表 1
常用的视紫红质离子通道蛋白(ChRs)
Table 1.
Commonly used channelrhodopsins (ChRs)

精准调控生物分子以及生理过程,对于理解疾病的发生发展和实施有效的治疗具有重大意义[1-2]。而离子通道作为细胞膜的主要成分,对于神经、肌肉等其它系统的电生理信号的传导和整合起到关键作用,它的活化或功能障碍会影响正常生理及病理进程,比如大脑思维,肌肉收缩和离子通道病[3-5]。目前,常用于离子通道调控的策略有以下几种:(1)化学分子作用,如离子通道激活剂或阻断剂;(2)基因工程干预,如特异性地表达、敲除或沉默目的基因来影响通道蛋白的活性;(3)物理电刺激,作用于特定的电压门控离子通道[6-9]。尽管这些方法都取得了一定的进展,但在实际应用中依然存在着许多挑战。例如,化学药物随血液循环的非识别性积累及作用难以避免,这极大地限制了调控的空间分辨率[10]。再者,化学或遗传扰动的不可逆性也阻碍了实际调控的时间准确性[7]。此外,尽管电学模式的物理刺激,尤其是对于深部大脑刺激而言,显示出很好的时空准确性和应用价值。但是,实际操作中需要高侵入性地在深层脑组织植入电极或芯片,会造成潜在的临床不良反应[11-12]。
因此,迫切需要开发精度高、损伤小的调控膜离子通道的有效技术。近年来,使用光来控制生物分子和生理过程已引起了广泛关注[13-15]。其中一种新兴的被称作光遗传学的生物技术被应用于神经科学领域,并且能高选择性地,甚至在毫秒级的时间分辨率下实现神经功能的操控[16-18]。更重要的是,通过基因工程对光敏视紫红质离子通道蛋白进行改造,使其在离子特异性以及光谱响应性2个方面进一步多样化。这为推动光遗传学在更复杂的生物体系应用的提供了机会,不仅在体外单细胞水平,还在自由活动动物上对脑回路介导的行为学进行调控[19-22]。然而,尽管这些技术取得了令人瞩目的成就,但目前报道的光敏感视紫红质蛋白,或其它用于膜通道调节的光遗传学工具主要是在可见光区工作。由于可见光组织穿透性不足,生物体吸收和散射严重,这些因素极大地限制了光遗传学在体内应用[18, 23-25]。虽然有研究报道通过光学纤维或微型发光二极管植入可以做到深层脑组织刺激,但这种高度侵入性的手段会引起一定的安全隐患[26-27]。由此,开发新的光遗传学技术,使其能够在活体条件下无(微)创地、有效地调控深层部位膜通道具有重要意义。
值得注意的是,科学家们目前为实现体内有效的光遗传学刺激做出了多方面努力,其中将光敏离子通道蛋白激发波长移至近红外窗口(> 700 nm)被认为有利于更深的组织穿透能力[24, 28-29]。比如通过基因工程化的策略,目前不同突变视紫红质离子通道蛋白的响应光谱已经从蓝光红移到黄光,甚至红光区域,但这些光遗传学体系仍局限在可见光波长范围内(表 1)[30-31]。此外,通过使用近红外光响应纳米材料作为光转换器,进一步原位刺激光遗传学工具,可以增强光穿透能力进而调控离子通道的活性[32-34]。其中,镧系元素掺杂的上转换纳米粒子作为独特的光学材料被选为潜在的光纳米转换器。该材料具有将近红外光(如980或808 nm)转换为紫外、可见或近红外区域的多种发射的性能(表 2),鉴于其较少的散射和更深的组织渗透深度,已被广泛地应用于生物成像和纳米医学研究领域[35-39]。基于此,近年来人们将镧系元素掺杂的上转换纳米粒子与各种光敏离子通道蛋白结合,实现了近红外上转换的光遗传学调控,并取得了显著的研究成果[32, 40-41]。
 下载:
导出CSV
下载:
导出CSV
| ChRsa | λmax (activation) / nm | Function and origin | Ref. |
| ChR2 | 480 | Excitatory cation channel; Native from C. reinhardtii | [42] |
| VChR1 | 545 | Red-shifted, excitatory cation channel; Native from V. carteri | [31] |
| ChIEF | 450 | Fast kinetics, excitatory cation channel; Chimera of ChR1 and VChR1 | [43] |
| ChETA | 470 | Fast kinetics, excitatory cation channel; Substitutions in ChR2 | [44] |
| Step function opsins | 470 (on) 590 (off) | Excitatory cation channel; Single mutations in ChR2 | [45] |
| Chronos | 500 | Fast kinetics, excitatory cation channel; Native from Stigeoclonium helveticum | [46] |
| Chrimson | 590 | Red-shifted, excitatory cation channel; Native from Chlamydomonas noctigama | |
| C1V1 | 540 | Red-shifted, excitatory cation channel; Chimera of ChR1 and VChR1 | [47] |
| NpHR | 590 | Inhibitory chloride pump; Native from N. pharaonis | [48] |
| ChloC | 480 | Inhibitory anion channel; Mutagenesis of ChR2 | [49] |
| GtACR2 | 520 | Inhibitory anion pump; Native from G. theta | [50] |
| ArCh | 570 | Inhibitory proton pump; Native from H. sodomense | [51] |
| a ChR2, channelrhodopsin-2; ReaChR, red-shifted channelrhodopsin; NpHR, natronomonas halorhodopsin; ChloC and GtACR2, chloride-conducting channelrhodopsin; Arch, archaerhodopsin; other mutations of channelrhodopsins (VChR1, C1V1, ChIEF, ChETA, mChR1, PsChR)[52] | |||
下载:
导出CSV
| UCNPs | Ln3+ | λex / nm | λem / nm |
| NaYF4:Yb/Ln3+ or NaGdF4:Yb/Ln3+ | Tm3+ | 980 | 450, 475, 800 |
| Er3+ | 545, 665 | ||
| Pr3+ | 480, 606 | ||
| Tb3+ | 480~650 | ||
| Eu3+ | 570~720 | ||
| Sm3+ | 601 | ||
| Ho3+ | 540 | ||
| Dy3+ | 486, 575 | ||
| NaYF4:Yb/Nd3+/Ln3+ or NaYF4:Yb/Nd3+/Ln3+/Dye | Ln3+ | 980 or 808 | As above |
因此,我们聚焦于生物医学研究中将近红外上转换纳米转换器用于光遗传学调控的最新成果。首先,我们对特定功能的上转换纳米平台与光敏离子通道蛋白结合的策略进行了总结;其次,详细地介绍了上转换光遗传学的广泛应用以及可改进的技术;最后,关于进一步推进该技术向临床转化并克服当前挑战提出了建议和展望。
通过匹配不同光敏视蛋白的激活波长,选择适当的上转换纳米粒子作为近红外光转换器,这种组合策略可以灵活地实现特定离子通道的激活。2011年,Deisseroth等[53]在一项专利申请中首先提出了这个理念,并列举了各种上转换纳米粒子与细胞膜上光响应视蛋白表达的神经元组合并进行光调控的方法。在2013年,Han等在一项基金申请里介绍了用于体内神经元无光纤操控的上转换光遗传学设计。此后在2015年,不同研究团队分别报道了在体外体内成功地实现了上转换光遗传学调控的研究[54-55]。
例如,Yawo等[40]使用蓝色发光(发射峰在450和480 nm)的上转换纳米材料NaYF4:Yb/Sc/Tm@ NaYF4,在近红外976 nm激光照射下,可以有效地激活PsChR离子通道,并产生明显的动作电位;另外,利用发出绿光(550 nm)的NaYF4:Sc/Yb/Er作为近红外光转换器,进而可用于激活表达细胞中的C1V1或mVChR1离子通道蛋白(图 1)。进一步地研究还证明,结合上转换纳米粒子的光遗传学调控效果显示了刺激的时间和功率依赖性。但是,该研究仍有待改进空间,比如上转换发光效率,纳米材料的生物安全性等方面。
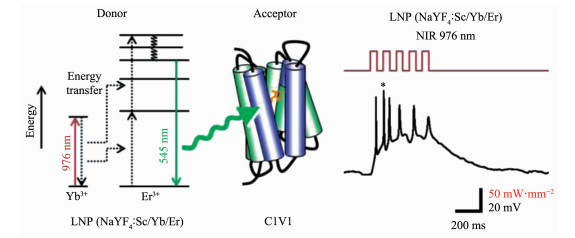
尽管上转换光遗传学系统具有灵活的适用性,但人们在实际应用中需要开发必要的刺激平台,以取得有效地光敏离子通道的近红外调控。目前报道了以下几种不同的设计:
(1) 将上转换纳米粒子嵌入细胞可生长的膜载体。如图 2a所示,Lee等[54]制备了上转换纳米材料(NaYF4:Yb/Tm@NaYF4)混合嵌入聚乳酸-羟基乙酸(PLGA)聚合物的薄膜。这种0.5 mm厚的薄膜不仅可用于神经元接触培养,还可以用作光遗传学调节的基础平台,将近红外激光转换为蓝光,随后激活表达有蓝色光敏通道视紫红质的蛋白(ChR2)的神经元。更关键地是,该体系通过1、5和10 Hz的980 nm脉冲激光刺激,实现了毫秒级分辨率的神经元活化响应。此外,Yawo等[40]比较分析了不同接触式细胞光遗传学调控平台的效率。根据图 2b所示,方法一将上转换纳米粒子与神经元直接接触,而方法二在二者之间采用玻片间隔。实验结果发现两种方法均以激光功率依赖性的方式显示出光刺激下向内性的膜电流响应。但是,在相同的近红外激发功率下,方法一的效率明显高于方法二,说明需要将上转换纳米材料尽可能靠近地放置于光敏通道蛋白的位置,因为光子的功率密度与距离的平方成反比。
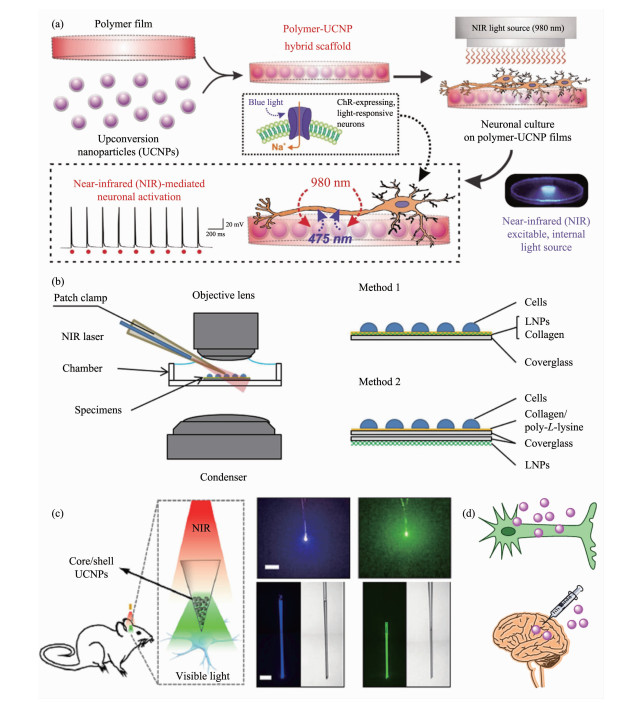
(2) 将上转换纳米粒子制成微光极。Shi等[57]首先将UCNPs包装到玻璃微光极中,制成可植入的光转换器将近红外能量转换为可见光,然后刺激具有不同ChRs表达的神经元(图 2c)。这些微型光学器件显示出极好的长期生物相容性,并且可以远程控制脑功能的调节,甚至用于复杂的动物行为学操控。
(3) 直接利用细胞或生物体组织摄取上转换纳米粒子(图 2d)。该方法在目前研究中最为常用,将功能修饰的上转换纳米粒子与所要调控的细胞孵育,或者直接通过注射的方式进入特定组织器官,待纳米粒子被有效摄入后,进行近红外激光照射并实现光遗传学调控。这种策略不仅可以达到亚细胞层面的精准光调控,而且在动物体内实验方面也易于实施。但是,局限也很突出,比如难以操作,生物安全性隐患等方面。
近红外上转换纳米技术和光遗传学的结合,从原理上克服了体内常用光遗传学研究中遇到的局限性,包括激发光的低穿透性或者光源植入的侵入性,为神经细胞或非神经体系中膜离子通道的调控提供了巨大的机会[32]。这种灵活的光学操控技术在神经科学领域取得了一系列成果,并进一步证明了其更广泛的适用性(表 3)[34, 69-70]。考虑到生物的内在复杂性,到目前为止,上转换光遗传学及其他调控技术,在详细阐明基本的生理、病理方面的探索仍然处于最初期阶段。因此,下面将着重讨论目前在不同生物模型上,采用近红外上转换光遗传学调节膜离子通道、钙信号以及相关生物学活性方面的研究进展。
下载:
导出CSV
| Optogenetic toolsa | UCNPs | λex / λem | Power densityb /(mW·mm-2) | Application | Ref. |
| LOV2-STIM1/ORAI1 | NaYF4:Yb/Tm@NaYF4 | 980 / 480 | 15~50 | Optogenetic activation Ca2+ signaling for immunomodulation in vitro and in vivo | [55] |
| ChR2 | NaYF4:Yb/Tm@NaYF4 | — | Optogenetic activation of Ca2+ channel in vitro | [54] | |
| NaYF4:Yb/Tm | — | Optogenetic activation of Ca2+ channel in vitro and in C. elegans | [58] | ||
| NaYbF4:Tm@NaYF4 | 50 | Optogenetic activation of ChR2 in vitro | [59] | ||
| NaYF4:Yb/Tm | 22.6 | [60] | |||
| NaYF4:Yb/Tm | 20 | Optogenetic stimulation of the spinal cord in behaving animals | [61] | ||
| NaYF4:Yb/Tm/Nd@NaYF4 | 808 / 480 | 8 | Optogenetic activation of Ca2+ channel in vitro and in zebrafish | [62] | |
| C1V1, mChR1, PsChR | NaYF4:Yb/Sc/Er or NaYF4:Yb/Sc/Tm@NaYF4 | 980 / 550 or 480 | 41 | Optogenetic activation of various ChRs in vitro | [40] |
| ChR2, C1V1 | NaYF4:Yb/Tm@NaYF4 NaYF4:Yb/Er@NaYF4 | 980 / 480 or 550 | 7 or 4 | Multiplexed optogenetic activation or inhibition of neurons in vitro and in vivo | [57, 63] |
| ReaChR | NaYF4:Yb/Er@ NaYF4@Dye | 800 / 550 | 20 | Optogenetic activation of hippocampal neurons in vitro | [64] |
| eNpHR | NaYF4@NaYF4:Yb/Er@NaYF4 | 980 / 550 | 6 | Optogenetic inhibition of neurons in vitro and in vivo | [65] |
| ChR2, Arch | NaYF4:Yb/Tm NaYF4:Yb/Er |
980 / 480 or 550 | 1 400 | Optogenetic deep brain activation and inhibition | [66] |
| Chrimson | NaGdF4:Yb/Er/Ca@ NaYbF4:Gd/Ca@NaNdF4:Gd | 808 / 550 | 640 | Optogenetic manipulation of the caenorhabditis elegans motor circuit | [67] |
| Jaw, VChR1 | NaErF4:Yb/Tm@ NaYF4:Yb@NaNdF4:Yb | 980 / 660 808 / 550 |
— | Programmable photoactivation of ion channels in vitro | [68] |
| a LOV2-STIM1/ORAI1, light-oxygen-voltage-sensing domain-engineered stromal interaction molecule-1/ORAI calcium release-activated calcium modulator 1; eNpHR, natronomonas halorhodopsin; b NIR laser power density used in practice, especially for in vivo stimulation | |||||
将光活性蛋白用于刺激活化或抑制神经活动是近年来神经学研究中的一大创新。这种光遗传手段由于具有较低的侵入性并且能够在时间与空间尺度精确地调节神经活动,因而在神经学研究中具有较为广阔的应用前景[23]。
早在2015年,Lee等[54]报道了通过UCNP进行神经调节的应用。该研究中视紫红质离子通道蛋白通过生物工程的手段表达于神经元细胞中,在近红外光(980 nm)的照射下,这些神经元能够在毫秒范围内产生持续的神经脉冲信号。在之后的研究中,科研人员设计并制备了一系列发光性质不同的UCNPs,用于不同光敏离子通道的活化[15, 31]。譬如,Han等[41]设计制备了IR806染料敏化的UCNP,通过800 nm的近红外光激发,实现了对红光响应离子通道(ReaChR)的调节。实验中,染料敏化的UCNP被包覆于聚甲基丙烯酸甲酯(PMMA)薄膜中,用于海马神经细胞的培养。这些神经细胞能够实现精确的时空调节,以光强度依赖性地方式被激活并产生神经信号。除此之外,在976 nm激发条件下能发射绿光(550 nm)的UCNPs(NaYF4:Sc/Yb/Er@NaYF4)也被用于刺激离子通道C1V1或mVChR1以产生光电流。而对于表达PsChR的神经元,则可以通过发射蓝光的UCNPs(NaYF4:Sc/Yb/Tm@NaYF4)实现近红外光遗传学刺激。上转换纳米材料可调的光学性质和不同光敏离子通道蛋白的灵活搭配为近红外光遗传学体系提供了丰富多样的选择,极大地促进了后续在动物模型上的生理、病理研究。
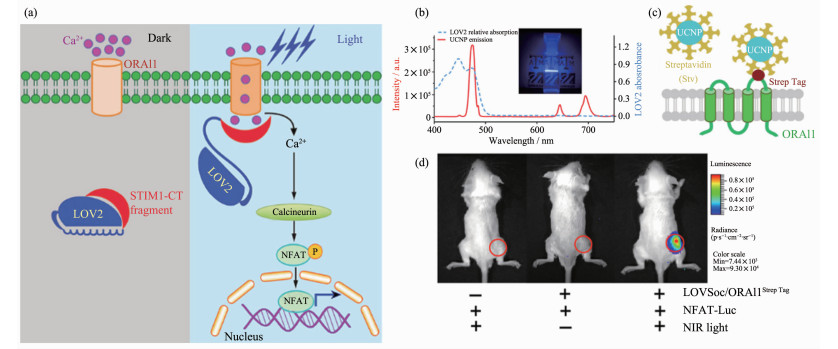
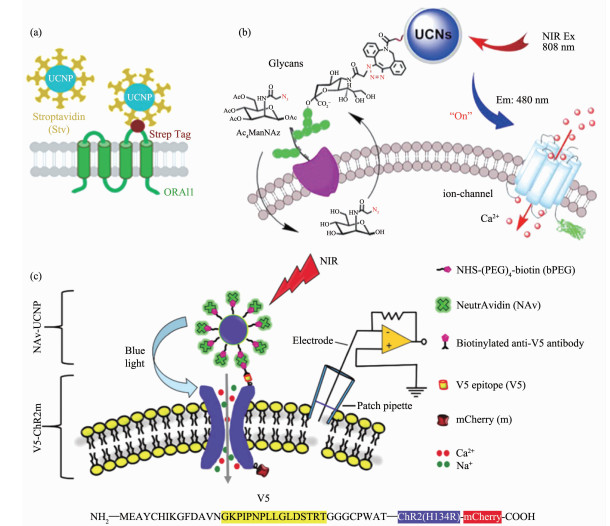
不同于常见报道的UCNP-ChR体系,Zhou和Han等[55]展示了另一种基于上转换纳米颗粒的近红外光遗传学平台,被称作“Opto-CRAC”(图 3a)。Opto-CRAC在细胞和活体环境中,通过近红外光照射而发出蓝光的UCNPs(NaYF4:Yb/Tm@NaYF4)作用于经基因工程改造的光敏钙离子通道蛋白,既可以选择性地控制细胞内钙离子的流入以及受此过程调控的基因表达,进而调节机体的免疫炎症反应(图 3(b~d))。通过对光信号的调节(如激光的脉冲、强度),该体系的光遗传模块LOVSoc能够可逆地产生持久且周期变化的钙离子信号。更为重要的是,Opto-CRAC介导的光致钙离子信号通路活化可以引发免疫细胞的特异性生理响应。通过使用近红外光激活光控-钙通道,可以促进树突状细胞的成熟及抗原的呈递,进而刺激T细胞的活化。通过这种手段实现了对细胞信号通路进行的精确操作,进而调控下游信号转导,方便其在动物生理/病理研究中发挥作用。
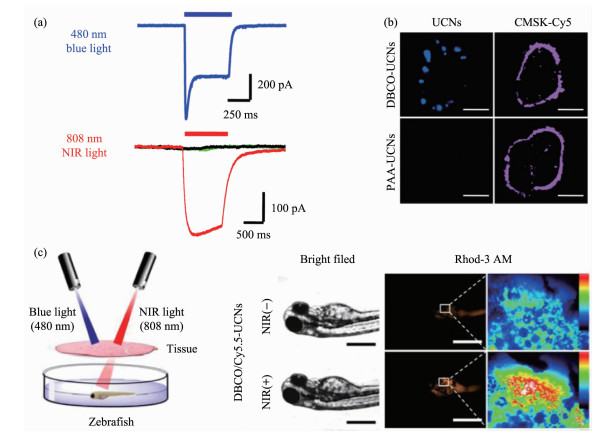
此外,斑马鱼活体模型广泛用于生命医药领域,比如生物造影,诊断治疗及生理病理研究[61]。目前,关于近红外光遗传学调控在斑马鱼模型中的可行性研究被Xing等[62]报道。该工作巧妙地设计了808 nm激发的上转换光遗传学体系,实现了对离子通道ChR2的调节以及钙离子介导的肿瘤细胞命运调控(图 4)。更为重要的是,该体系揭示了在体外和活体条件下,通过近红外光介导的离子通道的调节可引起细胞的凋亡,具有进一步在肿瘤治疗方面的应用前景。
除了能对细胞膜离子通道进行有效地调节外,在改进上转换光遗传学研究方面,Zhang等[58]通过利用准连续波的近红外激发手段,提高了上转换发光效率并在表达ChR2的线虫体内成功地实现光遗传学神经信号及行为学调控(图 5(a,b))。最近的一项研究中,Gao等[67]采用线虫模型,借助近红外激发绿光发射的UCNP对表达有Crimson光敏离子通道的不同神经元(运动神经元或中间神经元)进行调控,实现了对线虫多种运动行为的控制(图 5c)。这些工作不仅揭示了增强UCNP的多光子发光效率的可能性,还通过近红外光使线虫产生了受触动刺激的反应,展现出近红外光光遗传学这种非侵入性调控策略在不同活体动物模型应用上的优势和灵活性。

鼠类模型在生化、医药研究中作为一种最为普遍的动物模型,对于光遗传学的转化研究更具价值[30]。尽管深层组织的光学刺激在鼠类模型中很大程度上需要特殊的光学设施,以及复杂的实验手术操作,但随着光遗传学和上转换纳米技术的发展,通过近红外光进行小鼠脑部功能的光遗传学调控已经成为可能。
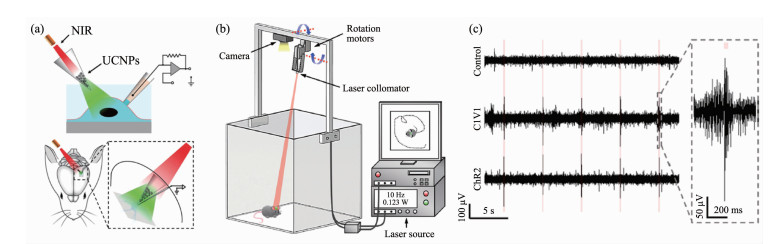
Shi等[63]报道了一种基于UCNP的微型光极器件实现了远程光学控制小鼠脑部神经元(表达有多种视蛋白如ChR2或C1V1)的目标(图 6)。机械激光投射系统通过发出的近红外光,可以有效地控制鼠的不同脑部区域的功能并产生神经脉冲活动,譬如脑部纹状体,中脑腹侧盖区以及视觉皮层。值得注意的是,在另一项工作中,Shi等[63]还通过设计制备不同光谱特征的UCNPs,实现了体外与体内条件下,神经细胞的多重刺激或抑制。上转换光遗传技术在小鼠神经活动刺激研究中的成功应用,极大地促进了基础生理调控以及神经科学的发展。
另外,McHugh及Liu[66]探究了将UCNPs作为光遗传调节器以微侵入注射式的方法调控小鼠脑部深层神经元功能的可能性(图 7)。该研究中,近红外介导的光遗学传刺激能够引发脑部腹侧被盖区域多巴胺的释放。在此基础上,通过刺激脑部内侧隔核的抑制性神经元,可以诱导产生神经振荡。更为重要的是,研究中这种上转换光遗传体系还可以抑制癫痫小鼠海马体中的兴奋性神经元,实现记忆恢复,展现了其在神经疾病治疗方面的巨大潜力。

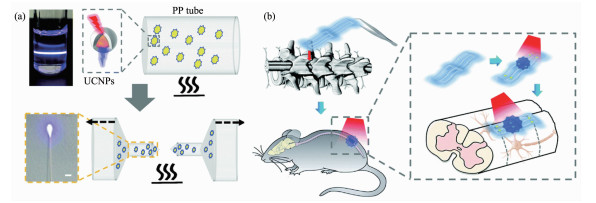
除了脑部神经活动调控,上转换光遗传体系还被成功地应用于小鼠的脊柱神经调节并用以控制小鼠的行为活动。Shi等[61]用聚丙烯以及UCNP制成光极器件,植入小鼠脊柱不同部位中(图 8)。这些小鼠在已麻醉的情况下,通过近红外光的刺激,可以由肌电图观测到其腿部肌肉的活动。而自由活动的小鼠,在光的刺激下其运动行为还能被有效地抑制。这种柔性器件与上转换光遗传学的结合,还展现出较好的生物相容性。在长达4个月的植入期内未引起明显的炎症,适用于动物行为学的长期跟踪研究。
尽管上转换光遗传学前景广阔,但目前仍然面临着纳米粒子生物安全性低,近红外光上转换效率低,以及持续照射引起的热效应显著等挑战[32, 71]。为了解决这些问题,科学家们已经从各方面入手来改进这项技术,以期实现更高效,更精确地生物功能调控,并进一步使未来的临床转化研究成为可能。
首先,上转换纳米材料的量子效率低是实现有效地光遗传学调控的最大限制因素。到目前为止,在低于100 W·cm-2的激发光功率密度下,近红外到可见光上转换效率最高仅约为5%,而考虑到实际应用中,生物机体对于辐射暴露的最大允许剂量(例如980 nm,皮肤组织MPE(maximum permissible exposure) < 1 W·cm-2)的限制,通常上转换效率会远远低于1%[72]。对此,许多研究团队提出了增强上转换效率的不同策略,包括合成核-壳结构或表面修饰来控制局部环境的猝灭效应,利用敏化剂/活化剂来改善能量转移效率,以及通过激发光源的工程化来促进光子转移等[73-74]。
将增强上转换发光效率的策略进一步应用于光遗传学调控的研究已有报道。Shi和Wang研究团队[65]最近通过合成核-壳-壳纳米结构,并优化Yb3+离子的掺杂含量,实现了3倍于传统核-壳结构纳米粒子的上转换发光增强,并将其进一步开发为一种可植入的光学传感器,用于对表达eNpHR氯离子通道的小鼠大脑活性及行为学进行光遗传学抑制。另外,Prasad等[59]在一项近红外光遗传学的工作中应用了新型的核-壳型上转换纳米粒子(NaYbF4:Tm@NaYF4),这种材料的发光强度比传统NaYF4:Yb/Tm@NaYF4纳米体系发光约高出6倍。此外,Zhang团队[58]通过使用准连续波作为激发光源来进行光遗传学实验,不仅提高了上转换蓝光发射,还降低了潜在的热效应,从而实现了有效的光遗传神经调控。
利用近红外上转换技术可以有效地激活深层组织中的光敏膜离子通道。但应该指出的是,大部分上转换纳米粒子在980 nm激光照射下可能会导致局部组织的热损伤[72]。为了避免这种热效应,在实际上转换光遗传学调控中,大部分的研究只能通过合理控制激光的功率以及照射时间来控制。除此以外,将上转换激发波长从980 nm移至800 nm,在很大程度上降低了机体组织水的热响应,可以极大地减少激光引起的热刺激[75]。例如,在Han等[64]的报道中,使用染料敏化的核/壳结构上转换材料,可以在800 nm近红外光照射下激活海马神经元中的离子通道蛋白(ReaChR)。
在生物医学乃至临床应用中,无机金属纳米材料在体内的生物相容性或潜在毒性是一个重要的问题[76]。迄今为止,尚无详尽的报道涉及上转换纳米粒子本身或用于表面功能化的相关试剂和配体的刺激性及长期毒性的研究,如免疫反应和诱变作用。但可以证实的是,上转换纳米材料的形貌尺寸、化学组成及表面修饰都影响其在体外和体内的安全性[76-80]。有一些常规的策略可以在一定程度上降低上转换纳米体系的毒性,比如制备超小尺寸纳米粒子(< 10 nm)以增强生物清除率[81];选择合适的配体进行表面修饰,例如聚乙二醇(PEG)、二氧化硅(SiO2)等安全性高的生物功能修饰剂;再者,通过增强上转换发光进而降低纳米材料的使用浓度,也是一种解决剂量依赖的毒性问题行之有效的方法[82-83]。另一方面,由于生物体本身没有光遗传学工具,而通过病毒或聚合物的基因转染手段在体内表达光敏蛋白也具有一定的安全性顾虑。
尽管通过上转换光遗传学在神经元或非神经元调节方面取得了初步成功,但是目前在实际应用中光控生理功能的效率还受限于调控的精确性。主要表现在以下几个方面:
(1) 光遗传学工具的离子选择性。目前广泛使用的光敏感视紫红质蛋白,尤其是阳离子通道蛋白(例如ChRs),其激活后对Ca2+、Na+或K+等的细胞内流缺乏选择性,因此难以做到精准控制生理信号。对于如何解决离子选择性问题,有以下两种可能的策略:一是对已有的ChRs进行基因工程改造,得到具有高离子选择性的突变体,但至今还鲜有相关报道;二是结合其他现有的光遗传学技术,构建离子选择性好的光遗传学工具。例如,基于特异性的Ca2+通道激活释放的光遗传学平台(Opto-CRAC),在体外和体内都体现出优秀的Ca2+信号调节能力[55, 84];另外,近年来报道的热敏离子通道(TRPs)也极具潜力,有望实现高选择性的Ca2+信号调控。
(2) 光遗传学调控的细胞/组织特异性。许多在细胞层面上的光遗传学研究表明,上转换纳米转换器尽可能地靠近膜离子通道蛋白,可显著提高光子能量转移的效率,对调控效果产生重要影响。目前已报道了不同的策略,被用于将上转换纳米粒子尽可能特异地连接在光敏蛋白表达的细胞膜上(图 9)。其中包括抗原-抗体结合的策略,将UCNPs定位于细胞表面,还可以通过糖代谢标记技术来共价连接。
而在动物层面,目前通过使用靶向的光基因递送技术,如细胞/组织特异性慢病毒感染等可以实现特异性的光遗传学调控[85-87]。但在实际的神经科学领域应用中,需要借助于脑定位注射来实现精确光遗传学基因及上转换纳米体系在目的组织区域表达。由于大脑结构的复杂性,很难保证操作的精准度,这在光遗传学研究中也是一个很大的挑战。
(3) 纳米粒子靶向性。除此以外,上转换纳米粒子的靶向能力是远程调控细胞/组织特异性离子通道的另一个限制因素[88-89]。例如,在深层脑区的光遗传学操作中,纳米粒子难以有效通过血脑屏障(BBB),因而只能通过注射的方法来输送上转换纳米颗粒。这种策略不仅增加了创伤性,还有可能引入一些潜在的不良反应。因此,开发完全无创的、可血液递送的上转换纳米平台,进而能够用于脑部的近红外光遗传学体系将是未来研究的一大方向。
最后,除了对上转换纳米转换器和光遗传学工具的改进之外,可靠的光学调控仪器以及信号记录设备在实际的上转换光遗传学应用也亟待开发。迄今为止,用于上转换材料光学表征的大多数仪器都是实验室个人定制。此外,在动物模型中用于近红外激光介导的光遗传刺激、监测的专用仪器也非常有限[90-91]。因此,目前的上转换光遗传学研究的稳定性和重复性还不尽人意,亟待研发商用的、标准化的仪器(例如光谱仪,刺激器,显微成像系统及膜片钳设备等)。总体而言,由于近红外上转换光遗传学是一个多学科、高度交叉的领域,学术界和工业界的任何建设性合作与整合都将有利于该技术的转化应用。
总之,通过将上转换纳米材料作为光转换器,与光遗传学体系结合起来,新兴的上转换光遗传学平台可实现对膜离子通道的近红外操控。并且该技术具有更深的组织穿透力,更高的时空分辨率和低的创伤性,克服了传统光遗传学在活体研究中遇到的问题。尽管上转换光遗传学具有广阔的前景,并已被广泛应用于神经活性、行为学调控,甚至疾病的治疗。但目前在实践中依然存在着局限,比如上转换效率低、热效应明显和特异性不足等。鉴于此,研究者们也做出了巨大的努力来不断地克服这些问题。我们相信,这项强大的化学生物学技术将加深我们对离子通道的生理和相关病理方面的理解,并为神经系统疾病及其他威胁生命的重大疾病的治疗开辟新的方向。
Ackerman M J, Clapham D E. New Engl. J. Med., 1997, 336: 1575-1586 doi: 10.1056/NEJM199705293362207
Dunlop J, Bowlby M, Peri R, et al. Nat. Rev. Drug Discov., 2008, 7:358 doi: 10.1038/nrd2552
Loewenstein W R. Physiol. Rev., 1981, 61:829-913 doi: 10.1152/physrev.1981.61.4.829
Hamill O P, McBride D. Pharmacol. Rev., 1996, 48:231-252 https://www.researchgate.net/publication/14400829_The_pharmacology_of_mechanogated_membrane_ion_channels_Review
Alexander S P, Mathie A, Peters J A. Br. J. Pharmacol., 2011, 164:S137-S174 doi: 10.1111/j.1476-5381.2011.01649_5.x
Reuter H. Nature, 1983, 301:569-574 doi: 10.1038/301569a0
Miller C. Neuron, 1989, 2:1195-1205 doi: 10.1016/0896-6273(89)90304-8
Armstrong C M, Hille B. Neuron, 1998, 20:371-380 doi: 10.1016/S0896-6273(00)80981-2
Nicolelis M A. Nat. Rev. Neurosci., 2003, 4:417-422 doi: 10.1038/nrn1105
Zamponi G W. Nat. Rev. Drug Discov., 2016, 15:19-34 doi: 10.1038/nrd.2015.5
Erwin B, Jr M M, Baker K K. Neurol. Res., 2000, 22:259-266 doi: 10.1080/01616412.2000.11740668
Kringelbach M L, Jenkinson N, Owen S L, et al. Nat. Rev. Neurosci., 2007, 8:623-635 doi: 10.1038/nrn2196
Gautier A, Gauron C, Volovitch M, et al. Nat. Chem. Biol., 2014, 10:533-541 doi: 10.1038/nchembio.1534
Ankenbruck N, Courtney T, Naro Y, et al. Angew. Chem. Int. Ed., 2018, 57:2768-2798 doi: 10.1002/anie.201700171
Grier D G. Nature, 2003, 424:810 doi: 10.1038/nature01935
Fenno L, Yizhar O, Deisseroth K. Annu. Rev. Neurosci., 2011, 34:389-412 doi: 10.1146/annurev-neuro-061010-113817
Yizhar O, Fenno L E, Davidson T J, et al. Neuron, 2011, 71: 9-34 doi: 10.1016/j.neuron.2011.06.004
Repina N A, Rosenbloom A, Mukherjee A, et al. Annu. Rev. Chem. Biomol. Eng., 2017, 8:13-39 doi: 10.1146/annurev-chembioeng-060816-101254
Rein M L, Deussing J M. Mol. Genet. Genomics, 2012, 287: 95-109 doi: 10.1007/s00438-011-0663-7
Zhang F, Vierock J, Yizhar O, et al. Cell, 2011, 147:1446- 1457 doi: 10.1016/j.cell.2011.12.004
Deisseroth K. Nat. Neurosci., 2015, 18:1213-1225 doi: 10.1038/nn.4091
Han X. ACS Chem. Neurosci., 2012, 3:577-584 doi: 10.1021/cn300065j
Husser M. Nat. Methods, 2014, 11:1012-1014 doi: 10.1038/nmeth.3111
Knpfel T, Lin M Z, Levskaya A, et al. J. Neurosci., 2010, 30: 14998-15004 doi: 10.1523/JNEUROSCI.4190-10.2010
Lin J Y. Exp. Physiol., 2011, 96:19-25 doi: 10.1113/expphysiol.2009.051961
Kale R P, Kouzani A Z, Walder K, et al. Neurophotonics, 2015, 2:031206 doi: 10.1117/1.NPh.2.3.031206
Rossi M A, Go V, Murphy T, et al. Front. Integr. Neurosci., 2015, 9:8
Piatkevich K D, Subach F V, Verkhusha V V. Chem. Soc. Rev., 2013, 42:3441-3452 doi: 10.1039/c3cs35458j
Redchuk T A, Omelina E S, Chernov K G, et al. Nat. Chem. Biol., 2017, 13:633-639 doi: 10.1038/nchembio.2343
Urmann D, Lorenz C, Linker S M, et al. Photochem. Photobiol., 2017, 93:782-795 doi: 10.1111/php.12741
Zhang F, Prigge M, Beyrière F, et al. Nat. Neurosci., 2008, 11: 631-633 doi: 10.1038/nn.2120
Huang K, Dou Q, Loh X J. RSC Adv., 2016, 6:60896-60906 doi: 10.1039/C6RA11289G
Shah S, Solanki A, Lee K B. Acc. Chem. Res., 2015, 49:17- 26
Scaini D, Ballerini L. Curr. Opin. Neurobiol., 2018, 50:50-55
Wang F, Liu X G. Chem. Soc. Rev., 2009, 38:976-989 doi: 10.1039/b809132n
Chen G Y, Qiu H L, Prasad P N, et al. Chem. Rev., 2014, 114:5161-5214 doi: 10.1021/cr400425h
Idris N M, Jayakumar M K G, Bansal A, et al. Chem. Soc. Rev., 2015, 44:1449-1478 doi: 10.1039/C4CS00158C
Wen S H, Zhou J J, Zheng K Z, et al. Nat. Commun., 2018, 9:2415 doi: 10.1038/s41467-018-04813-5
Yang Y M, Shao Q, Deng R R, et al. Angew. Chem. Int. Ed., 2012, 124:3179-3183 doi: 10.1002/ange.201107919
Hososhima S, Yuasa H, Ishizuka T, et al. Sci. Rep., 2015, 5: 16533 doi: 10.1038/srep16533
Wang Z M, Hu M, Ai X Z, et al. Adv. Biosyst., 2019, 3: 1800233 doi: 10.1002/adbi.201800233
Boyden E S, Zhang F, Bamberg E, et al. Nat. Neurosci., 2005, 8:1263-1268 doi: 10.1038/nn1525
Lin J Y, Lin M Z, Steinbach P, et al. Biophys. J., 2009, 96: 1803-1814 doi: 10.1016/j.bpj.2008.11.034
Gunaydin L A, Yizhar O, Berndt A, et al. Nat. Neurosci., 2010, 13:387-392 doi: 10.1038/nn.2495
Berndt A, Yizhar O, Gunaydin L A, et al. Nat. Neurosci., 2009, 12:229-234 doi: 10.1038/nn.2247
Klapoetke N C, Murata Y, Kim S S, et al. Nat. Methods, 2014, 11:338-346 doi: 10.1038/nmeth.2836
Yizhar O, Fenno L, Prigge M, et al. Nature, 2011, 477:171- 178 doi: 10.1038/nature10360
Zhang F, Wang L P, Brauner M, et al. Nature, 2007, 446: 633-639 doi: 10.1038/nature05744
Wietek J, Wiegert J S, Adeishvili N, et al. Science, 2014, 344:409-412 doi: 10.1126/science.1249375
Govorunova E G, Sineshchekov O A, Janz R, et al. Science, 2015, 349:647-650 doi: 10.1126/science.aaa7484
Chow B Y, Han X, Dobry A S, et al. Nature, 2010, 463:98- 102 doi: 10.1038/nature08652
Towne C, Thompson K R. Curr. Protoc. Pharmacol., 2016, 75: 11-19
Deisseroth K, Anikeeva P. US Patent, 9522288. 2016-12-20.
Shah S, Liu J J, Pasquale N, et al. Nanoscale, 2015, 7:16571 -16577 doi: 10.1039/C5NR03411F
He L, Zhang Y W, Ma G L, et al. Elife, 2015, 4:e10024 doi: 10.7554/eLife.10024
Hososhima S, Yuasa H, Ishizuka T, et al. International Society for Optics and Photonics: Optical Techniques in Neurosurgery, Neurophotonics, and Optogenetics Ⅱ, 2015:93052R
Wang Y, Lin X D, Chen X, et al. Biomaterials, 2017, 142: 136-148 doi: 10.1016/j.biomaterials.2017.07.017
Bansal A, Liu H C, Jayakumar M K G, et al. Small, 2016, 12:1732-1743 doi: 10.1002/smll.201503792
Pliss A, Ohulchanskyy T Y, Chen G Y, et al. ACS Photonics, 2017, 4:806-814 doi: 10.1021/acsphotonics.6b00475
Yadav K, Chou A C, Ulaganathan R K, et al. Nanoscale, 2017, 9:9457-9466 doi: 10.1039/C7NR03246C
Wang Y, Xie K, Yue H B, et al. Nanoscale, 2020, 12:2406- 2414 doi: 10.1039/C9NR07583F
Ai X Z, Lyu L N, Zhang Y, et al. Angew. Chem. Int. Ed., 2017, 56:3031-3035 doi: 10.1002/anie.201612142
Lin X D, Wang Y, Chen X, et al. Adv. Healthcare Mater., 2017, 6:1700446 doi: 10.1002/adhm.201700446
Wu X, Zhang Y W, Takle K, et al. ACS Nano, 2016, 10:1060- 1066 doi: 10.1021/acsnano.5b06383
Lin X D, Chen X, Zhang W C, et al. Nano Lett., 2018, 18: 948-956 doi: 10.1021/acs.nanolett.7b04339
Chen S, Weitemier A Z, Zeng X, et al. Science, 2018, 359: 679-684 doi: 10.1126/science.aaq1144
Ao Y X, Zeng K H, Yu B, et al. ACS Nano, 2019, 13:3373- 3386 doi: 10.1021/acsnano.8b09270
Mei Q, Bansal A, Jayakumar M K G, et al. Nat. Commun., 2019, 10:1-11 doi: 10.1038/s41467-018-07882-8
Zhang Y W, Huang L, Li Z J, et al. ACS Nano, 2016, 10: 3881-3885 doi: 10.1021/acsnano.6b02284
Tan P, He L, Han G, et al. Trends Biotechnol., 2017, 35:215 -226 doi: 10.1016/j.tibtech.2016.09.002
Vogt N. Nat. Methods, 2018, 15:242
Wilhelm S. ACS Nano, 2017, 11:10644-10653 doi: 10.1021/acsnano.7b07120
Han S Y, Deng R R, Xie X J, et al. Angew. Chem. Int. Ed., 2014, 53:11702-11715 doi: 10.1002/anie.201403408
Saboktakin M, Ye X, Chettiar U K, et al. ACS Nano, 2013, 7:7186-7192 doi: 10.1021/nn402598e
Wang Y F, Liu G Y, Sun L D, et al. ACS Nano, 2013, 7: 7200-7206 doi: 10.1021/nn402601d
Buzea C, Pacheco I I, Robbie K. Biointerphases, 2007, 2: MR17-MR71 doi: 10.1116/1.2815690
Sun Y, Feng W, Yang P Y, et al. Chem. Soc. Rev., 2015, 44: 1509-1525 doi: 10.1039/C4CS00175C
Gnach A, Lipinski T, Bednarkiewicz A, et al. Chem. Soc. Rev., 2015, 44:1561-1584 doi: 10.1039/C4CS00177J
Vecchio G, Galeone A, Brunetti V, et al. Nanomed. Nanotechnol. Biol. Med., 2012, 8:1-7 doi: 10.1016/j.nano.2011.11.001
Hoet P H, Brüske-Hohlfeld I, Salata O V. J. Nanobiotechnol., 2004, 2:12 doi: 10.1186/1477-3155-2-12
Zarschler K, Rocks L, Licciardello N, et al. Nanomed. Nanotechnol. Biol. Med., 2016, 12:1663-1701 doi: 10.1016/j.nano.2016.02.019
Zhan Q Q, Qian J, Liang H J, et al. ACS Nano, 2011, 5: 3744-3757 doi: 10.1021/nn200110j
McNally K M, Sorg B S, Welch A J, et al. Phys. Med. Biol., 1999, 44:983-1002 doi: 10.1088/0031-9155/44/4/013
Ma G L, Liu J D, Ke Y P, et al. Angew. Chem. Int. Ed., 2018, 57:7019-7022 doi: 10.1002/anie.201713080
Thomas C E, Ehrhardt A, Kay M A. Nat. Rev. Genet., 2003, 4:346-358 doi: 10.1038/nrg1066
Cardin J A, Carlén M, Meletis K, et al. Nat. Protoc., 2010, 5: 247-254 doi: 10.1038/nprot.2009.228
Zhao S, Ting J T, Atallah H E, et al. Nat. Methods, 2011, 8: 745-752 doi: 10.1038/nmeth.1668
Huang J G, Leshuk T, Gu F X. Nano Today, 2011, 6:478- 492 doi: 10.1016/j.nantod.2011.08.002
Bernstein J G, Garrity P A, Boyden E S. Curr. Opin. Neurobiol., 2012, 22:61-71 doi: 10.1016/j.conb.2011.10.023
Klimas A, Entcheva E G. J. Biomed. Opt., 2014, 19:080701 doi: 10.1117/1.JBO.19.8.080701
Papagiakoumou E. Biol. Cell, 2013, 105:443-464


图 2 不同上转换光遗传学刺激方法: (a)上转换纳米粒子-聚合物杂合的膜平台[54]; (b) 2种上转换纳米粒子调控体系[40]; (c)基于上转换纳米粒子的微型光极器件; (d)细胞或组织摄取上转换纳米粒子的示意图[57]
Figure 2 Different optogenetic stimulation platforms: (a) UCNPs-polymers hybrid membrane for cells culture and optogenetic activation[54]; (b) Two membrane-based UCNPs systems for optogenetic regulation[40]; (c) UCNPs-based optotrode micro-devices; (d) Schematic illustration of UCNPs uptake by cells and tissues[57]

图 3 Opto-CRAC (CRAC, Ca2+ release-activated Ca2+)体系用于细胞膜钙离子通道的光遗传调控[55]: (a) LOVSoc介导的细胞内光致钙离子内流以及NFAT(nuclear factor of activated T-cells)的核转移; (b) UCNPs (NaYF4:Yb/Tm@NaYF4)的发射光谱; (c)亲和素修饰的UCNPs与细胞膜表面的ORAI1钙离子通道相互作用示意图; (d)通过近红外光介导的活体内细胞钙离子内流, 进而引发NFAT依赖的萤光素酶表达
Figure 3 Illustration of Opto-CRAC (CRAC, Ca2+ release-activated Ca2+) system for optogenetic regulation of membrane Ca2+ channel[55]: (a) LOVSoc photoactivation mediated Ca2+ influx and NFAT (nuclear factor of activated T-cells) nuclear translocation; (b) UCNPs (NaYF4:Yb/Tm@NaYF4) luminescence spectra; (c) Specific targeting of streptavidin- conjugated UCNPs to engineered ORAI1 channels in the plasma membrane; (d) NFAT-dependent luciferase expression in vivo triggered by NIR light stimulation

图 4 斑马鱼模型中通过细胞膜靶向的上转换光遗传学技术进而实现离子通道的精确调控[62]: (a)细胞表达ChR2以及在可见光及近红外光照射下产生的电生理; (b)通过糖代谢途径标记于细胞膜表面的UCNPs成像; (c)近红外光触发的体内ChR2激活以及钙离子内流(CMSK-Cy5, 细胞膜特异性荧光染料)
Figure 4 Membrane targeted upconversion optogenetic technique for remote regulation of ion channels in the zebrafish model[62]: (a) Photocurrents in ChR2-expressing cells labeled with UCNPs under NIR and blue light excitation; (b) Imaging of plasma membrane localized UCNPs through glycan metabolic labeling strategy; (c) NIR light excited in vivo ChR2 activation and Ca2+ influx (CMSK-Cy5, cell membrane staining fluorescence dye)

图 5 上转换近红外光遗传平台实现对线虫行为学调控[58, 67]:机械感受神经元中表达ChR2的线虫, 在标记UCNP后, (a)于近红外光刺激下产生的不同行为动作及(b)在近红外光遗传刺激下表现的转向动作; (c)通过对Chrimson表达的神经元进行近红外光刺激, 实现线虫不同运动形态的调控
Figure 5 Upconversion optogenetic manipulation of C. elegans behaviors by NIR excitation[58, 67]: (a) Different movement behavior in worms expressing ChR2 in their mechanosensory neurons with NIR light illumination alone (no UCNPs), and (b) NIR light illumination in the presence of UCNPs; (c) Schematic diagram of UCNPs for chrimson-expressing cells activation illuminated by 808 nm NIR light, thereby changing the locomotion behaviors

图 6 通过近红外上转换光遗传手段实现的小鼠脑部活动[63]: (a)通过近红外光光遗传学平台在体外和活体条件下对表达ChR的神经元刺激活化; (b)用于对活动小鼠头部提供近红外光刺激的自动化激光投射系统; (c)由近红外刺激引起的小鼠脑部(表达ChR2和C1V1)神经脉冲信号
Figure 6 NIR upconversion optogenetic control of rat brain activity[63]: (a) NIR stimulation of ChRs-expressing neurons using UCNPs in vitro and in vivo; (b) Robotic laser projection and monitoring systems for automatic and consistent NIR irradiation of the heads of behaving animals; (c) Recordings of spiking trances in neurons in response to NIR illuminations

图 7 UCNP介导的脑部深层的近红外上转换光遗传学刺激[66]: (a)发射蓝光的UCNP介导的近红外上转换光遗传在TH-Cre转基因小鼠表达ChR2的神经元作用原理示意图; (b)脑部深层组织的AAV-DIO-ChR2-EYFP、UCNP的注射以及近红外光遗传刺激示意图; (c)通过颅部近红外照射抑制癫痫小鼠脑部海马区活动; (d)颅脑近红外光照射刺激内侧隔核以诱导产生神经振荡; (e)刺激海马印记细胞实现记忆恢复
Figure 7 UCNPs-mediated NIR optogenetics for deep brain stimulation[66]: (a) Illustration of blue emissive UCNPs for NIR optogenetic activation of ChR2-expressing neurons TH-Cre transgenic mice; (b) Scheme of AAV-DIO-ChR2-EYFP and UCNPs injection and NIR optogenetic stimulation in deep brain tissue; (c) Illustration of transcranial NIR inhibition of hippocampal (HIP) activity during chemically induced seizure, (d) medial septum (MS) for generation of θ oscillations and (e) hippocampal engram formemory recall

图 8 上转换光极器件用于光遗传学脊髓调控[61]: (a)含UCNPs的聚丙烯材料融化拉制成纤维状的上转换光极; (b)通过微创手术将上转换微光极(UCNP-PP-optrode)植入小鼠的脊柱并进行光遗传学调控
Figure 8 Implantable upconversion device for wireless optogenetic stimulation of the spinal cord in behaving animals[61]: (a) Diagram of the UCNPs-optrode fabrication; (b) Illustration of implantation of the UCNP-PP-optrode at the spinal cord of a mouse by minimal invasive surgery

图 9 几种细胞膜连接上转换纳米粒子用于精准光遗传学调控的策略: (a)基于亲和素-生物素系统将上转换纳米材料与光敏蛋白ORAI1连接[55]; (b, c)基于糖代谢以及亲和素-生物素的方法将上转换材料在细胞膜标记用于ChR2活化[60, 62]
Figure 9 Different plasma membrane labeling strategies for precise upconversion optogenetic manipulation: (a) Biotin-streptavidin interaction mediated UCNPs-ORAI1 labeling strategy[55]; (b, c) Glycan metabolic and biotin-streptavidin labeling strategies[60, 62]
表 1 常用的视紫红质离子通道蛋白(ChRs)
Table 1. Commonly used channelrhodopsins (ChRs)
| ChRsa | λmax (activation) / nm | Function and origin | Ref. |
| ChR2 | 480 | Excitatory cation channel; Native from C. reinhardtii | [42] |
| VChR1 | 545 | Red-shifted, excitatory cation channel; Native from V. carteri | [31] |
| ChIEF | 450 | Fast kinetics, excitatory cation channel; Chimera of ChR1 and VChR1 | [43] |
| ChETA | 470 | Fast kinetics, excitatory cation channel; Substitutions in ChR2 | [44] |
| Step function opsins | 470 (on) 590 (off) | Excitatory cation channel; Single mutations in ChR2 | [45] |
| Chronos | 500 | Fast kinetics, excitatory cation channel; Native from Stigeoclonium helveticum | [46] |
| Chrimson | 590 | Red-shifted, excitatory cation channel; Native from Chlamydomonas noctigama | |
| C1V1 | 540 | Red-shifted, excitatory cation channel; Chimera of ChR1 and VChR1 | [47] |
| NpHR | 590 | Inhibitory chloride pump; Native from N. pharaonis | [48] |
| ChloC | 480 | Inhibitory anion channel; Mutagenesis of ChR2 | [49] |
| GtACR2 | 520 | Inhibitory anion pump; Native from G. theta | [50] |
| ArCh | 570 | Inhibitory proton pump; Native from H. sodomense | [51] |
| a ChR2, channelrhodopsin-2; ReaChR, red-shifted channelrhodopsin; NpHR, natronomonas halorhodopsin; ChloC and GtACR2, chloride-conducting channelrhodopsin; Arch, archaerhodopsin; other mutations of channelrhodopsins (VChR1, C1V1, ChIEF, ChETA, mChR1, PsChR)[52] | |||
 下载: 导出CSV
下载: 导出CSV
表 2 常用的上转换纳米粒子(UCNPs)
Table 2. Commonly used upconversion nanoparticles (UCNPs)
| UCNPs | Ln3+ | λex / nm | λem / nm |
| NaYF4:Yb/Ln3+ or NaGdF4:Yb/Ln3+ | Tm3+ | 980 | 450, 475, 800 |
| Er3+ | 545, 665 | ||
| Pr3+ | 480, 606 | ||
| Tb3+ | 480~650 | ||
| Eu3+ | 570~720 | ||
| Sm3+ | 601 | ||
| Ho3+ | 540 | ||
| Dy3+ | 486, 575 | ||
| NaYF4:Yb/Nd3+/Ln3+ or NaYF4:Yb/Nd3+/Ln3+/Dye | Ln3+ | 980 or 808 | As above |
下载: 导出CSV
表 3 当前上转换光遗传学的应用总结
Table 3. Summary of current applications of upconversion optogenetics
| Optogenetic toolsa | UCNPs | λex / λem | Power densityb /(mW·mm-2) | Application | Ref. |
| LOV2-STIM1/ORAI1 | NaYF4:Yb/Tm@NaYF4 | 980 / 480 | 15~50 | Optogenetic activation Ca2+ signaling for immunomodulation in vitro and in vivo | [55] |
| ChR2 | NaYF4:Yb/Tm@NaYF4 | — | Optogenetic activation of Ca2+ channel in vitro | [54] | |
| NaYF4:Yb/Tm | — | Optogenetic activation of Ca2+ channel in vitro and in C. elegans | [58] | ||
| NaYbF4:Tm@NaYF4 | 50 | Optogenetic activation of ChR2 in vitro | [59] | ||
| NaYF4:Yb/Tm | 22.6 | [60] | |||
| NaYF4:Yb/Tm | 20 | Optogenetic stimulation of the spinal cord in behaving animals | [61] | ||
| NaYF4:Yb/Tm/Nd@NaYF4 | 808 / 480 | 8 | Optogenetic activation of Ca2+ channel in vitro and in zebrafish | [62] | |
| C1V1, mChR1, PsChR | NaYF4:Yb/Sc/Er or NaYF4:Yb/Sc/Tm@NaYF4 | 980 / 550 or 480 | 41 | Optogenetic activation of various ChRs in vitro | [40] |
| ChR2, C1V1 | NaYF4:Yb/Tm@NaYF4 NaYF4:Yb/Er@NaYF4 | 980 / 480 or 550 | 7 or 4 | Multiplexed optogenetic activation or inhibition of neurons in vitro and in vivo | [57, 63] |
| ReaChR | NaYF4:Yb/Er@ NaYF4@Dye | 800 / 550 | 20 | Optogenetic activation of hippocampal neurons in vitro | [64] |
| eNpHR | NaYF4@NaYF4:Yb/Er@NaYF4 | 980 / 550 | 6 | Optogenetic inhibition of neurons in vitro and in vivo | [65] |
| ChR2, Arch | NaYF4:Yb/Tm NaYF4:Yb/Er |
980 / 480 or 550 | 1 400 | Optogenetic deep brain activation and inhibition | [66] |
| Chrimson | NaGdF4:Yb/Er/Ca@ NaYbF4:Gd/Ca@NaNdF4:Gd | 808 / 550 | 640 | Optogenetic manipulation of the caenorhabditis elegans motor circuit | [67] |
| Jaw, VChR1 | NaErF4:Yb/Tm@ NaYF4:Yb@NaNdF4:Yb | 980 / 660 808 / 550 |
— | Programmable photoactivation of ion channels in vitro | [68] |
| a LOV2-STIM1/ORAI1, light-oxygen-voltage-sensing domain-engineered stromal interaction molecule-1/ORAI calcium release-activated calcium modulator 1; eNpHR, natronomonas halorhodopsin; b NIR laser power density used in practice, especially for in vivo stimulation | |||||
下载: 导出CSV

 扫一扫看文章
扫一扫看文章

扫一扫关注我们
 下载:
下载: