CAS Key Laboratory of Microbial Physiological and Metabolic Engineering, State Key Laboratory of Microbial Resources, Institute of Microbiology, Chinese Academy of Sciences, Beijing 100101
b.
College of Life Sciences, University of Chinese Academy of Sciences, Beijing 100149
Received Date:
13 June 2018 Revised Date:
08 August 2018 Available Online:
01 September 2018
Fund Project:
Project supported by the National Natural Science Foundation of China (Nos. 31470818, 31722002) and Ministry of Science and Technology of the People's Republic of China (No. 2015CB910300)
Abstract:
With the rapid progress of synthetic biology and other related filed, there is a continuous growth of the applications of genetically modified organisms in many aspects, including industry, agriculture, health and environment. However, unintended release or uncontrolled propagation of these genetically modified organisms may cause significant side effects to the nature ecological environment. In order to eradicate the escaping problem and horizontal gene transfer between artificial and natural organisms, many researches have been focused on how to limit genetically modified organisms to a controlled environment. The research progress of biocontainment of genetically modified organisms mainly from three aspects of traditional biocontainment strategies, the orthogonalization of central dogma and the design of complex genetic networks is highlighted. It is believed that the advanced biocontainment technology would promote the further application of synthetic biology.
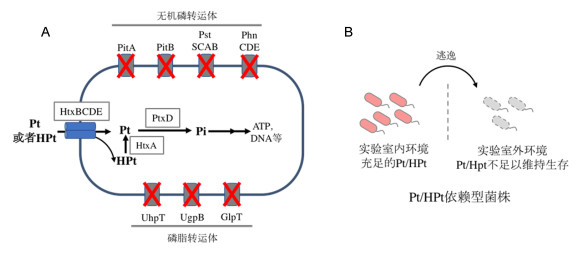
Figure 2.
Creation of engineered dependency on Pt/HPt
(A) Schematic of the engineered P metabolic pathway for biocontainment; (B) concept for the biocontainment strategy using engineered dependency on Pt/HPt
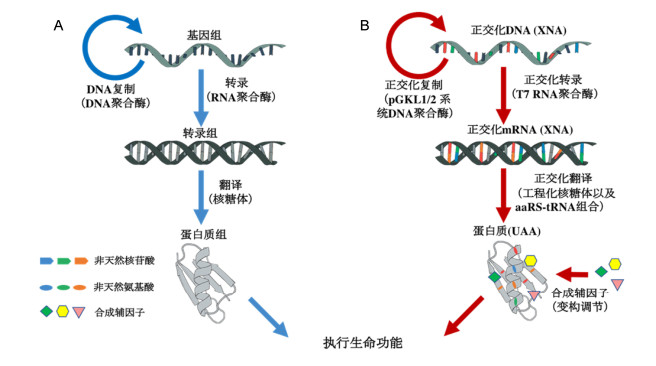
(A) The central dogma applies to most living organisms. Genetic information is stored by DNA replication, decoded into functional proteins by a transcription-translation process, and ultimately guides the life process; (B) the orthogonalization of central dogma includes two aspects, the introduction of non-natural chemicals into nucleic acids and proteins, and the orthogonalization of macromolecular machines
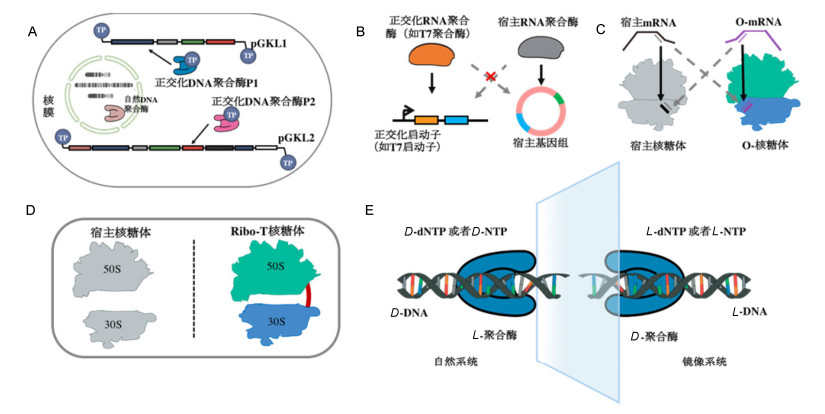
Wang等[58]从头合成了一种由D型氨基酸组成的镜像聚合酶-D型非洲猪瘟病毒聚合酶X (D-ASFV pol X).这一镜像聚合酶能以L型DNA为模板, 完成L型DNA或者L-RNA链的合成, 即D-ASFV pol X同时具有L型DNA与RNA的聚合酶活性, 使得镜像版本的DNA复制和转录这两个中心法则的关键步骤得以实现(图 5E).虽然D-ASFV pol X的持续合成能力较低且热稳定性差, 目前还不能在生命体内发挥作用, 但这为构建镜像生命体提供了可能, 也为实现人工生命体防逃逸提供了一种潜在的选择.
(a) Khalil, A. S.; Collins, J. J. Nat. Rev.Genet.2010, 11, 367. (b) Lee, J. W.; Na, D.; Park, J. M.; Lee, J.; Choi, S.; Lee, S. Y. Nat. Chem. Biol.2012, 8, 536. (c) Cameron, D. E.; Bashor, C. J.; Collins, J. J. Nat. Rev. Microbiol.2014, 12, 381.
(a) Alphey, L. Annu. Rev. Entomol.2014, 59, 205. (b) Gantz, V. M.; Jasinskiene, N.; Tatarenkova, O.; Fazekas, A.; Macias, V. M.; Bier, E.; James, A. A. Proc. Natl. Acad. Sci. U. S. A.2015, 112, E6736. (c) Harris, A. F.; Nimmo, D.; McKemey, A. R.; Kelly, N.; Scaife, S.; Donnelly, C. A.; Beech, C.; Petrie, W. D.; Alphey, L. Nat. Biotechnol.2011, 29, 1034. (d) Hammond, A.; Galizi, R.; Kyrou, K.; Simoni, A.; Siniscalchi, C.; Katsanos, D.; Gribble, M.; Baker, D.; Marois, E.; Russell, S.; Burt, A.; Windbichler, N.; Crisanti, A.; Nolan, T. Nat. Biotechnol.2016, 34, 78. (e) Lacroix, R.; McKemey, A. R.; Raduan, N.; Kwee Wee, L.; Hong Ming, W.; Guat Ney, T.; Rahidah, A. A. S.; Salman, S.; Subramaniam, S.; Nordin, O.; Hanum, A. T. N.; Angamuthu, C.; Marlina Mansor, S.; Lees, R. S.; Naish, N.; Scaife, S.; Gray, P.; Labbe, G.; Beech, C.; Nimmo, D.; Alphey, L.; Vasan, S. S.; Han Lim, L.; Wasi, A. N.; Murad, S. PLoS One2012, 7, e42771.
[8]
(a) Rovner, A. J.; Haimovich, A. D.; Katz, S. R.; Li, Z.; Grome, M. W.; Gassaway, B. M.; Amiram, M.; Patel, J. R.; Gallagher, R. R.; Rinehart, J.; Isaacs, F. J. Nature2015, 527. (b) Mandell, D. J.; Lajoie, M. J.; Mee, M. T.; Takeuchi, R.; Kuznetsov, G.; Norville, J. E.; Gregg, C. J.; Stoddard, B. L.; Church, G. M. Nature2015, 518, 55. (c) Lajoie, M. J.; Rovner, A. J.; Goodman, D. B.; Aerni, H. R.; Haimovich, A. D.; Kuznetsov, G.; Mercer, J. A.; Wang, H. H.; Carr, P. A.; Mosberg, J. A.; Rohland, N.; Schultz, P. G.; Jacobson, J. M.; Rinehart, J.; Church, G. M.; Isaacs, F. J. Science2013, 342, 357.
[9]
(a) Marlière, P.; Patrouix, J.; Döring, V.; Herdewijn, P.; Tricot, S.; Cruveiller, S.; Bouzon, M.; Mutzel, R. Angew. Chem., Int. Ed.2011, 50, 7109. (b) Pezo, V.; Liu, F. W.; Abramov, M.; Froeyen, M.; Herdewijn, P.; Marliere, P. Angew. Chem., Int. Ed.2013, 52, 8139. (c) Malyshev, D. A.; Dhami, K.; Lavergne, T.; Chen, T.; Dai, N.; Foster, J. M.; Corrêa, I. R.; Romesberg, F. E. Nature2014, 509, 385. (d) Malyshev, D. A.; Dhami, K.; Lavergne, T.; Chen, T.; Dai, N.; Foster, J. M.; Correa, I. R., Jr.; Romesberg, F. E. Nature2014, 509, 385.
[10]
(a) de Lorenzo, V. In Handbook of Hydrocarbon and Lipid Microbiologyed, Springer, Berlin, Heidelberg, 2010, p. 2435. (b) Solé, R. V.; Montañez, R.; Duran-Nebreda, S. Biology Direct2015, 10, 37.
[11]
Landrain, T.; Meyer, M.; Perez, A. M.; Sussan, R. Syst. Synth. Biol. 2013, 7, 115. doi: 10.1007/s11693-013-9116-4
[12]
Berg, P.; Baltimore, D.; Brenner, S.; Roblin, R. O.; Singer, M. F. Science 1975, 188, 991. doi: 10.1126/science.1056638
[13]
Dana, G. V.; Kuiken, T.; Rejeski, D.; Snow, A. A. Nature 2012, 483, 29. doi: 10.1038/483029a
(a) Wang, Z.; Xu, W.; Liu, L.; Zhu, T. F. Nat. Chem.2016, 8, 698; (b) Steidler, L.; Neirynck, S.; Huyghebaert, N.; Snoeck, V.; Ver- meire, A.; Goddeeris, B.; Cox, E.; Remon, J. P.; Remaut, E. Nat. Biotechnol.2003, 21, 785; (c) Cohen, S. S.; Barner, H. D. J. Biol. Chem.1957, 226, 631.
[16]
Curtiss, R.; Inoue, M.; Pereira, D.; Hsu, J. C.; Alexander, L.; Rock, L. In Molecular of Cloning of Recombinant DNA, Elsevier, Amsterdam, Netherlands, 1977, p. 99.
Szafranski, P.; Mello, C. M.; Sano, T.; Smith, C. L.; Kaplan, D. L.; Cantor, C. R. Proc. Natl. Acad. Sci. U. S. A. 1997, 94, 1059. doi: 10.1073/pnas.94.4.1059
Wang, L.; Brock, A.; Herberich, B.; Schultz, P. G. Science 2001, 292, 498. doi: 10.1126/science.1060077
[33]
Chin, J. W.; Cropp, T. A.; Anderson, J. C.; Mukherji, M.; Zhang, Z. W.; Schultz, P. G. Science 2003, 301, 964. doi: 10.1126/science.1084772
[34]
Wang, H. H.; Isaacs, F. J.; Carr, P. A.; Sun, Z. Z.; Xu, G.; Forest, C. R.; Church, G. M. Nature 2009, 460, 894. doi: 10.1038/nature08187
[35]
Isaacs, F. J.; Carr, P. A.; Wang, H. H.; Lajoie, M. J.; Sterling, B.; Kraal, L.; Tolonen, A. C.; Gianoulis, T. A.; Goodman, D. B.; Reppas, N. B.; Emig, C. J.; Bang, D.; Hwang, S. J.; Jewett, M. C.; Jacobson, J. M.; Church, G. M. Science 2011, 333, 348. doi: 10.1126/science.1205822
[36]
Ostrov, N.; Landon, M.; Guell, M.; Kuznetsov, G.; Teramoto, J.; Cervantes, N.; Zhou, M.; Singh, K.; Napolitano, M. G.; Moos-burner, M.; Shrock, E.; Pruitt, B. W.; Conway, N.; Goodman, D. B.; Gardner, C. L.; Tyree, G.; Gonzales, A.; Wanner, B. L.; Norville, J. E.; Lajoie, M. J.; Church, G. M. Science 2016, 353, 819. doi: 10.1126/science.aaf3639
[37]
(a) Wang, K.; Schmied, W. H.; Chin, J. W. Angew. Chem., Int. Ed.2012, 51, 2288; (b) Niu, W.; Schultz, P. G.; Guo, J. ACS Chem. Biol.2013, 8, 1640; (c) Anderson, J. C.; Wu, N.; Santoro, S. W.; Lakshman, V.; King, D. S.; Schultz, P. G. Proc. Natl. Acad. Sci. U. S. A.2004, 101, 7566.
[38]
Tack, D. S.; Ellefson, J. W.; Thyer, R.; Wang, B.; Gollihar, J.; Forster, M. T.; Ellington, A. D. Nat. Chem. Biol. 2016, 12, 138. doi: 10.1038/nchembio.2002
[39]
Wang, N. X.; Li, Y.; Niu, W.; Sun, M.; Cerny, R.; Li, Q. S.; Guo, J. T. Angew. Chem., Int. Ed. 2014, 53, 4867. doi: 10.1002/anie.201402092
[40]
Si, L. L.; Xu, H.; Zhou, X. Y.; Zhang, Z. W.; Tian, Z. Y.; Wang, Y.; Wu, Y. M.; Zhang, B.; Niu, Z. L.; Zhang, C. L.; Fu, G.; Xiao, S. L.; Xia, Q.; Zhang, L. H.; Zhou, D. M. Science 2016, 354, 1170. doi: 10.1126/science.aah5869
Yang, Z. Y.; Hutter, D.; Sheng, P. P.; Sismour, A. M.; Benner, S. A. Nucleic Acids Res. 2006, 34, 6095. doi: 10.1093/nar/gkl633
[43]
Yang, Z. Y.; Chen, F.; Alvarado, J. B.; Benner, S. A. J. Am. Chem. Soc. 2011, 133, 15105. doi: 10.1021/ja204910n
[44]
Kim, H. J.; Leal, N. A.; Hoshika, S.; Benner, S. A. J. Org. Chem. 2014, 79, 3194. doi: 10.1021/jo402665d
[45]
Leconte, A. M.; Hwang, G. T.; Matsuda, S.; Capek, P.; Hari, Y.; Romesberg, F. E. J. Am. Chem. Soc. 2008, 130, 2336. doi: 10.1021/ja078223d
[46]
Seo, Y. J.; Hwang, G. T.; Ordoukhanian, P.; Romesberg, F. E. J. Am. Chem. Soc. 2009, 131, 14596. doi: 10.1021/ja907027a
[47]
Malyshev, D. A.; Dhami, K.; Lavergne, T.; Chen, T. J.; Dai, N.; Foster, J. M.; Correa, I. R.; Romesberg, F. E. Nature 2014, 509, 385. doi: 10.1038/nature13314
[48]
Zhang, Y.; Ptacin, J. L.; Fischer, E. C.; Aerni, H. R.; Caffaro, C. E.; Jose, K. S.; Feldman, A. W.; Turner, C. R.; Romesberg, F. E. Nature 2017, 551, 644. doi: 10.1038/nature24659
[49]
Zhang, Y.; Lamb, B. M.; Feldman, A. W.; Zhou, A. X.; Lavergne, T.; Li, L.; Romesberg, F. E. Proc. Natl. Acad. Sci. U. S. A. 2017, 114, 1317. doi: 10.1073/pnas.1616443114
Chan, C. T.; Lee, J. W.; Cameron, D. E.; Bashor, C. J.; Collins, J. J. Nat. Chem. Biol. 2016, 12, 82. doi: 10.1038/nchembio.1979
[62]
Gallagher, R. R.; Patel, J. R.; Interiano, A. L.; Rovner, A. J.; Isaacs, F. J. Nucl. Acids Res. 2015, 43, 1945. doi: 10.1093/nar/gku1378
[63]
Cai, Y.; Agmon, N.; Choi, W. J.; Ubide, A.; Stracquadanio, G.; Caravelli, K.; Hao, H.; Bader, J. S.; Boeke, J. D. Proc. Natl. Acad. Sci. U. S. A. 2015, 112, 1803. doi: 10.1073/pnas.1424704112
[64]
Schmidt, M.; Pei, L. Toxicol. Sci. 2010, 120, S204.
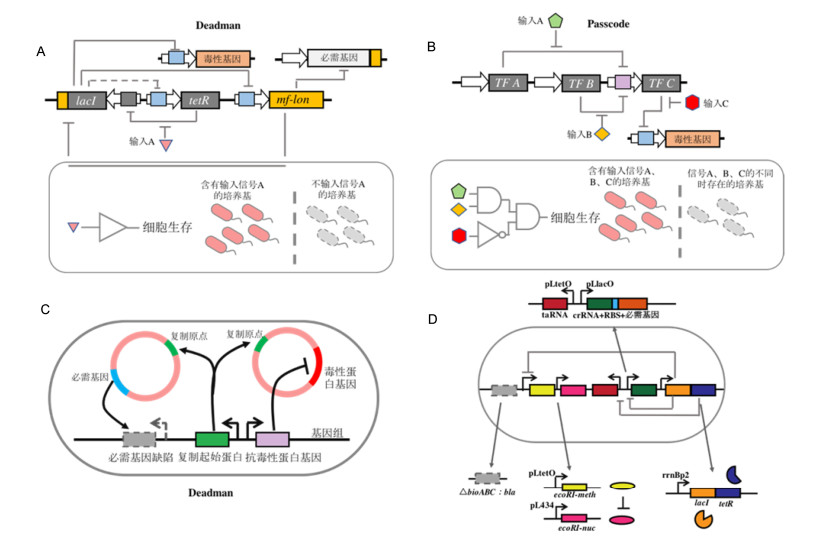
图 1
传统遗传改造生命体的防逃逸策略
Figure 1
Established biocontainment strategies of genetically modified organisms
(A) Auxotroph; (B) simple kill switches; (C) inducible gene switch to control essential gene; (D) gene flow barrier
Figure 2
Creation of engineered dependency on Pt/HPt
(A) Schematic of the engineered P metabolic pathway for biocontainment; (B) concept for the biocontainment strategy using engineered dependency on Pt/HPt
(A) The central dogma applies to most living organisms. Genetic information is stored by DNA replication, decoded into functional proteins by a transcription-translation process, and ultimately guides the life process; (B) the orthogonalization of central dogma includes two aspects, the introduction of non-natural chemicals into nucleic acids and proteins, and the orthogonalization of macromolecular machines


 下载:
下载:

 下载:
下载:







 下载:
下载:
