗犬蜱和跳蚤的感染,其化学名为(S)-5-[5-(3, 4, 5-三氯苯基)-5-三氟甲基-4, 5-二氢-异噁唑-3-基]-3-甲基-噻吩-2-羧酸-[2, 2, 2-三氟-乙基-氨基甲酰基)-甲基]-酰胺。与其他异噁唑啉类杀寄生虫剂相同,洛替拉纳具有生物活性的结构是S-对映体,其R-对映体(AHC-222954)无活性。给猫服用洛替拉纳纯对映体后,洛替拉纳在体内未发生外消旋化,因此R-对映体的药代动力学和安全性无实际研究意义[139]。目前为止,洛替拉纳是对猫最有效的口服异噁唑啉类抗蜱虫药剂,且安全性优于传统体外杀寄生虫类药物[140, 141]。
洛替拉纳咀嚼片口服最低剂量为6.0mg/kg时,在给药后2h开始杀死跳蚤和蜱虫,并于24h内达到100%的疗效,同时能在35d内迅速杀死新感染的寄生虫[7, 144, 145],并且可以预防宠物再次被感染[138, 141~143]。Otaki等[146~147]比较了犬在服用洛替拉纳和阿福拉纳后对长角血蜱(Haemaphysalis longicornis)的疗效,治疗48h后,洛替拉纳的疗效大于95%且可持续37d,而阿福拉纳疗效可达到100%,但30d后疗效逐渐下降到91.9%。此外,洛替拉纳可以高效的消灭寄生在犬身上的篦子硬蜱、网纹革蜱、血红扇头蜱和钝眼蜱等[139, 143, 210~222],且在一个月内,对这些蜱虫的杀灭效果依旧显著[140, 147~149]。
为研究洛替拉纳与GABA受体的作用机理,Rufener等测试了洛替拉纳对黑腹果蝇野生型(DmS-GABAR,下同为GABA受体缩写形式)、黑腹果蝇狄氏剂/氟虫腈耐药型(DmR2-GABAR)、鲑鱼虱野生型(Lepeophtheirus salmonis, Ls-GABAR)、小扇头蜱野生型(Rhipicephalus microplus,Rm-GABAR)和犬野生型(Canis lupus familiaris,α1β2γ2 GABAAR)等5种GABA受体电生理活性。洛替拉纳对DmS-GABAR和DmR2-GABAR的IC50值分别为23.84和38.25 nmol/L(表 2),无显著差异;另一方面,DmR2-GABAR对狄氏剂、氟虫腈和洛替拉纳的耐药因子分别为12000、8.35和1.60,因此洛替拉纳与狄氏剂或氟虫腈之间不存在交叉抗性。这一现象和其他异噁唑啉杀寄生虫剂的结果相一致[7, 145, 151, 152],原因可能是异噁唑啉类化合物与GABA门控氯离子通道中独特的结合口袋结合而发挥抑制作用。
下载:
导出CSV
| GABA受体类型 | 拮抗剂 | IC50/(nmol/L) |
| DmR2-GABAR | Dieldrin | > 10, 000 |
| DmR2-GABAR | Fipronil | 230.32 |
| DmR2-GABAR | Lotilaner | 38.25 |
| DmS-GABAR | AHC-2229544 | 959.47 |
| DmS-GABAR | Dieldrin | 4170 |
| DmS-GABAR | Fipronil | 27.58 |
| DmS-GABAR | Lotilaner | 23.84 |
| Ls-GABAR | Fipronil | 164.85 |
| Ls-GABAR | Lotilaner | 52.40 |
| Rm-GABAR | Fipronil | 25.56 |
| Rm-GABAR | Lotilaner | 36.79 |
| α1β2γ2GABAAR | Lotilaner | > 10, 000 |
洛替拉纳在最高测试浓度(10μmol/L)下对犬野生型α1β2γ2 GABAR无明显抑制作用,这表明洛替拉纳对对宠物毒性较低或基本无毒害作用。洛替拉纳是Ls-GABAR强效拮抗剂,IC50值比氟虫腈高(约为3倍),对Rm-GABAR同样具有很强的拮抗作用。洛替拉纳对鱼类、甲壳类动物和其他哺乳动物是否有毒害作用需进一步研究证实[138, 139]。
洛替拉纳分子量为596.76,LogP值为5.3,可与血浆蛋白高度结合。犬口服给药后进食,生物利用度可达80%,而猫为100%,因此CredelioTM在进食前后服用较好。犬服用洛替拉纳后体内半衰期为30.7d(猫为33.6d),2h内(猫为4h)血药浓度达到最大值[143]。但静脉注射洛替拉纳,半衰期为24.6d,代谢清除率低。这种长半衰期和高生物利用度的特性可以使洛替拉纳维持较高的血药浓度,快速杀灭寄生虫,同时能预防寄主再次被感染[7, 138, 152]。
Toutain等[150~153]研究发现,洛替拉纳主要通过粪便排出,在血液和组织中,几乎不存在任何代谢物。与氟雷拉纳、阿福拉纳和沙罗拉纳相比,洛替拉纳的半衰期最长(约为4周,其他化合物约为2周),这种差异可能是由于洛替拉纳在体内分布的体积最大(约为6L/kg,其他化合物为3L/kg)。Kuntz等[142, 153, 154]评估了猫在口服洛替拉纳后的临床症状,仅观察到如腹胀、软/水样粪便等常见症状。研究表明,在剂量范围内(20~43 mg/kg)口服给药时,洛替拉纳对动物体均是安全的。此外,氟雷拉纳对犬的神经系统有一定程度的损伤,但洛替拉纳未观察到因治疗引起的相关神经性临床症状。
氟噁唑酰胺(Fluxametamide)(图式 5)是由日本日产化学工业株式会社研发的一种新型异噁唑啉类广谱杀虫剂,化学名称为4-[5-(3, 5-二氯苯基)-4, 5-二氢-5-(三氟甲基)-3-异噁唑基-N-[(甲氧基氨基)-亚甲基]-2-甲基苯甲酰胺,其S构型为主要的活性成分[155~158]。氟噁唑酰胺对多种昆虫如鳞翅目、缨翅目和双翅目显示出较高的杀虫活性[155]。氟噁唑酰胺是新型杀虫剂Gracia(开发代码:NC-515)的主要活性成分,其于2018年后在韩国、日本上市,作为广谱叶面杀虫剂使用,特别对蔬菜、棉花、大豆和茶叶作物中的鳞翅目和白粉虱等害虫[156, 160, 165]防治效果显著。
Asahi等[159, 160]研究发现,氟噁唑酰胺在亚纳摩尔浓度下可以抑制放射性配体[3H]EBOB与家蝇头部细胞膜的特异性结合,氟噁唑酰胺的抑制率约为80%,而氟虫腈完全抑制其结合。电生理测试显示,氟噁唑酰胺对家蝇GABA和谷氨酸门控氯离子通道均有抑制活性,IC50值分别为1.95和225 nmol/L[156~159, 161],这表明氟噁唑酰胺对家蝇GABA和谷氨酸门控氯离子通道具有选择性[156, 159, 162, 163];此外,研究表明氟噁唑酰胺对大鼠α1β2γ2 GABA受体和人α1甘氨酸受体几乎不产生抑制作用[159]。因此,氟噁唑酰胺对人类和其他哺乳动物可能具有较高的选择性和安全性。
Li等[166]通过同源建模和分子对接的方法,分析了氟噁唑酰胺对映体对小菜蛾GABA受体的选择性机理。结果显示,S-(+)-氟噁唑酰胺与小菜蛾GABA受体结合得更好,其结合自由能低于R-(-)-氟噁唑酰胺;S-(+)-氟噁唑酰胺可与Leu305形成卤键,与Tyr253和Lys307形成氢键,同时与Lys307还形成阳离子-π相互作用。Li等认为氟噁唑酰胺在小菜蛾GABA受体中的结合位点位于TMD的离子通道内,类似于氟雷拉纳,而伊维菌素结合于通道外,因此,氟噁唑酰胺的结合方式与伊维菌素等大环内酯类有较大区别。
氟噁唑酰胺主要用于农业和公共卫生害虫的防治,对家蝇有较高的杀虫活性,LD50和LD90值分别为12.9和38.7 ng/头[91, 156, 159]。氟噁唑酰胺对二斑叶螨的杀虫活性比氟虫腈高约50倍(LC50值分别为5.02和264 mg/L)[159, 164],而氟虫腈对褐飞虱(Nilaparvata lugens)的杀虫活性比氟噁唑酰胺高约17倍(LC50值分别为0.277和4.67 mg/L);此外,研究表明昆虫对氟虫腈和氟噁唑酰胺没有交叉抗性[160, 165]。Li等[166]优化了氟噁唑酰胺对映体的分离和制备方法,发现并证实了S-(+)-氟噁唑酰胺是主要的活性成分,且该构型对小菜蛾、甜菜夜蛾(Spodoptera exigue)、棉蚜(Aphis gossypii)、二斑叶螨四种靶标生物的活性是消旋体的2.5~3.7倍,R-(-)-氟噁唑酰胺几乎无活性。其消旋体对蜜蜂(Apis mellifera ligustica)显示的毒性最高,为活性异构体的4.3倍,因此在农业生产上,尽量避免使用氟噁唑酰胺的消旋体制剂。Ozawa等[167, 168]测试了氟噁唑酰胺(含量为8%的乳油剂型)对茶树常见害虫红点唇瓢虫(Chilocoruskuwanae silvestri)和方突毛瓢虫(Pseudoscymnus hareja)的杀虫活性,死亡率达到100%需要的时间分别为6d和2d。Tokumaru等[164]测试氟噁唑酰胺对蓑蛾类(Psychidae)害虫幼虫的杀虫活性,结果表明氟噁唑酰胺对茶蓑蛾(Eumeta minuscula butler)无杀虫活性,对褐蓑蛾(Mahasena aurea butler)杀虫活性较高,同时能够延缓害虫抗性的产生。以上研究表明,氟噁唑酰胺对许多主要虫害具有功效,杀虫谱较广[168],在农业和公共卫生害虫防治方面有潜在的应用价值。
日本食品安全委员会(Food Safety Commission of Japan)对氟噁唑酰胺进行了农药安全性评价[170]。大鼠在服用氟噁唑酰胺后,在8h后达到血浆最大血药浓度,半衰期为126h,生物吸收率为17.6%~27.4%。氟噁唑酰胺在大鼠体内主要分布在脂肪中,肝脏和肾脏中含量较低,其主要代谢途径是:(1)氯苯环的氢氧化;(2)异噁唑啉环的开裂;(3)侧链的水解。大鼠粪便中存在少部分未代谢的氟噁唑酰胺和代谢产物4-(3-(3, 5-二氯苯基)-4, 4, 4-三氟-3-羟基丁酰基)-N-甲酰基-2-甲基苯甲酰胺。毒性试验结果显示,氟噁唑酰胺对大鼠造成的不良症状主要是肺泡巨噬细胞聚集、小肠上皮细胞空泡化及肝细胞空泡化等,未发现神经毒性、繁殖影响、致畸形及遗传毒性。
Li等[166]测得S-(+)-氟噁唑酰胺在卷心菜、白菜和土壤中的半衰期分为0.84、4.23、2.26 d,与R-(-)-氟噁唑酰胺(为别为0.85、4.21、2.10 d)无明显区别,氟噁唑酰胺对映体在两种蔬菜和土壤中降解均较快。农作物残留试验结果表明,氟噁唑酰胺在叶状莴苣(茎叶)残留较多,最大值为5.23mg/kg,而在草莓和茄子等作物茎叶部的残留值为0.087~4.68 mg/kg,低于检测界限,且根部含量极低。土壤残留试验中,检测出氟噁唑酰胺易从水层转移到土壤层,其在好氧土壤中半衰期为52d,而厌氧土壤中的半衰期为498d[170]。
先正达公司的Crossthwaite等报道了一种新型异噁唑啉类杀虫剂Isocycloseram(图式 5),研发代号为SYN547407,于2018年6月经国际标准化组织农药通用名技术委员会批准获英文通用名。其化学名为4-[5-(3, 5-二氯-4-氟苯基)-4, 5-一氢-5-(三氟甲基)-3-异噁唑基]-N-(2-乙基-3-氧代-4-异噁唑烷基)-2-甲基苯甲酰胺,为4种活性异构体的混合物,其中(5S, 4R)-异构体活性最高。Isocycloseram是一种广谱杀虫剂和杀螨剂,对鳞翅目、半翅目、鞘翅目、翅目和二翅目等害虫均有杀虫活性。Isocycloseram与吡啶喹唑啉混配使用时,对番茄烟粉虱的杀虫活性增加,可减少农药使用成本,延缓害虫抗药性的产生[169]。其与氟雷拉纳可能具有相同的结合位点,有望用于农业害虫防治。
自氟雷拉纳作为体外杀寄生虫剂上市以来,其优良的杀虫特性和安全性促进了其他同类型异噁唑啉类杀虫剂的研究。目前为止,只有氟噁唑酰胺被用在农业害虫的预防,而氟雷拉纳、阿福拉纳、沙罗拉纳和洛替拉纳仅用于动物体外寄生虫的治疗和预防,使用范围有一定的局限性。研究证明氟雷拉纳等异噁唑啉类杀虫剂对农业害虫的杀虫活性较好,能为其他同类农业杀虫剂的开发提供很好的借鉴价值。异噁唑啉类化合物作为GABA受体新型NCA,与传统的杀虫剂相比其作用位点和作用方式尚不明确,需进一步探索;阿福拉纳等其他几种化合物均是基于氟雷拉纳的结构基础设计研发,结构有一定的相似性,进一步明确该类化合物与GABA受体的作用机制,进行合理的药物设计,可以为新型农业杀虫剂的研发提供新的思路和指导。
Weber T, Selzer P M. Chem. Med. Chem., 2016, 11(3): 270~276.
陈佳丽, 翟纳, 刘根炎, 等.武汉工程大学学报, 2019, 41(1): 13~18.
郑小娇, 李华光, 刘根炎, 等.农药学学报, 2017, 19(6): 665~671.
巨修炼.世界农药, 2007, 29(1): 28~33.
Casida J E, Durkin K A. Pestic. Biochem. Physiol., 2015, 121(2): 22~30.
Halos L, Beugnet F, Cardoso L, et al. Trends Parasitol., 2014, 30(5): 228~233.
Wright I. Parasit. Vector., 2018, 11(1): 407~410.
Casida J E. Chem. Res. Toxicol., 2015, 28(4): 560~566.
Weston D P, Lydy M J. Environ. Sci. Technol., 2014, 48(2): 1290~1297.
Gibbons D, Morrissey C, Mineau P. Environ. Sci. Pollut. Res., 2015, 22(1): 103~118.
Garcia-Reynaga P, Zhao C Q, Sarpong R. Chem. Res. Toxicol., 2013, 26(4): 514~516.
Rahman M M, Liu G Y, Furuta K, et al. J. Pestic. Sci., 2014, 39(3): 133~143.
Johnston G A. Neurochem. Res., 2014, 39(10): 1942~1947.
Liu G Y, Furuta K, Nakajima H, et al. Bioorgan. Med. Chem., 2014, 22(17): 4637~4645.
Liu G Y, Ozoe F, Furuta K, et al. J. Agr. Food Chem., 2015, 63(28): 6304~6312.
李明辉, 松布尔, 穆赫塔尔·伊米尔艾山.有机化学, 2018, 11(2): 378~400.
张平竹, 王晓芬, 郑雪阳, 等.有机化学, 2018, 38(12): 150~157.
Lopes E F, Penteado F, Thurow S, et al. J. Org. Chem., 2019, 84(19): 12452~12460.
Kamal A, Bharathi E V, Reddy J S, et al. Eur. J. Med. Chem., 2012, 46(2): 691~703.
Fery-Forgues S, Delavaux-Nicot B. J. Photoch. Photobio. A, 2000, 132(3): 137~159.
Mita T, Kikuchi T, Mizukoshi T, et al. Am. Historical Rev., 2018, 18(2): 525~526.
张一宾.世界农药, 2015, 37(1): 4~8.
Yang R, Lv M, Xu H. J. Agr. Food Chem., 2018, 66(43): 11254~11264.
何秀玲.世界农药, 2015, 37(1): 58~59.
Wang D W, Zhang R B, Yu S Y, et al. J. Agr. Food Chem., 2019, 67(45): 12382~12392.
Zhang J F, Xu J Y, Wang B L, et al J. Agr. Food Chem., 2012, 60(31): 7565~7572.
Liu G Y, Li H G, Shi J, et al. Bioorg. Med. Chem., 2019, 27(2): 416~424.
Ozoe Y, Asahi M, Ozoe F, et al. Biochem. Biophys. Res. Commun., 2010, 391(1): 744~749.
Asahi M, Kobayashi M, Matsui H, et al. Pest. Manag. Sci., 2015, 71(1): 91~95.
Gassel M, Wolf C, Noack S, et al. Insect. Biochem. Mol. Biol., 2014, 45(2): 111~124.
Zhao C Q, Casida J E. J. Agr. Food Chem., 2014, 62(5): 1019~1024.
Djuric M, Matic N, Davitkov D, et al. Parasit. Vector., 2019, 12(1): 270~276.
Walther F M, Allan M J, Roepke R K, et al. Part. Fibre. Toxicol., 2014, 7(1): 1~4.
Williams H, Young D R, Qureshi T, et al. Parasit. Vector., 2014, 7(1): 275~280.
Meadows C, Guerino F, Sun F. Parasit. Vector., 2014, 7(1): 375~379.
Price K L, Lummis S C. Insect Biochem. Mol. Biol., 2014, 55(2): 19~25.
K Timm, R Mueller, C S Nett. Vet. Dermatol., 2016. 27: 5~10.
Walther F M, Paul A J, Allan M J, et al. Parasit. Vector., 2014, 7(1): 89~91.
Dryden M W, Smith V. Parasit. Vector., 2014, 8(1): 364~369.
Fisara P, Webster M. Parasit. Vector., 2015, 8(1): 257~260.
Sheinberg G, Romero C, Heredia R, et al. Vet. Dermatol., 2017, 28(4): 393~e91.
Fourie J, Meyer L, Thomas E. Parasit. Vector., 2019, 12(1): 59~65.
赵春青, 韩召军, 唐涛.农药学学报, 2015, 17(3): 251~256.
Taenzler J, de Vos C, Heckeroth A R, et al. Parasit. Vector., 2017, 10(1): 30~36.
Beugnet F, Liebenberg J, Halos L. Vet. Parasitol., 2015, 209(1~2): 142~145.
Dayan F E, Haesaert G, Crossthwaite A, et al. Outlooks Pest Manag., 2019, 30(4): 157~163.
Kita T, Irie T, Nomura K, et al. Neurotoxicology, 2017, 60(1): 245~253.
Liu D, Jia Z Q, Peng Y C, et al. Pestic. Biochem. Physiol., 2018, 152: 8~16.
Mullens B A, Murillo A C, Zoller H, et al. Parasit. Vector., 2017, 10(1): 358~364.
Rowe M L, Whiteley P L, Carver S. Parasit. Vector., 2019, 12(1): 99~112.
Sheng C W, Jia Z Q, Ozoe Y, et al. Insect Biochem. Mol. Biol., 2018, 94(2): 18~27.
Emmanuel T, Hartmut Z, Gabriele L, et al. Parasit. Vector., 2018, 11(1): 361~367.
Brauneis M D, Zoller H, Williams H, et al. Parasit. Vector., 2017, 10(1): 594~601.
Kilp S, Ramirez D, Allan M J, et al. Parasit. Vector., 2016, 9(1): 296~300.
Zewe C M, Altet L, Lam A T, et al. Vet. Dermatol., 2017, 28(5): 46~e107.
Jiang S, Maia T, Ulrich B, et al. Int J. Environ. Res. Public Health, 2017, 14(2): 154~170.
蒋洁, 张莹, 唐涛, 等.中华卫生杀虫药械, 2019, 25(5): 492~498.
孙洪扬, 杨旭, 张静, 等.精细化工, 2020: 1~7.
凌汉, 李创, 唐涛, 等.环境昆虫学报, 2019, 41(4): 875~881.
Leviticus K, Cui L, Ling H, et al. Pest Manag. Sci., 2019, 76(3): 888~893.
Brauneis M D, Zoller H, Williams H, et al. Parasit. Vector., 2017, 10(1): 594~599.
Thomas E, Chiquet M, Sander B, et al. Parasit. Vector., 2017, 10(1): 457~461.
Sojka P A. J. Exotic Pet. Med., 2018, 27(2): 118~122.
Taenzler J, De Vos C, Roepke R K, et al. Parasit. Vector., 2017, 10(1): 30~37.
Taenzler J, Liebenberg J, Roepke R K, et al. Parasit. Vector., 2015, 8(1): 305~315.
Ozoe Y. Adv. Insect Physiol., 2013, 44(1): 211~286.
Ratra G S, Casida J E. Toxicol. Lett., 2001, 122(3): 215~222.
Nakata Y, Fuse T, Yamato K, et al Mol. Pharmacol., 2017, 92(5): 546~555.
Kilp S, Ramirez D, Allan M J, et al. Parasit. Vector., 2016, 9(1): 296~302.
Pfister K, Armstrong R. Parasit. Vector., 2016, 9(1): 436~451.
Asahi M, Kobayashi M, Matsui H, et al. Pest Manag. Sci., 2015, 71(1): 91~95.
Zhao C Q, Hwang S H, Buchholz B A, et al. PNAS, 2014, 111(23): 8607~8612.
Duangkaew L, Hoffman H. Vet. Dermatol., 2018, 29(3): 262~262.
Romero C, eredia H R, Pineda J, et al. Vet. Dermatol., 2016, 27(5): 353~e88.
Sheinberg G, Romero C, Heredia R, et al. Vet. Dermatol., 2017, 28(4): 393 ~e391.
Huyghe B, Traon L G, Flochlay-Sigognault A. Parasit. Vector., 2017, 10(1): 540~545.
Sheng C W, Huang Q T, Liu G Y, et al. Pest Manag. Sci., 2019, 75(11): 2901~2909.
Huang Q T, Sheng C W, Jiang J, et al. Insect Mol. Biol., 2018, 28(3): 313~320.
Siviter H, Brown M, Leadbeater E. Nature, 2018, 561(7721): 109~112.
Sheng C W, Jia Z Q, Liu D, et al. J. Asia-Pac. Entomol., 2017, 20(4): 1213~1218.
Jia Z Q, Liu D, Sheng C W, et al. Env. Pollut., 2018, 232(1): 183~190.
Lavan R P, King A I, Sutton D J, et al. Vaccine, 2017, 35(13): 1668~1674.
Lebon W, Beccati M, Bourdeau P, et al. Parasit. Vector., 2018, 11(1): 506~515.
Paterson S. Top. Companion Anim. Med., 2019, 24: 452~460.
Beugnet F, Halos L, Larsen D, et al. Parasites, 2016, 23(1): 14~21.
Maugeri C, Alisi M A, Apicella C, et al. Bioorg. Med. Chem., 2008, 16(6): 3091~3107.
Mueller R S, Bensignor E, Ferrer L, et al. Vet. Dermatol., 2012, 23(2): 86~90.
Hampel V, Knaus M, Schafer J, et al. Parasite, 2018, 25(2): 63~71.
Kunkle B N, Drag M D, Chester T S, et al. Vet. Parasitol., 2014, 201(3~4): 204~206.
Baker C F, McCall J W, McCall S D, et al. Comp. Immunol. Microbiol. Infect. Dis., 2016, 49(1): 65~69.
Shoop W L, Hartline E J, Gould B R, et al. Vet. Parasitol., 2014, 201(3~4): 179~189.
Yonetake W, Fujii T, Naito M, et al. Vet. Parasitol., 2019, 270(S1): S38~S44.
Beugnet F, Halos L, Diane L, et al. Parasit. Vector., 2014, 21(42): 1~6.
Carithers D E, Crawford J, de Vos C, et al. Parasit. Vector., 2016, 9(1): 635~639.
Hunter J S, Dumont P, Chester T S, et al. Vet. Parasitol., 2014, 201(3~4): 207~211
Miglianico M, Eldering M, Slater H, et al. PNAS, 2018, 115(29): E6920~E6926.
Liebenberg J, Fourie J, Lebon W, et al. Parasites, 2017, 24(2): 39~45.
Packianathan R, Hodge A, Bruellke N, et al. Parasit. Vector., 2017, 10(1): 98~95.
Yarto J E, Osorio M R, Rangel D J, et al. Vet. Dermatol., 2019, 30(2): 167~e50.
Lahm G P, Cordova D, Barry J D, et al. Bioorg. Med. Chem. Lett., 2013, 23(10): 3001~3006.
Hinkle N C, Jirjis F, Szewcyzyk E, et al. Parasit. Vector., 2018, 11(5): 99~105.
Casida J E. J. Agr. Food Chem., 2018, 66(13): 3277~3290.
Letendre L, Harriman J, Drag M, et al. Vet. Pharmacol. Ther., 2017, 40(1): 35~43.
Letendre L, Huang R, Kvaternick V, et al. Vet. Parasitol., 2014, 201(3~4): 190~197.
Beugnet F, Halos L, Larsen D, et al. Parasite, 2016, 23(14): 1~8.
Beugnet F, Crafford D, Fourie J, et al. Vet. Parasitol., 2016, 226(2): 150~161.
Kunkle B, Daly S, Dumont P, et al. Vet. Parasitol., 2014, 201(3~4): 226~228.
Dryden M W, Canfield M S, Kalosy K, et al. Parasit. Vector., 2016, 9(1): 365~375.
Ohmes C M, Hostetler J, Davis W L, et al. Parasitol. Res., 2017, 114(S1): 81~94.
Baker C F, Mccall J W, Mccall S D, et al. Comp. Immunol. Microb., 2016, 49(2~3): 65~69.
Halos L, Wilfreid L, Karine C M. Parasit. Vector., 2014, 7(3): 425~430.
Fankhauser R, Hamel D, Dorr P, et al. Vet. Parasitol., 2017, 225(1~2): 117~122.
Padivitage N, Kumar S, Rustum A. J. AOAC Int, 2017, 100(1): 65~73.
E B Mitchell, J W McCall, C S Theodore, et al. Vet. Parasitol., 2014, 20(3~4): 223~225.
Dumont P, Blair J, Fourie J, et al. Vet. Parasitol., 2014, 201(3~4): 216~219.
Otranto D, Little S. Vet. Parasitol., 2017, 238(l): S1~S2.
Cole L M, Nicholson R A, Casida J E. Pestic. Biochem. Physiol., 1993, 46(5): 47~50.
Becskei C, Cuppens O, Mahabir S P. Vet. Dermatol., 2018, 29(3): 203~207.
Becskei C, Bock F D, Illambas J, et al. Vet. Parasitol., 2016, 222(1): 49~55.
Curtis M P, Vaillancourt V, Goodwin R M, et al. Bioorg. Med. Chem. Lett., 2016, 26(1): 56~61.
Six R H, Becskei C, Mazaleski M M, et al. Vet. Parasitol., 2016, 222(1): 62~66.
McTier T L, Chubb N, Curtis M P, et al. Vet. Parasitol., 2016, 222(3): 3~11.
McTier T L, Six R H, Fourie J, et al. Vet. Parasitol., 2016, 222: 12~17.
Geurden T, Becskei C, Grace S, et al. Vet. Parasitol., 2016, 222(2): 33~36.
Machado M A, Campos D R, Lopes N L, et al. Rev. Bras. Parasitol. Vet., 2019, 28(4): 760~763.
Packianathan R, Colgan S, Hodge A, et al. Parasit. Vector., 2017, 10(1): 387~395.
Carithers D E, Sheila G, Jordan C. Int. J. Appl. Res. Vet. Med., 2018, 16(1): 7~11.
Six R H, Young D R, Myers M R, et al. Parasit. Vector., 2016, 9(1): 79~85.
Six R H, Everett W R, Young D R, et al. Vet. Parasitol., 2016, 222(2): 28~32.
Six R H, Liebenberg J, Honsberger N A, et al. Parasit. Vector., 2016, 9(1): 90~96.
Otranto D, Little S. Vet. Parasitol., 2017, 238: S1~S2.
Geurden T, Becskei C, Vatta A F, et al. Vet. Parasitol., 2017, 238(S1): S3~S7.
Packianathan R, Hodge A, Bruellke N, et al. Parasit. Vector., 2017, 10(1): 98~105.
Armstrong R. Parasit. Vector., 2017, 10(1): 159~165.
Scott F, Franz L, Campos D R, et al. Parasit. Vector., 2017, 10(1): 390~395.
Yonetake W, Fujii T, Naito M, et al. Vet. Parasitol., 2019, 270(S1): S38~S44.
Cherni J A, Mahabir S P, Six R H. Vet. Parasitol., 2016, 222(1): 43~48.
Little S E. Parasit. Vector., 2017, 10(1): 539~542.
Cavalleri D, Murphy M, Gorbea R L, et al. Parasit. Vector., 2017, 10(1): 527~534.
Cavalleri D, Murphy M, Nanchen S, et al. Parasit. Vector., 2018, 11(1): 408~417.
Karadzovska D, Chappell K, Coble S, et al. Parasit. Vector., 2017, 10(1): 528~527.
Kuntz E A, Srinivas K. Parasit. Vector., 2017, 10(1): 538~544.
Lasmar P V, Murphy M, Nanchen S, et al. Parasit. Vector., 2018, 11(1): 537~543.
Cavalleri D, Murphy M, Seewald W, et al. Parasit. Vector., 2018, 11(2): 410~420.
Young L, Karadzovska D, Wiseman S, et al. Parasit. Vector., 2020, 13(1): 25~31.
Toutain C E, Wolfgang S, Martin J, et al. Parasit. Vector., 2018, 11(3): 412~418.
Otaki H, Sonobe J, Murphy M. Parasit. Vector., 2018, 11(1): 448~453.
Kuntz E A. Parasit. Vector., 2018, 11(1): 409~415.
Mermans C, Dermauw W, Geibel S, et al. Pest Manag. Sci., 2017, 73(12): 2413~2418.
Toutain C E, Wolfgang S, Jung M. Parasit. Vector., 2017, 10(1): 522~530.
Rufener L, Danelli V, Bertrand D, et al. Parasit. Vector., 2017, 10(1): 530~545.
Snyder D E, Wiseman S, Liebenberg J E. Parasit. Vector., 2017, 10(3): 532~539.
Crotti S, Iorio N D, Mazzanti A, et al. J Org. Chem., 2018, 83(20): 12440~12448.
Gaens D, Rummel C, Schmidt M, et al. BMC Vet. Res., 2019, 15(1): 283~288.
Bu C Y, Feng X J, Wang X Q, et al. J. Econ. Entomol., 2015, 108(2): 769~776.
Ji Y K, Yoon J C, Jong S K, et al. Korean J. Environ. Agr., 2019, 38(1): 1~9.
Nakao T, Naoi A, Hama M, et al. J. Econ. Entomol., 2012, 105: 1781~1796.
Wang D, Qiao X, Tao J, et al. ACS Appl. Mater. Interf., 2018, 10(31): 26451~26455.
Asahi M, Kobayashi M, Kagami T, et al. Pestic. Biochem. Physiol., 2018, 151: 67~72.
Bukinham S D, Ihara M, Sattelle D B, et al. Curr. Med. Chem., 2017, 24(2): 115~119.
Guo Y, Zhang Q, Liu Z, et al. Ind. Crop. Prod., 2019, 40(2): 134~141.
Lamberth C. J. Heterocyclic Chem., 2018, 55(9): 2035~2045.
Wang J D, Chen L F, Lin D J, et al. Pestic. Biochem. Physiol., 2019, 156(2): 56~62.
Tokumaru S, Yamashita K. Tea Res. J., 2016, 121: 17~21.
Sayyed A H, Wright D J. J. Econ. Entomol., 2004, 97(6): 2043~2050.
Li R, Pan X, Wang Q, et al. Environ. Sci. Technol., 2019, 53(23): 13657~13665.
Saelinger D, Dickhaut J, Koerber K, et al. US: 20170238554, 2017.
Ozawa A, Uchiyama T. Ann. Rept. Kansai PI. Prot., 2019, 61: 151~153.
王礼文. CN: 201910388199, 2019.
Food Safety Commission of Japan. Food Safety, 2020, 8(1): 10~11.

图式 5 沙罗拉纳、洛替拉纳、氟噁唑酰胺和Isocycloseram的化学结构
Scheme 5 Chemical structures of afoxolaner, lotilaner, fluxametamide, and isocycloseram
表 1 沙罗拉纳对蜱虫的杀虫谱
Table 1. Insecticidal spectrums of sarolaner against ticks
| 蜱虫种类 | 拉丁学名 | 文献 |
| 肩突硬蜱 | Ixodes scapularis | [132] |
| 篦子硬蜱 | Ixodes ricinus | [119, 120, 124] |
| 六角硬蜱 | Ixodes hexagonus | [119, 133] |
| 全沟硬蜱 | Ixodes pesuleatus | [132] |
| 全环硬蜱 | Ixodes holocyclus | [119] |
| 网纹革蜱 | Dermacentor reticulatus | [119, 126, 137] |
| 斑点钝眼蜱 | Amblyomma maculatum | [131] |
| 美洲钝眼蜱 | Amblyomma americanum | [134] |
| 变异革蜱 | Dermacentor variabilis | [124] |
| 血红扇头蜱 | Rhipicephalus sanguineus | [119, 124] |
 下载: 导出CSV
下载: 导出CSV
表 2 不同GABA门控氯离子通道对不同拮抗剂的效应[152]
Table 2. Effects of different GABA-gated chloride channels against different antagonists[152]
| GABA受体类型 | 拮抗剂 | IC50/(nmol/L) |
| DmR2-GABAR | Dieldrin | > 10, 000 |
| DmR2-GABAR | Fipronil | 230.32 |
| DmR2-GABAR | Lotilaner | 38.25 |
| DmS-GABAR | AHC-2229544 | 959.47 |
| DmS-GABAR | Dieldrin | 4170 |
| DmS-GABAR | Fipronil | 27.58 |
| DmS-GABAR | Lotilaner | 23.84 |
| Ls-GABAR | Fipronil | 164.85 |
| Ls-GABAR | Lotilaner | 52.40 |
| Rm-GABAR | Fipronil | 25.56 |
| Rm-GABAR | Lotilaner | 36.79 |
| α1β2γ2GABAAR | Lotilaner | > 10, 000 |
下载: 导出CSV

 扫一扫看文章
扫一扫看文章

扫一扫关注我们







 下载:
下载: