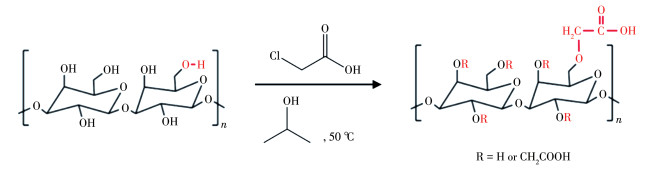
Scheme 1.
Carboxylation of Desmodium styracifolium polysaccharide (DSP0)

肾结石中存在多种矿化物成分,包括草酸钙(CaC2O4)(75%)、碳磷灰石(10.3%)、尿酸盐(10%)、鸟粪石(8%)和透磷石(1.3%)等[1-3];在CaC2O4结石中,一水合草酸钙(COM)和二水合草酸钙(COD)的质量分数分别约为53%和22%[4-5]。减少肾脏中的CaC2O4的结晶以及晶体-细胞间的黏附可以降低肾结石的形成风险[6]。
植物多糖具有抗氧化、抗肿瘤、抗病毒和免疫调节等活性[7-9],但天然多糖通常具有的高分子量和较低的活性基团含量使其生物活性受到限制。对多糖进行化学修饰可以提高多糖的生物活性。例如,Xie等[10]将青钱柳多糖(CP)分别进行硫酸化和羧基化增加其活性基团含量,得到的衍生物(S-CP)和(CM-CP)比CP更能减少细胞活性氧(ROS)的产生,也比CP更能抑制异常细胞凋亡,更有效地保护了RAW264.7巨噬细胞免受H2O2诱导的细胞损伤。羧基化后的茯苓多糖(CMP)比未修饰的茯苓多糖(MP)更能增加人肾近端曲小管上皮细胞(HK-2细胞)的NO、TNF-α和IL-6的分泌,降低ROS的产生,表现出更好的免疫调节作用[11]。Zhao等[12]制备了取代度为0.88的羧基化五味子多糖(CSPP),其比未修饰的多糖更能逆转多氯联苯126氧化应激引起的小鼠脾脏损伤,原因之一可能是羧甲基的插入减少了分子内和分子间的相互作用,从而使分子的溶解度和活性增加[13]。然而,也有一些研究表明,并非所有的化学修饰都能增强多糖的活性。例如,在40 mg·L-1时,羧基化后的石斛多糖对RAW264.7巨噬细胞的增殖作用由羧基化前的147.6%降至107.4%[14]。
广金钱草分布于广东、广西和海南,已广泛应用于治疗肾结石和肾损伤。广金钱草可以减少尿中草酸盐的排泄,促进尿液排出,减少肾脏中的CaC2O4沉积,并通过抗炎和抗氧化功能减轻晶体对肾脏的损伤[14-16]。广金钱草的主要药效成分为黄酮、多糖和三萜类化合物[17]。广金钱草多糖(DSP)具有抗氧化、消炎和镇痛作用[18-19]。例如,DSP对超氧阴离子和各类自由基具有清除能力[18];对离体膀胱平滑肌有一定的收缩作用,从而促进排尿。
我们通过对—COOH含量(质量分数)为1.17%的原始广金钱草多糖(DSP0)羧基化[20],获得了—COOH含量分别为7.45%、12.2%和17.7%的3种衍生多糖DSP1、DSP2和DSP3。DSP0不含支链,主要是由葡萄糖残基(占98.24%)相连的嵌段;在羧甲基修饰后,DSP0的部分羟基被—CH2COOH取代,没有新的支链形成。羧基化修饰能够明显提升多糖的体外抗氧化活性和修复受损伤细胞的能力。本工作进一步研究这4种具有不同—COOH含量的DSPs对CaC2O4晶体生长、聚集的调控作用以及对细胞表面CaC2O4晶体黏附的抑制作用,期望为开发新型防结石药物提供更多的理论依据。
生化试剂和材料包括HK-2细胞(中科院上海细胞库)、胎牛血清和培养液DMEM-F12(Gibco,美国Gibco公司)、青霉素和链霉素(北京普博生物技术有限公司)、6和96孔细胞培养板(NEST,中国)、细胞增殖毒性检测试剂盒(Cell Counting Kit-8,CCK-8)、ELISA试剂盒(上海碧云天生物技术有限公司)。其余试剂均为市售。
原始广金钱草多糖(DSP0)来自西安青芷生物技术有限公司,其—COOH含量为1.17%。DSP0羧基化参照文献[20]进行(Scheme 1),获得的衍生多糖DSP1、DSP2和DSP3中—COOH含量分别为7.45%、12.2%和17.7%。
主要仪器有D/max2400粉末X射线衍射仪(日本理学)、傅里叶变换红外光谱仪(美国Nicolet公司)、场发射扫描电子显微镜(ULTRA55,德国蔡司)、Zsizer Nano ZS型的激光纳米粒度仪(英国Mallern公司)、热重分析仪(TGA/DSC 3+,美国Mettler Toledo)、多功能酶标仪(SafireZ,瑞士Tecan)、流式细胞仪(FACS Aria,美国BD)。
实验步骤在文献[21]的基础上略加修改。在一组烧杯中各加入40 mL CaCl2溶液(22 mmol·L-1),然后加入一定量的多糖,补充蒸馏水至48 mL,磁力搅拌10 min后,加入Na2C2O4溶液(40 mL,22 mmol·L-1),使体系终体积为88 mL,此时溶液中 cCa2+=cC2O42-=10 mmol·L-1,DSPs最终质量浓度为0.1、0.2、0.4、0.8、1.0 g·L-1。搅拌反应10 min后,在37 ℃恒温静置2 h,离心,用无水乙醇清洗沉淀3遍,置于50 ℃烘箱中干燥。然后对形成的CaC2O4晶体进行FTIR、扫描电子显微镜(SEM)、粉末X射线衍射(PXRD)、ζ电位和热重分析(TGA)表征。
取上述合成的晶体在基片上,于粉末X射线衍射仪中分析,测试条件:Cu Kα射线,波长0.154 16 nm,石墨单色器,30 kV,25 mA,扫描范围5°~60°,扫描速度8 (°)·min-1,步宽0.02 (°)·s-1,进行定性和定量分析。CaC2O4中COM和COD的相对百分含量采用K值法计算[22],如COD的相对质量分数:
|
|
其中ICOD和ICOM分别为COD的(200)晶面(d=0.618 nm)主衍射峰和COM的(101)晶面(d=0.593 nm)主衍射峰的强度。取2 mg的已干燥的CaC2O4样品,与200 mg KBr充分混匀,用玛瑙研钵研磨至看不见金属光泽的粉末,压片,于4 000~400 cm-1波数范围内扫描。采用氮气流,升温速度为10 ℃·min-1,在25~900 ℃测定有没有多糖存在时生成的CaC2O4晶体的TG曲线,分析其热稳定性,计算晶体中的多糖含量。取CaC2O4样品1 mg分散在10 mL无水乙醇中,在低功率下超声3 min后,点样在10 mm×10 mm的玻片上,于50 ℃下烘干。样品喷金后用SEM观察晶体的尺寸与形貌。称取各多糖调控的晶体10 mg,超声10 min使其分散在30.0 mL纯水中,采用纳米粒度仪检测晶体的ζ电位。
HK-2细胞采用含有10%胎牛血清和1%青霉素、链霉素的DMEM-F12培养基在37 ℃、CO2体积分数5%和饱和湿度下的培养箱中培养。采用胰蛋白酶消化法对细胞进行消化传代,当细胞达到80%~90%汇合后,除去原培养液并用磷酸盐缓冲液(PBS)洗涤细胞2次,加入0.25%的胰蛋白酶-EDTA消化液,于37 ℃培养箱放置2~4 min,在光学显微镜下观察消化程度,当细胞彼此间连接疏松且即将悬浮于溶液时为消化适度,其后加入含10%胎牛血清的DMEM/F-12细胞培养液终止消化,充分吹打分散细胞,形成单细胞悬液。将HK-2细胞以密度1×105 mL-1、每孔100 μL接种于96孔板中,放置于37 ℃、CO2体积分数5%的培养箱中孵育24 h,使细胞汇合成单层,改用无血清培养液孵育12 h,弃培养液,用PBS洗涤细胞2次。实验模型分为3组:(1) 对照组,仅加入无血清培养基培养24 h;(2) nano-COM损伤组,加入含200 μg·mL-1 nano-COM的无血清培养基培养24 h;(3) 多糖保护组,将80 μg·mL-1的各种DSPs分别与nano-COM混合,然后与HK-2细胞共孵育24 h。
对于CD44检测,细胞培养和分组同上。达到培育时间后,吸除培养液,用PBS清洗2遍后加入多聚甲醛固定15 min,接着用羊血清封闭20 min,然后滴加CD44一抗在4 ℃孵育过夜。孵育时间到达后,用PBS洗涤3次,避光滴加iFluor 594二抗,在37 ℃孵育0.5 h。最后用DAPI标记细胞核,通过共聚焦显微镜观察CD44表达。
Annexin A2检测的细胞培养和分组同CD44检测。用羊血清封闭20 min后,滴加Annexin A2一抗,在4 ℃孵育过夜。孵育时间到达后,用PBS洗涤3次,避光滴加iFluor 594二抗,在37 ℃孵育0.5 h后,用DAPI标记细胞核,通过共聚焦显微镜观察Annexin A2表达。
Nano-COM的FITC荧光标记:在74 ℃使AMPTES(5 mL)与COM(0.05 g)在无水乙醇(50 mL)中反应3 h。随后加入FITC(0.025 g),反应保持6 h。过滤,用无水乙醇和去离子水洗涤数次,直至不含游离FITC。干燥,备用。
荧光显微镜定性观察晶体黏附:细胞培养和分组同上。转至4 ℃下培养6 h后,每孔板细胞用300 μL的DiI染色10 min,细胞核用DAPI染色,4%多聚甲醛固定10 min。采用激光共聚焦显微镜定性观察nano-COM晶体在细胞内外的定位。
流式细胞术定量检测晶体黏附:到达培养时间后,吸除培养基,用PBS洗2次,去掉未黏附和黏附不紧的COM,胰酶消化重悬细胞。采用流式细胞仪检测黏附晶体的细胞比例,有FITC信号的可以视为有黏附晶体的细胞。
实验获得的数据用均数±标准差(x±SD)表示。实验结果采用SPSS 13.0软件(SPSS Inc.美国Chicago,IL)统计分析,采用Tukey检验分析各实验组与对照组均数间的差异。P < 0.05表示有统计学意义,存在显著性差异;P < 0.01表示有极显著性差异;P > 0.05表示无显著性差异。
图 1为不同质量浓度的—COOH含量分别为1.17%、7.45%、12.2%和17.7%的4种多糖DSP0、DSP1、DSP2和DSP3调控生成的CaC2O4晶体的PXRD图。0.1~1.0 g·L-1的DSP0只诱导形成了COM晶体(图 1A),COM的衍射峰出现在2θ=14.88°、24.2°、30.16°和38.24°处,分别归属于COM的(101)、(020)、(202)和(130)晶面。
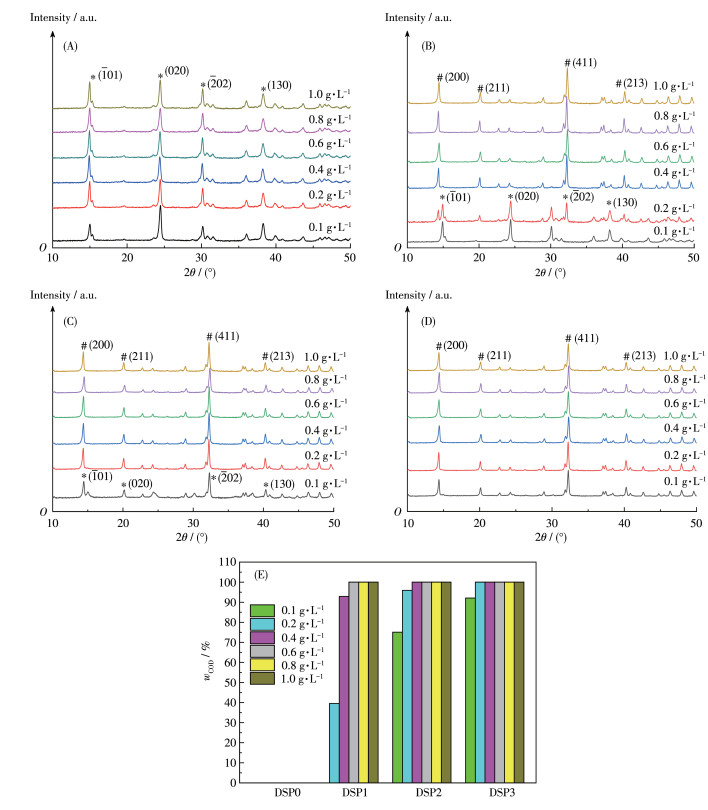
*means the characteristic diffraction peak of COM, and # means the characteristic diffraction peak of COD.
对于DSP1,在0.1 g·L-1时也只诱导COM形成(图 1B)。但0.2~0.4 g·L-1时诱导了COM和COD形成,COD的衍射峰出现在2θ=14.28°、20.12°、32.28°和40.28°处,分别归属于COD的(200)、(211)、(411)和(213)晶面。对PXRD图进行定量分析表明,ρDSP1=0.2、0.4 g·L-1时,生成的晶体中COD含量分别为39.6%、92.9%。当ρDSP1≥0.6 g·L-1时,全部形成COD晶体,没有COM形成。
在质量浓度为0.2 g·L-1时,DSP0、DSP1、DSP2和DSP3诱导生成的CaC2O4晶体中COD含量依次为0、39.6%、75.1%和92.1%。而DSP1、DSP2和DSP3诱导100%COD形成的最低质量浓度分别为0.6、0.4和0.2 g·L-1(图 1B~1E),说明随着多糖中的—COOH含量升高或多糖浓度增加,其诱导生成COD的能力均增强。
图 2为不同质量浓度的DSPs调控生成的CaC2O4晶体的FTIR谱图,其中COM在3 490~3 059 cm-1处归属于结晶水O—H键的拉伸振动峰分裂为5个小的吸收峰,而COD在该区域中只有1个宽吸收峰;COM晶体中的羰基不对称伸缩振动νas(COO-)出现在1 618 cm-1,对称伸缩振动νs(COO-)出现在1 316 cm-1;而COD晶体的νas(COO-)和νs(COO-)分别在1 647和1 324 cm-1。在指纹区,COM的吸收带在951、886和671 cm-1处,而COD在916和615 cm-1处。
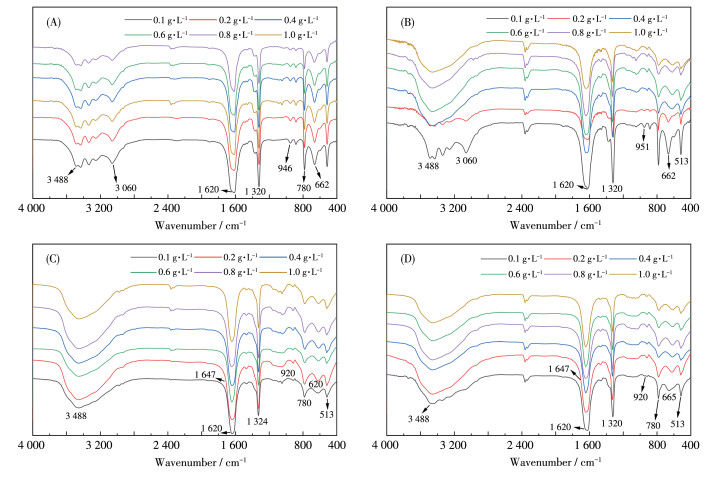
从图 2A可以看出,DSP0诱导形成的晶体均只出现COM的吸收峰。ρDSP1=0.1 g·L-1时形成的晶体的FTIR谱图与图 2A没有明显区别,即只形成COM晶体。但随着ρDSP1增加,在3 490~3 059 cm-1处5个小峰逐渐消失,νas(COO-)和νs(COO-)也不断蓝移,表明COD占比增加。相比之下,DSP2和DSP3诱导形成CaC2O4晶体的FTIR谱均与COD晶体接近,表明DSP2和DSP3主要诱导COD晶体形成。各晶体的νas(COO-)和νs(COO-)吸收峰值见表 1。
 下载:
导出CSV
下载:
导出CSV
| DSP | ρDSP/(g·L-1) | wCOD/% | COM | COD | |||
| νas(COO-)/cm-1 | νs(COO-)/cm-1 | νas(COO-)/cm-1 | νs(COO-)/cm-1 | ||||
| DSP0 | 0.1 | 0 | 1 620 | 1 323 | — | — | |
| 0.2 | 0 | 1 618 | 1 323 | — | — | ||
| 0.4 | 0 | 1 620 | 1 323 | — | — | ||
| 0.6 | 0 | 1 618 | 1 323 | — | — | ||
| 0.8 | 0 | 1 620 | 1 323 | — | — | ||
| 1.0 | 0 | 1 620 | 1 323 | — | — | ||
| DSP1 | 0.1 | 0 | 1 620 | 1 320 | |||
| 0.2 | 39.58 | 1 620 | 1 319 | 1 643 | 1 360 | ||
| 0.4 | 92.91 | 1 639 | 1 327 | 1 643 | 1 329 | ||
| 0.6 | 100 | 1 641 | 1 329 | ||||
| 0.8 | 100 | 1 643 | 1 329 | ||||
| 1.0 | 100 | 1 647 | 1 329 | ||||
| DSP2 | 0.1 | 75.09 | 1 620 | 1 323 | 1 641 | 1 327 | |
| 0.2 | 95.92 | 1 620 | 1 323 | 1 641 | 1 329 | ||
| 0.4 | 100 | 1 643 | 1 327 | ||||
| 0.6 | 100 | 1 641 | 1 327 | ||||
| 0.8 | 100 | 1 643 | 1 327 | ||||
| 1.0 | 100 | 1 643 | 1 327 | ||||
| DSP3 | 0.1 | 92.09 | 1 624 | 1 323 | 1 641 | 1 329 | |
| 0.2 | 100 | 1641 | 1 329 | ||||
| 0.4 | 100 | 1 643 | 1 329 | ||||
| 0.6 | 100 | 1 643 | 1 327 | ||||
| 0.8 | 100 | 1 643 | 1 327 | ||||
| 1.0 | 100 | 1 643 | 1 327 | ||||
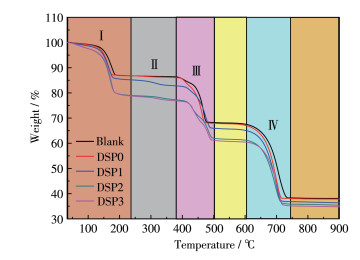
图 3为不同—COOH含量DSPs诱导形成的CaC2O4晶体的TGA曲线。
ρDSP=0.2 g·L-1.
在第Ⅰ阶段(100~200 ℃)的晶体失重归因于COM和COD的晶体结晶水的失去。DSP0组晶体的失重与没有多糖存在的空白组(只形成COM)基本一致,分别为12.3%和12.1%。但DSP1、DSP2和DSP3组晶体的失重比例依次为13.14%、18.06%和18.16%,说明各DSPs调控的晶体中结晶水的质量分数大小顺序为DSP3 > DSP2 > DSP1 > DSP0 > blank,即晶体中COD的质量分数大小顺序为DSP3 > DSP2 > DSP1 > DSP0,归因于COD含有2个结晶水,而COM只有1个结晶水。此外,—COOH越高的多糖调控的晶体失重温度越高,这是由于多糖中的—COOH与CaC2O4晶体表面的相互作用,使得DSPs晶体相对于纯COM具有更优异的稳定性[23]。
在第Ⅱ阶段(240~380 ℃)的失重归因于多糖分解[24-26]。DSP0、DSP1、DSP2和DSP3诱导形成的晶体在该阶段的失重百分比分别为1.54%、2.94%、7.96%和8.12%,即多糖掺入晶体中的比例。可见,羧基化程度最高的DSP3掺入到晶体的比例最高,且具有最高的分解温度,说明DSPs与晶体的特异性相互作用能力更强,结合更牢固。Souissi等[26]研究表明竹蛏多糖(Solen marginatus polysaccharide)的失重温度在200~600 ℃之间。El-Naggar等[27]对小球藻的可溶性多糖进行了热重分析,发现多糖的失重主要发生在240~650 ℃范围内。
在第Ⅲ阶段(380~510 ℃)的明显失重归因于CaC2O4晶体进一步失去一氧化碳(CO)和二氧化碳(CO2),最终分解物为CaO。
在第Ⅳ阶段(510~740 ℃),CaC2O4样品基本分解完成,最终残留重量百分比分别为38.86%、38.74%、37.71%、37.44%和37.26%,失重大小顺序与第Ⅰ、第Ⅱ阶段相反,归因于在这2个阶段失去了较高百分比的结晶水和多糖后,剩下的组分比例较低所致。
图 4是在0.2 g·L-1时4种多糖调控生成的CaC2O4晶体的SEM图像。没有多糖存在时,生成的晶体全部为COM晶体,其形貌不规则,且聚集现象严重;采用纳米测量仪统计的100个晶体尺寸为(0.77±0.41) μm。DSP0调控生成的晶体也只形成COM晶体,但其尺寸更小,为(0.44±0.35) μm。而DSP1、DSP2调控的晶体中同时存在COM与COD;DSP3只调控COD晶体形成,这些结果与PXRD检测的结果一致。即随着多糖中的—COOH含量越多,其诱导生成的COD晶体百分比越大,晶体尺寸则越小。

ρDSP=0.2 g·L-1.
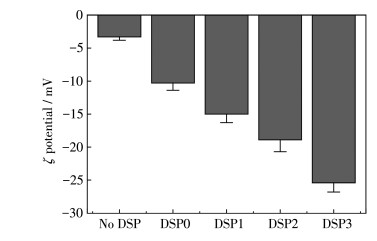
图 5为在0.2 g·L-1时不同DSPs调控形成的CaC2O4晶体的ζ电位。随着DSPs中—COOH含量从1.17%(DSP0)增加到7.45%(DSP1)、12.2%(DSP2)和17.7%(DSP3),晶体的ζ电位从-10.3 mV降低至-15.0、-18.9和-25.3 mV。晶体的ζ电位越低,表明晶体表面的负电荷密度越大,晶体间排斥力会越大,晶体聚集沉降的可能性越低,越有利于抑制肾结石的形成。
 下载:
导出CSV
下载:
导出CSV
| DSP | Weight loss/% | Remaining weight/% | |||
| First stage | Second stage* | Third stage | Fourth stage | ||
| Blank | 12.10 | — | 19.36 | 29.68 | 38.86 |
| DSP0 | 12.30 | 1.54 | 18.15 | 29.27 | 38.74 |
| DSP1 | 13.14 | 2.94 | 17.45 | 28.75 | 37.71 |
| DSP2 | 18.06 | 7.96 | 12.52 | 24.02 | 37.44 |
| DSP3 | 18.16 | 8.12 | 12.34 | 24.12 | 37.26 |
| *The weight loss in stage Ⅱ corresponds to the loss of polysaccharides, i.e., the mass fraction of polysaccharides in crystals. | |||||
尿微晶黏附在肾小管上皮细胞上是肾结石形成的关键步骤之一[28]。一方面,晶体黏附后进一步损伤细胞,另一方面,细胞损伤又加剧晶体的黏附[29-30],形成的恶性循环导致晶体在肾脏的滞留和沉积。
CD44是肾小管上皮细胞表达的一种黏附分子,在肾结石形成过程中发挥着重要的作用[31-32]。通过免疫荧光染色和半定量分析(图 6)评估了各组细胞的CD44表达量变化。可以看出,对照组细胞荧光强度最弱,即CD44表达量最低;nano-COM组细胞的CD44表达量最高。半定量结果显示CD44的表达量约为对照组16倍(图 6B),说明nano-COM的损伤促进了黏附蛋白CD44的表达。
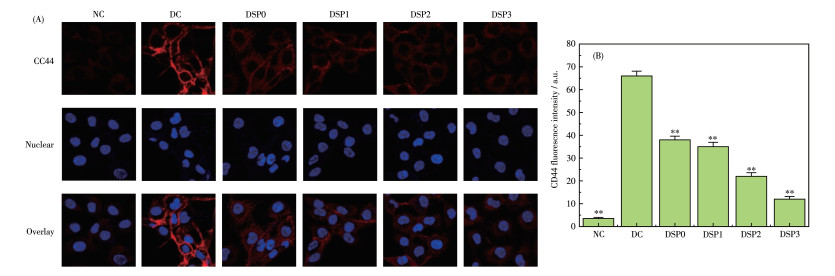
NC: normal control group, DC: crystal damage group, ρDSP=80 μg·mL-1, ρnano-COM=200 μg·mL-1, protection time: 24 h, scale bar: 10 μm, compared with DC group, **P < 0.01.
经不同DSPs保护后的细胞表面CD44表达量介于对照组和损伤组之间,其中—COOH含量最高的DSP3组的CD44表达量最接近对照组。多糖保护后CD44表达量降低,有利于减少CaC2O4晶体的黏附,降低成石风险。文献[33]报道,条斑紫菜多糖(PYPs)同样可以降低细胞的CD44表达,进而使得nano-COM对HK-2细胞的黏附受到抑制。
Annexin A2是一种Ca2+-磷脂结合蛋白,Annexin A2与各种肾脏疾病的发病机制和进展有关[34]。在肾结石形成过程中,Annexin A2可能介导晶体与肾小管上皮细胞黏附,促进晶体滞留。而特异性抗体对表面Annexin A2的阻断和Annexin A2下调可以降低CaC2O4晶体的黏附[35]。
如图 7所示,晶体损伤组细胞的Annexin A2表达量明显高于对照组,说明nano-COM介导了黏附蛋白Annexin A2表达的增加。经不同羧基含量的DSPs保护后,细胞的Annexin A2表达量逐渐下降,且随多糖中—COOH含量的增加而降低,即羧基化修饰后可以提高多糖对HK-2细胞的保护效果。Annexin A2表达量降低后,可以减少晶体的黏附,从而抑制肾结石的形成。
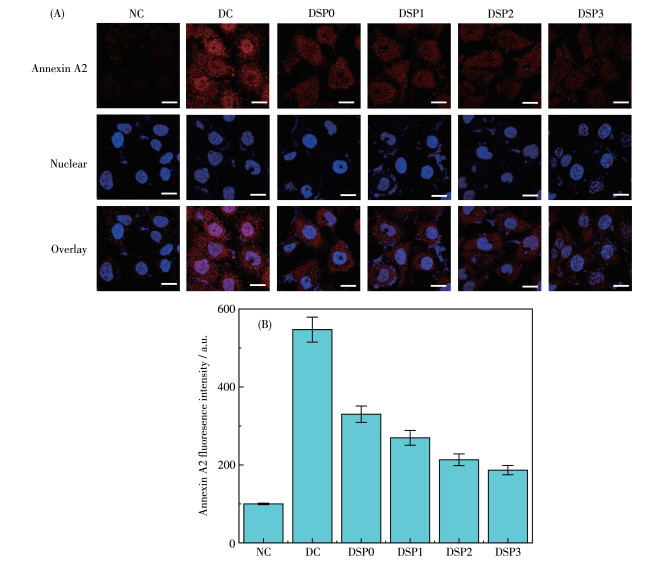
NC: normal control group, DC: crystal damage group, ρDSP=80 μg·mL-1, ρnano-COM=200 μg·mL-1, protection time: 24 h, scale bar: 10 μm, compared with DC group.
纳米颗粒与细胞相互作用的第一步发生在细胞表面,包括纳米颗粒与细胞膜的黏附以及与细胞受体的潜在相互作用。纳米颗粒对细胞的黏附可能会引发细胞的损伤及死亡[36-37]。图 8为采用共聚焦显微镜观察的各DSPs保护前后nano-COM晶体在细胞膜上的黏附,其中COM晶体采用FITC荧光标记。实验在4 ℃下进行,此时观察到的晶体都是黏附的晶体,因为晶体的内吞作用在4 ℃被抑制。图 8结果显示,正常组细胞表面黏附的COM晶体(绿色荧光)很少;而损伤组细胞黏附了大量晶体,且晶体聚集。相比之下,多糖保护组细胞黏附的晶体量比损伤组明显减少,且多糖中—COOH含量越高,黏附的晶体量越少。
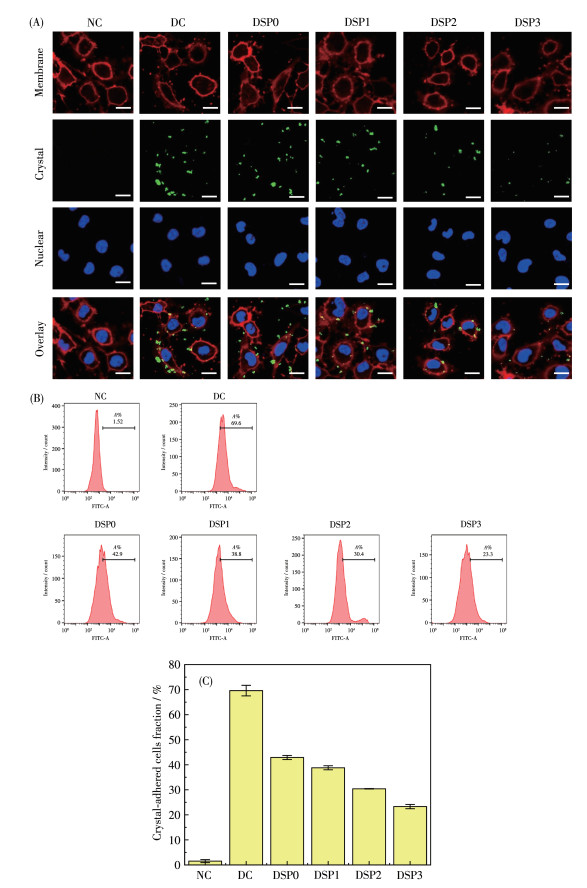
NC: normal control group, DC: crystal damage group, ρDSP=80 μg·mL-1, ρnano-COM=200 μg·mL-1, protection time: 24 h; scale bar: 20 μm.
同时采用流式细胞仪定量检测了各DSPs保护前后黏附nano-COM晶体的细胞百分比(图 8B)。损伤组细胞黏附晶体的细胞百分比(69.6%)显著高于对照组(1.52%)。各组细胞对晶体的黏附能力大小为:损伤组 > DSP0 > DSP1 > DSP2 > DSP3 > 对照组(图 8C),即经DSP3保护后的细胞抗晶体黏附的能力最强。
多糖上的羟基和羧基可以通过氢键缔合形成网络结构[38-39],这种结构在维持多糖的构象过程中发挥重要的作用[40-41]。本工作研究的广金钱草DSP0以葡萄糖为主,在羧基化修饰后的DSPs中由于带负电荷的羧基(—COO-)增加,其形成氢键和网络结构的能力增强。当具有网络结构的多糖与Ca2+作用时,Ca2+离子会取代网络中的部分氢原子,形成钙桥,并通过钙桥形成“蛋盒”结构[42-43],这种结构的配合物具有较好的稳定性,热分解温度比纯CaC2O4增高。TGA热重分析(图 3)表明随着多糖中—COOH含量增加,其调控形成的晶体失重温度升高。相关研究表明,以葡萄糖为主的多糖与Ca2+结合包含了2个层次上的分子相互作用:一是正负电荷静电作用,二是Ca2+与多糖的配位作用。而羧基化后多糖的这些作用进一步加强[42]。Cui等[44]通过制备香菇多糖-钙配合物(香菇多糖中有87.5%的D-葡萄糖),发现Ca2+离子增加了多糖间的分子作用力。
DSPs抑制COM形成与多糖-Ca2+之间的配位作用密切相关。DSPs含有大量带负电荷的羧基(—COO-),这些基团与Ca2+具有很强的配位能力,能够形成配合物。形成配合物后,由于多糖与体系中的Ca2+离子配位,体系中游离的Ca2+离子浓度降低,使得生成COM沉淀减少[43]。由于DSP1、DSP2和DSP3中—COOH含量从DSP0的1.17%增加至7.45%、12.2%和17.7%,因此修饰多糖与Ca2+离子的配位能力大大增强,更能抑制COM晶体的形成。我们推测DSPs与Ca2+离子的配合物结构如图 9所示。
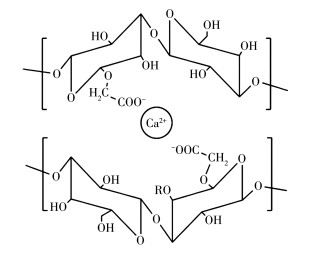
众多研究表明,多糖与Ca2+形成的配合物与反应物(CaCl2/多糖)物质的量之比、反应时间、pH、温度等因素相关[44-45];加上多糖结构较为复杂,存在主链和副链,且多糖的分子量大多是在一个范围内,因而难以确定配位反应中多糖与Ca2+离子的准确质量比,也难以计算出多糖-钙配合物的稳定常数。一般采用“配合物钙离子质量分数”或“多糖-钙配合物中钙的含量”来表达配位效果[46]。
一些文献将羧基化多糖的活性增强归因于离子基团通过扩展其构象及其与某些蛋白质相互作用[47-49]。Chen等[49]报道,羧基化多糖的相对延伸链构象有利于通过与巨噬细胞表面受体相互作用以激活免疫功能来增强抗肿瘤活性,其中—COOH基团在这种相互作用中起重要作用。
多糖中的—COOH含量增加后,其活性增强和调控CaC2O4结晶能力增强的原因:
第一,随着多糖的羧基化程度增加,即—COOH含量增加后,多糖的水溶性增强,其亲水亲油平衡值(HLB值)增加,乳化能力增强,从而提高了CaC2O4晶体的稳定性,抑制了CaC2O4晶体的沉积。
第二,富含酸性—COOH基的多糖可与游离的Ca2+离子结合生成可溶性配合物,且多糖中—COOH基含量越高,其与Ca2+配位的能力越强。体系中可溶性Ca2+浓度增大后,可以增加CaC2O4晶体的过饱和度,减少与C2O42-结合的Ca2+量,即减少了CaC2O4的沉积,有利于抑制草酸结石的形成。
第三,多糖抑制CaC2O4晶体聚集。从SEM图像(图 4)可知,没有多糖存在或多糖中—COOH基含量较低时,生成的CaC2O4晶体聚集程度较大。多糖吸附在晶体表面后,晶体表面的ζ电位变低(图 5),且随着多糖中—COOH基含量增加,ζ电位更低,这可以增加晶体间的排斥力,从而抑制晶体的聚集。聚集的晶体不但增大了对肾上皮细胞的损伤,而且更容易滞留在体内,这均会增加结石形成的危险。
第四,多糖在溶液里形成聚阴离子,可吸附在带正电荷的COM晶体的表面,导致晶体生长的缺陷,阻止游离粒子渗入,从而抑制COM的生长。
第五,多糖促进COD生成。多糖中的酸性—COOH可通过静电引力吸附溶液里的大量Ca2+离子,在多糖表面处的阳离子浓度比体相中显著提高,从而使阴、阳离子之比(Rion=cCa2+/cC2O42-)偏离晶体的化学计量比。由于Ca2+离子过量(Rion > 1.0)有利于COD晶体形成,而C2O42-离子过量(Rion < 1.0)有利于COM晶体形成[41],因此,多糖吸附Ca2+离子后促进了COD的形成。此外,多糖的—COOH与Ca2+离子配位后,使得多糖表面形成高能界面,同时配位后的Ca2+离子自由度降低,导致钙能态增加,这种高能界面和高钙能态均会促进热力学亚稳定的COD形成[51]。由于DSP3中的—COOH含量最高,对溶液中Ca2+离子的配位能力最强,因此,DSP3更容易诱导COD的形成。由于COD晶体表面的吸附位点较COM少,且COM晶体表面带更多正电荷,与带负电荷的受损伤肾上皮细胞黏附力显著大于COD晶体[50],因此,COD相对容易随尿液排出体外。将尿液中的COM转化为COD,可减小肾结石形成的风险。故羧基含量高的多糖抑制尿结石的效果会更好。
第六,随着多糖中—COOH基含量增加,生成的COD更圆钝(图 4),圆钝的晶体也更容易随尿液排出体外。多糖抑制COM形成和减少COM黏附的机制图 10所示。
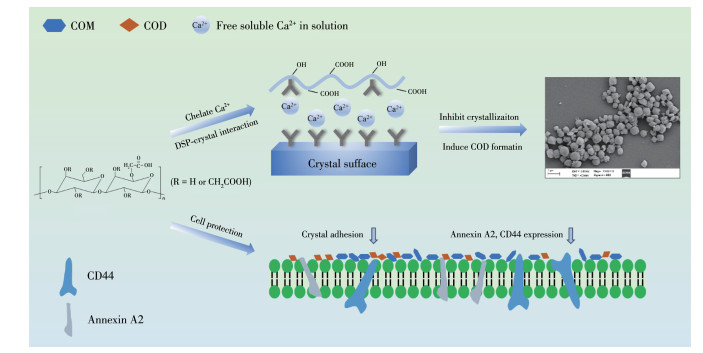
羧基化修饰的广金钱草多糖DSPs(—COOH含量为1.17%~17.7%)能有效抑制与细胞黏附能力更强的COM形成,诱导容易排出体外的COD生长,并抑制CaC2O4晶体的聚集,降低细胞表面黏附分子表达,减少nano-COM在细胞表面的黏附量,从而有利于抑制CaC2O4结石形成。DSPs的抑制活性与其—COOH含量成正相关。—COOH含量最高的DSP3具有最大的抑制CaC2O4结石形成的潜力。
Siener R, Herwig H, Rüdy J, Schaefer R M, Lossin P. Urinary stone composition in Germany: Results from 45, 783 stone analyses[J]. World J. Urol., 2022, 40(7): 1813-1820. doi: 10.1007/s00345-022-04060-w
Tian Y, Han G, Qu R, Xiao C. Major and trace elements in human kidney stones: A preliminary investigation in Beijing, China[J]. Minerals, 2022, 12(5): 512. doi: 10.3390/min12050512
Onal E G, Tekgul H. Assessing kidney stone composition using smartphone microscopy and deep neural networks[J]. BJUI Compass, 2022, 3(4): 310-315. doi: 10.1002/bco2.137
Van de Perre E, Bazin D, Estrade V, Bouderlique E, Wissing K M, Daudon M, Letavernier E. Randall's plaque as the origin of idiopathic calcium oxalate stone formation: An update[J]. C. R. Chim., 2021, 25(S1): 373-391.
Maruyama M, Sawada K P, Tanaka Y, Okada A, Momma K. Quantitative analysis of calcium oxalate monohydrate and dihydrate for elucidating the formation mechanism of calcium oxalate kidney stones[J]. PloS One, 2023, 18(3): e0282743. doi: 10.1371/journal.pone.0282743
Song Q L, Song C, Chen X, Xiong Y H, Li L J, Liao W B, Xue L J, Yang S X. FKBP5 deficiency attenuates calcium oxalate kidney stone formation by suppressing cell-crystal adhesion, apoptosis and macrophage M1 polarization via inhibition of NF-κB signaling[J]. Cell. Mol. Life Sci., 2023, 80(10): 301. doi: 10.1007/s00018-023-04958-7
Kuang S D, Liu L M, Hu Z R, Luo M, Fu X Y, Lin C X, He Q H. A review focusing on the benefits of plant-derived polysaccharides for osteoarthritis[J]. Int. J. Biol. Macromol., 2023, 228: 582-593. doi: 10.1016/j.ijbiomac.2022.12.153
Zhu Y Y, Feng X W, Guo J H, Wang L, Guo X D, Zhu X Z. A review of extraction, purification, structural properties and biological activities of legumes polysaccharides[J]. Front. Nutr., 2022, 9: 1021448. doi: 10.3389/fnut.2022.1021448
陈宇, 郑彦楷, 李大彪, 邢媛媛. 植物多糖对内质网应激介导的动物细胞凋亡、炎症和氧化损伤的影响及其作用机制[J]. 动物营养学报, 2023,35,(12): 7641-7647. doi: 10.12418/CJAN2023.694CHEN Y, ZHENG Y K, LI D B, XING Y Y. Effects of plant polysaccharides on endoplasmic reticulum stress-mediated apoptosis, inflammation and oxidative damage of animal cells and their mechanisms[J]. Chinese Journal of Animal Nutrition, 2023, 35(12): 7641-7647. doi: 10.12418/CJAN2023.694
Xie L M, Shen M Y, Wang Z J, Xie J H. Structure, function and food applications of carboxymethylated polysaccharides: A comprehensive review[J]. Trends Food Sci. Technol., 2021, 118: 539-557. doi: 10.1016/j.tifs.2021.09.016
Liu F, Liu Y, Feng X, Ibrahim S A, Huang W. Structure characterization and in vitro immunomodulatory activities of carboxymethyl pachymaran[J]. Int. J. Biol. Macromol., 2021, 178: 94-103. doi: 10.1016/j.ijbiomac.2021.02.046
Zhao T, Guo Y C, Yan S Y, Li N, Ji H C, Hu Q H, Zhang M, Li Q, Gao H, Yang L Q, Wu X Y. Preparation, structure characterization of carboxymethylated schisandra polysaccharides and their intervention in immunotoxicity to polychlorinated biphenyls[J]. Process Biochem., 2022, 115: 30-41. doi: 10.1016/j.procbio.2022.02.005
Kagimura F Y, da Cunha M A, Theis T V, Malfatti C R, Dekker R F, Barbosa A M, Teixeira S D, Salome K. Carboxymethylation of (1→6)-β-glucan (lasiodiplodan): Preparation, characterization and antioxidant evaluation[J]. Carbohydr. Polym., 2015, 127: 390-399. doi: 10.1016/j.carbpol.2015.03.045
Wotschadlo J, Liebert T, Heinze T, Wagner K, Schnabelrauch M, Dutz S, Müller R, Steiniger F, Schwalbe M, Kroll T C, Höffken K, Buske N, Clement J H. Magnetic nanoparticles coated with carboxymethylated polysaccharide shells—Interaction with human cells[J]. J. Magn. Magn. Mater., 2009, 321(10): 1469-1473. doi: 10.1016/j.jmmm.2009.02.069
Lai Y C, Zheng H X, Sun X, Lin J, Li Q Y, Huang H N, Hou Y, Zhong H Z, Zhang D F, Tang F C, He Z H. The advances of calcium oxalate calculi associated drugs and targets[J]. Eur. J. Pharmacol., 2022, 935: 175324. doi: 10.1016/j.ejphar.2022.175324
张会香, 汤霞利, 林军全, 陈海珊, 林伟国, 梁筱彬, 李霞, 关媛. 广金钱草不同极性萃取物体外降脂及降血糖活性的比较[J]. 现代食品科技, 2023,39,(12): 192-198. ZHANG H X, TANG X L, LIN Q J, CHEN H S, LIN W G, LIANG X B, LI X, GUAN Y. Comparison of the in vitro lipid-lowering and hypoglycemic activities of Desmodium styracifolium extracts with different polarities[J]. Modern Food Science and Technology, 2023, 39(12): 192-198.
Li X, Chen C, Zhang T J, Ding N, Zheng P Y, Yang M. Comparative pharmacokinetic studies of five C-glycosylflavones in normal and urolithiasis model rats following administration of total flavonoids from Desmodium styracifolium by liquid chromatography-tandem mass spectrometry[J]. J. Sep. Sci., 2022, 45(15): 2901-2913. doi: 10.1002/jssc.202200010
Li X, Liu C X, Liang J, Zhou L, Li J, Chen H Y, Jiang T M, Guang Y, Khoo H E. Antioxidative mechanisms and anticolitic potential of Desmodium styracifolium (Osb.) Merr. in DSS-induced colitic mice[J]. J. Funct. Foods, 2022, 93: 105077. doi: 10.1016/j.jff.2022.105077
Liu Q, Chen J, Liao Z M, Hu S W, Xu J, Yan W, Zeng L Q, Zhang Z M, Gao W X. Randomized, double-blind, placebo parallel controlled clinical study on the treatment of ureteral calculi with total flavonoids of Desmodium styracifolium[J]. J. Clin. Urol., 2023, 38(4): 246-250.
Tang G H, Liu J H, Sun X Y, Ouyang J M. Carboxymethylation of Desmodium styracifolium polysaccharide and its repair effect on damaged HK-2 cells[J]. Oxidative Med. Cell. Longev., 2022, : 2082263.
Huang L S, Sun X Y, Gui Q, Ouyang J M. Effects of plant polysaccharides with different carboxyl group contents on calcium oxalate crystal growth[J]. CrystEngComm, 2017, 19(32): 4838-4847. doi: 10.1039/C7CE00983F
Donnet M, Jongen N, Lemaitre J, Bowen P. New morphology of calcium oxalate trihydrate precipitated in a segmented flow tubular reactor[J]. J. Mater. Sci. Lett., 2000, 19: 749-750. doi: 10.1023/A:1006771428827
Zhu P Z, Xu J D, Sahar N, Morris M D, Kohn D H, Ramamoorthy A. Time-resolved dehydration-induced structural changes in an intact bovine cortical bone revealed by solid-state NMR spectroscopy[J]. J. Am. Chem. Soc., 2009, 131(47): 17064-17065. doi: 10.1021/ja9081028
Fang W C, Zhang H, Yin J W, Yang B G, Zhang Y B, Li J J, Yao F L. Hydroxyapatite crystal formation in the presence of polysaccharide[J]. Cryst. Growth. Des., 2016, 16(3): 1247-1255. doi: 10.1021/acs.cgd.5b01235
Zhang W J, Huang J, Wang W, Li Q, Chen Y, Feng W W, Zheng D H, Zhao T, Mao G H, Yang L Q, Wu X Y. Extraction, purification, characterization and antioxidant activities of polysaccharides from Cistanche tubulosa[J]. Int. J. Biol. Macromol., 2016, 93: 448-458. doi: 10.1016/j.ijbiomac.2016.08.079
Souissi N, Boughriba S, Abdelhedi O, Hamdi M, Jridi M, Li S, Nasri M. Extraction, structural characterization, and thermal and biomedical properties of sulfated polysaccharides from razor clam Solen marginatus[J]. RSC Adv., 2019, 9(20): 11538-11551. doi: 10.1039/C9RA00959K
El-Naggar N E A, Hussein M H, Shaaban-Dessuuki S A, Dalal S R. Production, extraction and characterization of Chlorella vulgaris soluble polysaccharides and their applications in AgNPs biosynthesis and biostimulation of plant growth[J]. Sci. Rep., 2020, 10(1): 3011. doi: 10.1038/s41598-020-59945-w
Li X R, Chen S Y, Feng D M, Fu Y Q, Wu H, Lu J Z, Bao J S. Calcium-sensing receptor promotes calcium oxalate crystal adhesion and renal injury in Wistar rats by promoting ROS production and subsequent regulation of PS ectropion, OPN, KIM-1, and ERK expression[J]. Ren. Fail., 2021, 43(1): 465-476. doi: 10.1080/0886022X.2021.1881554
Peerapen P, Thongboonkerd V. Differential bound proteins and adhesive capabilities of calcium oxalate monohydrate crystals with various sizes[J]. Int. J. Biol. Macromol., 2020, 163: 2210-2223. doi: 10.1016/j.ijbiomac.2020.09.085
杨雄, 靳潇潇, 何文强. 肾上皮细胞损伤与肾结石Randall斑块形成[J]. 基础医学与临床, 2022,42,(9): 1454-1458. YANG X, JIN X X, HE W Q. Injury of renal epithelial cell and the formation of Randall's plaque in kidney stones[J]. Basic and Clinical Medicine, 2022, 42(9): 1454-1458.
Ying Q, Liu G X, Zhou W J, Lan J H, Du J H, Tang X F, Xu X H. The rs13347 polymorphism of the CD44 gene is associated with the risk of kidney stones disease in the Chinese Han population of northeast Sichuan, China[J]. Comput. Math. Methods Med., 2022, : 6481260.
Wang B H, He G F, Xu G, Wen J M, Yu X. miRNA-34a inhibits cell adhesion by targeting CD44 in human renal epithelial cells: Implications for renal stone disease[J]. Urolithiasis, 2020, 48: 109-116. doi: 10.1007/s00240-019-01155-9
Zhang H, Sun X Y, Chen X W, Ouyang J M. Degraded Porphyra yezoensis polysaccharide protects HK-2 cells and reduces nano-COM crystal toxicity, adhesion and endocytosis[J]. J. Mater. Chem. B, 2020, 8(32): 7233-7252. doi: 10.1039/D0TB00360C
Lin L, Hu K B. Annexin A2 and kidney diseases[J]. Front. Cell Dev. Biol., 2022, 10: 974381. doi: 10.3389/fcell.2022.974381
Peerapen P, Boonmark W, Thongboonkerd V. Trigonelline prevents kidney stone formation processes by inhibiting calcium oxalate crystallization, growth and crystal-cell adhesion, and downregulating crystal receptors[J]. Biomed. Pharmacother., 2022, 149: 112876. doi: 10.1016/j.biopha.2022.112876
Francia V, Montizaan D, Salvati A. Interactions at the cell membrane and pathways of internalization of nano-sized materials for nanomedicine[J]. Beilstein J. Nanotechnol., 2020, 11(1): 338-353.
Septiadi D, Crippa F, Moore T L. Nanoparticle-cell interaction: A cell mechanics perspective[J]. Adv. Mater., 2018, 30(19): e1704463. doi: 10.1002/adma.201704463
Wang J Q, Yin J Y, Nie S P, Xie M Y. A review of NMR analysis in polysaccharide structure and conformation: Progress, challenge and perspective[J]. Food Res. Int., 2021, 143: 110290. doi: 10.1016/j.foodres.2021.110290
Yu Y, Tyrikos-Ergas T, Zhu Y, Fittolani G, Bordoni V. Systematic hydrogen-bond manipulations to establish polysaccharide structure-property correlations[J]. Angew. Chem. Int. Ed., 2019, 131(37): 13261-13266. doi: 10.1002/ange.201906577
Ziemniak M, Zawadzka-Kazimierczuk A, Pawlędzio S, Malinska M, Sołtyka M, Trzybiński D, Pająk B. Experimental and computational studies on structure and energetic properties of halogen derivatives of 2-deoxy-D-glucose[J]. Int. J. Mol. Sci., 2021, 22(7): 3720. doi: 10.3390/ijms22073720
Meng Y, Lyu F Z, Xu X J, Zhang L N. Recent advances in chain conformation and bioactivities of triple-helix polysaccharides[J]. Biomacromolecules, 2020, 21(5): 1653-1677. doi: 10.1021/acs.biomac.9b01644
王雪松, 方积年. 糖类的金属络合物[J]. 化学通报, 2004,67,(12): 883-890. doi: 10.3969/j.issn.0441-3776.2004.12.003WANG X S, FANG J N. Sugar-metal complex[J]. Chem. Bull., 2004, 67(12): 883-890. doi: 10.3969/j.issn.0441-3776.2004.12.003
Wang W J, Shen M Y, Jiang L, Song Q Q, Liu S C, Xie M Y, Xie J H. Rheological behavior, microstructure characterization and formation mechanism of Mesona blumes polysaccharide gels induced by calcium ions[J]. Food Hydrocolloids, 2019, 94: 136-143. doi: 10.1016/j.foodhyd.2019.03.014
Cui Y J, Yan H D, Zhang X W. Preparation of Lentinula edodes polysaccharide‑calcium complex and its immunoactivity[J]. Biosci. Biotechnol. Biochem., 2015, 79(10): 1619-1623. doi: 10.1080/09168451.2015.1044930
李海鹰, 杨祥禾, 魏珍珍, 贾蓓, 杨文智. 羧甲基普鲁兰多糖螯合钙的制备及其功效评价[J]. 食品科学, 2021,42,(15): 143-149. doi: 10.7506/spkx1002-6630-20200724-330LI H Y, YANG X H, WEI Z Z, JIA B, YANG W Z. Preparation and evaluation of carboxymethyl pullulan-calcium complex in vivo[J]. Food Sci., 2021, 42(15): 143-149. doi: 10.7506/spkx1002-6630-20200724-330
Pan X H, Wu S H, Yan Y J, Chen X, Guan J T, Bao Y P, Xiong X M, Liu L. Rice bran polysaccharide-metal complexes showed safe antioxidant activity in vitro[J]. Int. J. Biol. Macromol., 2019, 126: 934-940. doi: 10.1016/j.ijbiomac.2018.12.265
Wu J Y, Zhang Y H, Ye L, Wang C L. The anti-cancer effects and mechanisms of lactic acid bacteria exopolysaccharides in vitro: A review[J]. Carbohydr. Polym., 2021, 253: 117308. doi: 10.1016/j.carbpol.2020.117308
Liu Y T, Duan X Y, Zhang M Y, Li C, Zhang Z Q, Hu B, Liu A P, Li Q, Chen H, Tang Z Z, Wu W J, Chen D W. Extraction, structure characterization, carboxymethylation and antioxidant activity of acidic polysaccharides from Craterellus cornucopioides[J]. Ind. Crop. Prod., 2021, 159: 113079. doi: 10.1016/j.indcrop.2020.113079
Chen X Y, Zhang L, Cheung P C K. Immunopotentiation and antitumor activity of carboxymethylated-sulfated β-(1→3)-D-glucan from Poria cocos[J]. Int. Immunopharmacol., 2010, 10(4): 398-405. doi: 10.1016/j.intimp.2010.01.002
Jung T S, Kim W S, Choi C K. Biomineralization of calcium oxalate for controlling crystal structure and morphology[J]. Mater. Sci. Eng. C, 2004, 24(1/2): 31-33.
Gomes D L, Melo K R T, Queiroz M F, Batista L, Santos P, Costa M, Lima J, Camara R, Costa L, Rocha H. In vitro studies reveal antiurolithic effect of antioxidant sulfated polysaccharides from the green seaweed Caulerpa cupressoides var flabellata[J]. Mar. Drugs, 2019, 17(6): 326. doi: 10.3390/md17060326

图 1 不同质量浓度、不同—COOH含量DSPs调控形成的CaC2O4晶体的PXRD图: (A) DSP0、(B) DSP1、(C) DSP2、(D) DSP3; (E) 晶体中COD的质量分数
Figure 1 PXRD patterns of CaC2O4 crystals formed by the regulation of DSPs at different concentrations and different—COOH contents: (A) DSP0, (B) DSP1, (C) DSP2, (D) DSP3; (E) Mass fractions of COD in the crystals
*means the characteristic diffraction peak of COM, and # means the characteristic diffraction peak of COD.

图 2 不同质量浓度、不同—COOH含量DSPs调控形成的CaC2O4晶体的FTIR谱图: (A) DSP0、(B) DSP1、(C) DSP2、(D) DSP3
Figure 2 FTIR spectra of CaC2O4 crystals formed by the regulation of DSPs at different concentrations and different —COOH contents: (A) DSP0, (B) DSP1, (C) DSP2, (D) DSP3

图 3 不同—COOH含量DSPs诱导形成的CaC2O4晶体的TGA曲线
Figure 3 TGA curves of CaC2O4 crystals induced by DSPs with different —COOH contents
ρDSP=0.2 g·L-1.

图 4 不同—COOH含量DSPs诱导形成的CaC2O4晶体的SEM图
Figure 4 SEM images of CaC2O4 crystals induced by DSPs with different —COOH contents
ρDSP=0.2 g·L-1.

图 5 在0.2 g·L-1 DSPs存在下形成的CaC2O4晶体的ζ电位
Figure 5 ζ potentials of CaC2O4 crystals formed by the regulation of 0.2 g·L-1 DSPs

图 6 DSPs保护前后HK-2细胞表面CD44表达量变化: (A) CD44荧光图; (B) CD44表达量统计柱状图
Figure 6 Changes in the expression levels of CD44 before and after DSPs protecting the HK-2 cells: (A) CD44 fluorescence picture; (B) CD44 expression statistics histogram
NC: normal control group, DC: crystal damage group, ρDSP=80 μg·mL-1, ρnano-COM=200 μg·mL-1, protection time: 24 h, scale bar: 10 μm, compared with DC group, **P < 0.01.

图 7 DSPs保护前后HK-2细胞的Annexin A2表达量变化: (A)Annexin A2荧光图; (B) Annexin A2表达定量柱状图
Figure 7 Changes in the expression levels of Annexin A2 before and after DSPs protecting the HK-2 cells: (A) Annexin A2 fluorescence picture; (B) Annexin A2 expression quantitative histogram
NC: normal control group, DC: crystal damage group, ρDSP=80 μg·mL-1, ρnano-COM=200 μg·mL-1, protection time: 24 h, scale bar: 10 μm, compared with DC group.

图 8 不同—COOH含量DSPs保护前后nano-COM晶体在HK-2细胞表面的黏附变化: (A) 共聚焦显微镜观察nano-COM在细胞膜上的黏附; (B) 流式细胞仪定量检测黏附晶体的细胞百分比; (C) nano-COM黏附量柱状图
Figure 8 Adhesion changes of nano-COM crystals on HK-2 cell surface before and after protection by DSPs with different —COOH content: (A) confocal microscopy to observe the adhesion of nano-COM crystals; (B) percentage of cells adhering to the crystals measured by flow cytometry; (C) nano-COM adhesion statistics histogram
NC: normal control group, DC: crystal damage group, ρDSP=80 μg·mL-1, ρnano-COM=200 μg·mL-1, protection time: 24 h; scale bar: 20 μm.

图 9 DSPs与Ca2+离子配合物的可能结构
Figure 9 Possible structures of the complex of DSPs and Ca2+ ion

图 10 DSPs抑制COM形成和减少COM黏附机制示意图
Figure 10 Schematic diagram of the mechanism of DSPs inhibiting the growth of COM and reducing the adhesion of nano-COM crystals
表 1 不同质量浓度、不同—COOH含量DSPs诱导形成的CaC2O4晶体中COD的质量分数及FTIR谱图中νas(COO-)和νs(COO-)吸收峰
Table 1. Mass fraction of COD in CaC2O4 crystals induced by DSPs with different mass concentrations and contents of —COOH, as well as the absorption peaks of νas(COO-) and νs(COO-) in FTIR spectra
| DSP | ρDSP/(g·L-1) | wCOD/% | COM | COD | |||
| νas(COO-)/cm-1 | νs(COO-)/cm-1 | νas(COO-)/cm-1 | νs(COO-)/cm-1 | ||||
| DSP0 | 0.1 | 0 | 1 620 | 1 323 | — | — | |
| 0.2 | 0 | 1 618 | 1 323 | — | — | ||
| 0.4 | 0 | 1 620 | 1 323 | — | — | ||
| 0.6 | 0 | 1 618 | 1 323 | — | — | ||
| 0.8 | 0 | 1 620 | 1 323 | — | — | ||
| 1.0 | 0 | 1 620 | 1 323 | — | — | ||
| DSP1 | 0.1 | 0 | 1 620 | 1 320 | |||
| 0.2 | 39.58 | 1 620 | 1 319 | 1 643 | 1 360 | ||
| 0.4 | 92.91 | 1 639 | 1 327 | 1 643 | 1 329 | ||
| 0.6 | 100 | 1 641 | 1 329 | ||||
| 0.8 | 100 | 1 643 | 1 329 | ||||
| 1.0 | 100 | 1 647 | 1 329 | ||||
| DSP2 | 0.1 | 75.09 | 1 620 | 1 323 | 1 641 | 1 327 | |
| 0.2 | 95.92 | 1 620 | 1 323 | 1 641 | 1 329 | ||
| 0.4 | 100 | 1 643 | 1 327 | ||||
| 0.6 | 100 | 1 641 | 1 327 | ||||
| 0.8 | 100 | 1 643 | 1 327 | ||||
| 1.0 | 100 | 1 643 | 1 327 | ||||
| DSP3 | 0.1 | 92.09 | 1 624 | 1 323 | 1 641 | 1 329 | |
| 0.2 | 100 | 1641 | 1 329 | ||||
| 0.4 | 100 | 1 643 | 1 329 | ||||
| 0.6 | 100 | 1 643 | 1 327 | ||||
| 0.8 | 100 | 1 643 | 1 327 | ||||
| 1.0 | 100 | 1 643 | 1 327 | ||||
 下载: 导出CSV
下载: 导出CSV
表 2 0.2 g·L-1的DSPs诱导形成的CaC2O4晶体的失重百分比
Table 2. Weight loss percentage of CaC2O4 crystals induced by DSPs at 0.2 g·L-1
| DSP | Weight loss/% | Remaining weight/% | |||
| First stage | Second stage* | Third stage | Fourth stage | ||
| Blank | 12.10 | — | 19.36 | 29.68 | 38.86 |
| DSP0 | 12.30 | 1.54 | 18.15 | 29.27 | 38.74 |
| DSP1 | 13.14 | 2.94 | 17.45 | 28.75 | 37.71 |
| DSP2 | 18.06 | 7.96 | 12.52 | 24.02 | 37.44 |
| DSP3 | 18.16 | 8.12 | 12.34 | 24.12 | 37.26 |
| *The weight loss in stage Ⅱ corresponds to the loss of polysaccharides, i.e., the mass fraction of polysaccharides in crystals. | |||||
下载: 导出CSV

 扫一扫看文章
扫一扫看文章

扫一扫关注我们

 下载:
下载: