School of Traditional Chinese Materia Medica, Shenyang Pharmaceutical University, Shenyang 110016, China
b.
State Key Laboratory of Bioactive Substance and Function of Natural Medicines, Institute of Materia Medica, Chinese Academy of Medical Sciences & Peking Union Medical College, Beijing 100050, China
c.
Beijing Key Laboratory of Key Technologies for Natural Drug Delivery and Novel Formulations, Institute of Materia Medica, Chinese Academy of Medical Sciences & Peking Union Medical College, Beijing 100050, China
d.
University-Town Hospital of Chongqing Medical University, Chongqing 401331, China
Received Date:
15 November 2025 Accepted Date:
05 February 2026 Revised Date:
03 February 2026 Available Online:
15 July 2026
Abstract:
Skin aging is a multifactorial biological process driven by oxidative stress (OS), chronic inflammation, cellular senescence, mitochondrial dysfunction, and non-enzymatic glycation (NEG), and is markedly accelerated by environmental stressors such as ultraviolet radiation (UVR) and air pollution. Natural bioactive compounds, including flavonoids, polyphenols, terpenoids, alkaloids, vitamins, polysaccharides, peptides, and indole derivatives, have demonstrated considerable potential in mitigating these processes through multi-target antioxidant, anti-inflammatory, anti-photoaging, and extracellular matrix (ECM)–protective mechanisms. However, their clinical and cosmetic translation is often limited by poor solubility, chemical instability, and insufficient skin penetration. Rather than providing a simple compilation of existing studies, this review offers a mechanism-oriented and delivery-driven synthesis of recent advances in natural anti-skin-aging research. We systematically integrate current knowledge on the molecular drivers of skin aging with a critical comparison of emerging transdermal and nanotechnology-based delivery systems, including lipid-based, polymer-based, inorganic/hybrid, and nucleic acid–assembled nanocarriers. Particular attention is given to how rational carrier design modulates stability, permeability, skin-layer targeting, and controlled release behavior. Furthermore, this review critically discusses current limitations, contradictory findings across studies, and key translational challenges, and identifies promising future research directions, such as stimuli-responsive systems, skin-layer-specific targeting, and mechanism-guided material selection. By bridging biological mechanisms with delivery material innovation, this work provides a forward-looking framework for the development of safer, more effective, and scientifically grounded anti-skin-aging therapeutics and cosmeceuticals.
Skin aging is the most notable manifestation of human aging and a significant consequence of a decline in skin functionality [1]. The skin, as the largest organ in the human body, plays several pivotal roles, including protecting the body from external damage, regulating body temperature, and sensing external stimuli [2]. However, the structure and function gradually deteriorate with age due to intrinsic aging processes and extrinsic environmental factors, resulting in wrinkles, sagging, pigmentation, and other visible aging signs that affect both appearance and normal physiological function. Skin aging is a complex, multifactorial process involving interactions at cellular and molecular levels, including OS, inflammatory responses, DNA damage, mitochondrial dysfunction, and cellular senescence [3]. A comprehensive understanding of these mechanisms provides a solid theoretical foundation for developing effective anti-aging strategies.
Natural products, ranging from plant extracts to marine bioactive compounds, have attracted considerable attention for their anti-aging potential, due to their antioxidant [4], anti-inflammatory [5], anti-photoaging [6], anti-glycation [7], and collagen-promoting properties [8]. Despite promising bioactivities in preclinical models, many natural anti-aging compounds show limited clinical efficacy due to intrinsic physicochemical limitations, including poor aqueous solubility, chemical instability, and insufficient skin penetration imposed by the stratum corneum (SC) barrier. Recent advances in transdermal and nanotechnology-based drug delivery systems (DDSs) provide effective strategies to overcome these challenges by enhancing cutaneous permeation, stabilizing labile molecules, and enabling controlled or targeted release.
Although numerous reviews have summarized natural anti-aging agents or emerging delivery platforms, few have systematically integrated the biological mechanisms of skin aging with the rational design principles of advanced DDSs. This review addresses this gap by offering a mechanism-oriented and delivery-driven synthesis, critically comparing major nanocarrier platforms and highlighting emerging trends such as stimuli-responsive systems and skin-layer-specific targeting. By explicitly linking aging-related biological targets with delivery strategies, this work provides a forward-looking framework to facilitate the translation of natural anti-skin-aging compounds into clinically and commercially viable applications.
2.
Skin aging
Skin, a mirror that reflects the aging process of the human organism [9] and is therefore an important quality of the external attractiveness of humans. Skin aging is a complex biological process and the most intuitive manifestation of aging, defined by continuous and inevitable histological and morphological changes in the skin. During aging, the external skin shows a variety of notable clinical phenotypes, such as dry skin, increased fine lines, decreased elasticity, and pigmentation, and further benign tumors such as hemangiomas, fibroma moles, and seborrheic keratoses may appear.
As aging progresses, notable signs also appear in the internal tissues of the skin, such as thinning of the epidermis and dermis [10], flattening of junctions, and weakening of adhesions. Further, at the cellular level, the number of healthy keratinocytes, melanocytes, and Langerhans cells (LCs) in the epidermis declines [11], whereas the number of aging keratinocytes and melanocytes increases [12]. In the dermis, the activity and proliferative ability of healthy fibroblasts gradually decrease, and many senescent fibroblasts are produced. In addition, as extracellular matrix (ECM) proteins begin to degrade, the amounts of collagen (especially type Ⅰ and Ⅲ collagen) and elastin decrease and become loose [13], and the number of hair follicles and sebaceous glands decreases (Fig. 1).
Figure 1
Figure 1.
Compare aging skin and young skin at the tissue, internal structure, and cellular levels. (A) ECM, composed of collagen, elastin, and reticular fibers, provides structural support for the dermis. Aging or UV exposure leads to ECM degradation and reduced synthesis, causing wrinkles and sagging. (B) SC, the outermost epidermal layer, consists of anucleate keratinocytes and intercellular lipids, forming a "brick wall" barrier. Aging slows SC turnover, reduces moisture, and weakens barrier function, leading to dry and sensitive skin. (C) Aged skin exhibits wrinkles, thinning, flattened dermal-epidermal junctions, dermal papillae atrophy, decreased cell density, collagen breakdown, and elastin loss. (D) Young skin shows a smooth surface, tight dermal-epidermal junctions, dense cellularity, and well-organized collagen and elastin fibers. Created in https://BioRender.com.
Skin aging is jointly affected by a variety of endogenous and exogenous factors, among which age and genetic factors constitute intrinsic factors, whereas the environment and diet constitute extrinsic factors [14].
Intrinsic skin aging, also known as natural aging, is caused by factors such as time, genetics and hormone levels as age increases. Chronological aging represents an inevitable, time-associated disruption of tissue homeostasis that leads to marked functional decline in the skin. This intrinsic aging process is largely dictated by individual and ethnic genetic backgrounds [15] and is further driven by multiple time-dependent physiological alterations. Notably, the chronological aging process is stable, with morphological and structural changes in the skin occurring in a gradual and uniform manner. Abdominal skin samples were obtained from the autopsies of 25 patients to allow the researchers to examine histological features associated with chronological aging [16]. The results showed that epidermal thickness, interdigitation, and mitotic activity declined with age, whereas melanocyte numbers increased. In the dermis, papillary thickness increased, likely due to accumulation of glycosaminoglycans (GAGs) and expansion of mast cells, while the reticular dermis exhibited reduced thickness, accompanied by a decline in GAGs and the elastic fibers. A study examined chronological aging of rat skin by analyzing structural and redox changes from 3 days to 21 months of age. Young (3-day-old) and old (21-month-old) skin exhibited higher reactive oxygen species (ROS)-scavenging enzyme activities compared to adults (3-month-old). The results suggest that chronological aging involves a gradual decline in redox flexibility and repair capacity rather than an overall loss of antioxidant activity [17]. Overall, chronological aging is a gradual, time-dependent process characterized by progressive structural, morphological, and functional changes across the epidermis and dermis. Chronological aging also effects redox homeostasis, leading to a slow decline in the skin's flexibility and repair capacity rather than a uniform loss of antioxidant activity. Beyond chronological aging, intrinsic skin aging is influenced by racial and anatomical skin characteristics and endocrine regulation [18–20], particularly sex hormone–mediated signaling [21] that governs skin structure and aging dynamics.
Extrinsic aging is an inevitable process defined by intrinsic aging of the body coupled with long-term exposure to the external environment or changes in diet and rest. UVR, which causes photoaging [22], is the primary external driver of external skin aging, accounting for approximately 80% of facial aging cases (Fig. 2A) [23]. The degree of photoaging mainly depends on the duration and intensity of solar radiation. Clinically, photoaging can manifest as skin wrinkles, sagging, roughness, yellowing or shallow skin color, capillary dilation, pigment spot formation, and even skin cancer (Fig. 2B) [24–26].
Figure 2
Figure 2.
(A) The solar spectrum consists of electromagnetic rays of varying wavelengths that penetrate the skin to different depths. (B) UVR damages the skin through multiple mechanisms. UVB induces DNA damage and promotes skin cancer. Keratinocytes release alpha‑melanocyte‑stimulating hormone (α-MSH) and endothelin‑1 (ET‑1), stimulating melanocytes to produce melanin, resulting in pigmentation and age spots. ROS generation triggers inflammatory responses by promoting the release of TNF-α, IL-1β, and IL-6 from keratinocytes, fibroblasts, and LCs, leading to sunburn and skin inflammation. ROS also upregulates MMPs (MMP-1, MMP-3, MMP-9), causing collagen degradation, wrinkles, and accelerated skin aging. Created in https://BioRender.com.
Controlling the release of iron from cytosolic ferritin is the key to preventing ultraviolet A (UVA)-induced inflammation. Smith et al. found that UVA irradiation of human dermal fibroblasts and endothelial cells induces a transient increase in cytosolic Fe(II) levels [27]. Moreover, UVA irradiation also increases H2O2 in dermal endothelial cells, thereby increasing superoxide levels and initiating an inflammatory response. Further, a combination of UVA exposure and glycosylation can upregulate the mRNA expression of matrix metalloproteinases (MMPs), including MMP-1 and MMP-3, which enhances the degradation of ECM proteases, affects ECM activity, and promotes the expression of cellular inflammatory factors [28]. UVB, as a strong mutagen [29], exerts strong biological effects on the epidermis and some effects on the papillary dermis and causes sunburn and erythema [30]. UVB can directly induce DNA mutations and form cyclobutane pyrimidine dimers, thereby damaging the skin DNA [31]. Notably, UVB exposure can significantly increase ROS production in human epidermal keratinocytes, which causes OS, activates the arachidonic acid pathway, induce inflammatory responses [32], and ultimately lower cell viability [33]. Finally, although UVC has the strongest mutagenicity, it is absorbed by the ozone layer before it reaches the Earth's surface, thereby limiting its impact on skin aging [10]. Visible light (VL) and infrared radiation (IR) induce MMP-1 and MMP-9 expression and reduce collagen production [34]. In addition, VL and IR exert different effects on skin photobiology at different wavelengths. For example, red light and IRA do not affect keratinocyte proliferation, whereas blue light (412, 419, and 426 nm) inhibits proliferation and promotes keratinocyte differentiation [35]. Different VL and IR wavelengths also exert different effects on the gene expression profiles of fibroblasts [36]. Schieke et al. showed that exposure to IR deregulates metalloproteinase expression in primary human fibroblasts [37]. Further, animal models exposed to IR for long periods also develop the typical clinical symptoms of photoaging [38]. Extrinsic skin aging is further driven by environmental and lifestyle factors, including air pollution [39,40], cigarette smoking [41–43], dietary imbalance [44,45], thermal stress [46,47], psychological stress [48,49], and sleep deprivation [50,51], which collectively accelerate OS, inflammation, epigenetic aging, and ECM degradation.
2.2
Mechanisms of skin aging
ROS mainly originate from cellular oxidative metabolism and UVR. Sources of cellular oxidative metabolism include a variety of enzymes and non-enzymatic substances such as the mitochondrial electron transport chain, cyclooxygenase, peroxisomal oxidase, NADPH oxidase, and lipoxygenase [10]. The signaling pathways involved in ROS production include the nuclear factor kappa B (NF-κB) signaling pathway and the activator protein-1 (AP-1) pathway stimulated by mitogen-activated protein kinase (MAPK) (Fig. 3). Inhibitor of kappa B alpha (IκB-α) is an inhibitory protein of the NF-κB signaling pathway. Excessive ROS in the human body inhibits IκB-α synthesis, thereby activating the NF-κB signaling pathway and leading to increased levels of tumor necrosis factor-alpha (TNF-α) and MMPs [52]. MMPs degrade the ECM, mainly collagen and elastin, to promote skin aging. Activated NF-κB also affects the expression of various cytokines that mediate inflammation [53], including interleukin-6 (IL-6), IL-8, and cyclooxygenase-2 (COX-2). IL-6 accelerates the production of MMP-1, the main protease that initiates collagen cleavage. In addition, activated NF-κB can also regulate heme oxygenase-1 (HO-1) expression, which indirectly increases the level of free iron in the cells and promotes the further production of ROS via the Fenton reaction [54]. ROS activate AP-1 expression involving cellular Fos (c-Fos) and cellular Jun (c-Jun) in fibroblasts and keratinocytes through the MAPK pathway and induce MMPs synthesis, especially MMP-1, MMP-3, and MMP-9 [52]. Transforming growth factor-beta (TGF-β) is a key factor in the synthesis of type Ⅰ procollagen. Activated AP-1 also downregulates TGF-β type Ⅱ receptor expression [55], which attenuates the TGF-β/SMA and MAD related protein (Smad) signaling pathway, inhibits collagen synthesis, and promotes skin aging. In addition, activated NF-κB and AP-1 can reduce the transcription levels of tissue inhibitors of MMPs and specific inhibitors of MMPs [56], thereby elevating MMPs levels, degrading collagen and elastin, and promoting secretion of the senescence-associated secretory phenotype (SASP). AP-1 is a key transcription factor regulating the expression of most MMPs, and it becomes activated upon elevated ROS production [57]. Finally, NF-κB and AP-1 are involved in the skin cancer pathogenesis in older adults as regulators of the balance of cell proliferation and apoptosis [58].
Figure 3
Figure 3.
Sources of ROS and the signal pathways involved. Exogenous ROS include: UVR, air pollution, cigarette smoke, and heavy metal exposure. Endogenous ROS include the electron respiratory transport chain in mitochondria, NADPH oxidase, endoplasmic reticulum stress, and peroxisomes. The signal pathways involved in ROS mainly include: AP-1 signaling pathway, TGF-β/Smad signaling pathway, NF-κB signaling pathway, Nrf2/ARE signaling pathway, AGEs-RAGE signaling pathway, mTOR signaling pathway, p53/p21/p16 signaling pathway, etc. Solid arrows indicate activation, and dotted arrows indicate inhibition. Created in https://BioRender.com.
Cellular senescence is a stress-induced cellular state characterized by irreversible cell cycle arrest accompanied by distinctive secretory, metabolic, and macromolecular alterations [59]. Cyclin-dependent kinases (CDKs), a major class of serine/threonine protein kinases that regulate cell cycle progression, are activated after phosphorylation. The major CDKs inhibitors responsible for cell cycle arrest during aging are encoded by CDKN2A (p16INK4A), CDKN2B (p15INK4b), and CDKN1A (p21WAF1/CIP1) loci [60]. Multiple signaling pathways can lead to cellular senescence, and cell cycle arrest is caused by the p53/p21WAF1/CIP1 and p16INK4A/ retinoblastoma protein (pRB) tumor suppressor pathways, both of which are activated in response to cellular stressors [61]. These two pathways can be activated in parallel or can induce cell cycle arrest independently of each other (Fig. 3) [62]. p21WAF1/CIP1 is regulated by direct cross-activation of p53 but can also be activated by p53-independent mechanisms through tumor necrosis factor beta (TNF-β) and specificity protein 1 (Sp1) [63], thereby inactivating CDK2. In CDK2 absence, pRB remains in its active, hypophosphorylated form and binds to E2F transcription factor 1 (E2F1), thereby inhibiting the transcriptional targets of E2F1 and causing cell cycle arrest in the G1 phase [64]. p16INK4A, a cell cycle regulator, is the only specific aging marker and is widely used to detect aging in vivo [65]. p16INK4A directly interacts with and inhibits CDK4/6 [66], thereby activating pRB. Moreover, p16INK4A is responsible for stress-induced premature aging; however, this effect is not irreversible. If senescence is triggered by p16INK4A activation, silencing of p16INK4A through promoter methylation or deletion of the p16INK4A site allows cells to resume the cell cycle [64]. Nevertheless, p16INK4A overexpression has been detected in aging human skin, suggesting a possible link between this inhibitory protein and aging.
3.
Anti-skin aging properties of natural products
Natural products from animals, plants, and microorganisms often confer antioxidant, anti-inflammatory, neuroprotective, liver-protective, and other effects in human cells [67]. As more people are pursuing green and healthy natural beauty in modern society, many natural products are widely used to improve skin aging and health.
3.1
Flavonoids
Flavonoids share a phenylchromen-4-one core, with a phenyl ring at C2 or C3 forming flavone or isoflavone backbones. Structural variations, including a C4 ketone, C2-C3 saturation or double bond, and hydroxy or methoxy substitutions on rings, give rise to flavanones, flavanols, flavonols, flavones, anthocyanidins, and isoflavones [68]. de Souza Farias et al. analyzed the conformational and electronic properties of seven flavonoids to identify structural features underlying their antioxidant activity, finding that increased planarity-stabilized by hydrogen bond between O3H and the ketone oxygen or between O1 and HC6'-enhances the aromaticity of ring C and strengthens their antioxidant potential [69]. Hydroxyl groups (-HO) are critical for the anti-inflammatory activity of flavones, with C-5 and C-4' -OH enhancing and C-6, C-7, C-8, and C-3' -OH reducing their efficacy. Additionally, a C2-C3 single bond and -OH at C-3 and B-ring positions diminish flavone aglycone activity, which primarily acts via NF-κB, MAPK, and c-Jun N-terminal kinase–signal transducer and activator of transcription (JNK-STAT) pathways, targeting kinases, aryl hydrocarbon receptor (AhR), G-protein coupled receptors, and estrogen receptor (ER) [70].
Baicalein, a flavone glycoside, exerts antioxidant effects by scavenging ROS through various mechanisms, especially by inhibiting NF-κB activity [71] and the expressions of various inflammatory cytokines and enzymes [72]. Wang et al. [73] investigated baicalein's protective effects against DNA oxidative damage, showing that it scavenges free radicals (·OH, (2,2-diphenyl-1-picrylhydrazyl) radical (DPPH·), 2,2′-azinobis-(3-ethylbenzothiazoline-6-sulfonic acid) radical cation (ABTS+·) and chelates metal ions (Fe2+). Baicalein also binds to DNA minor grooves, forming a stable complex that preserves the double-helix structure and protects DNA from oxidative damage without altering its conformation. This protective effect involves activation of nuclear factor erythroid 2-related factor 2 (Nrf2) and HO-1 pathways. Sun et al. knocked out Rev-erbα/Bmal1 to gauge its role in regulating skin aging and demonstrated that Scutellaria baicalensis extract antagonized Rev-erbα (nuclear receptor subfamily 1 group D member 1, NR1D1), upregulated Bmal1 (brain and muscle ARNT-like protein 1, ARNTL), and prevented skin aging in mice [74]. Wang et al. found that when Scutellaria baicalensis extract was administered to the backs of mice, it effectively inhibited RAW264.7 cell survival at the administration site and reduced the number of F4/80 macrophages [75]. Notably, Scutellaria baicalensis extract can reduce the excessive activation and infiltration of macrophages, thereby alleviating Imiquimod-induced inflammation and skin damage. Luteolin (LU) inhibits pro-inflammatory factors (IL-1β, IL-6, IL-8, IL-17, IL-22, TNF-α, and COX-2) and regulates various signaling pathways (such as NF-κB, Janus kinase–signal transducer and activator of transcription (JAK-STAT), and Toll-like receptor (TLR)), exerting antioxidant and anti-inflammatory effects in keratinocytes, fibroblasts, and various immune cells [76]. Hajem et al. treated human skin dermal fibroblasts with purple tulip extract and observed an obvious induction of immune response-related genes 3 h later [77], including cytokines, chemokines, and cell factor receptors. Further, prostaglandin-endoperoxide synthase 2 (PTGS2) expression, which is responsible for inflammatory prostaglandin production, was also induced after 3h of treatment and remained overexpressed after 24 h. Chaikul et al. found that green tea ingredients have anti-aging effects on skin, including significant inhibition of melanin production by limiting the activities of tyrosinase and tyrosinase-related protein 2 [78]. Further, green tea has potent antioxidant properties and exhibits a significant MMP-2 inhibitory effect. Boonpisuttinant et al. found that the extract of giant Indian gooseberry, which has high total phenols, total flavonoids, and quercetin contents, exhibited good antioxidant activity and reduced melanin production by inhibiting tyrosinase activity and promoting collagen biosynthesis [79].
3.2
Polyphenols
Phenolic compounds consist of benzene rings bearing hydroxyl groups, often conjugated to sugar moieties, and span a spectrum from simple monomeric forms to complex polymeric structures [80]. Evaluation of the structure-activity relationship of phenolic antioxidants revealed that para-substitution with ethyl or n-butyl groups, instead of a methyl group, enhances their antioxidant activity [81]. Nevertheless, the degree of hydroxylation and the number and configuration of electron-donating substituents (-OH and -OCH3) play a critical role in determining antioxidant activity [82].
Resveratrol (RES) reduces the expression of AP-1 and NF-κB factors to activate Nrf2 and increases type Ⅰ, Ⅱ, and Ⅲ collagen in fibroblasts [83]. RES inhibits melanin production, induces Sirtuins1 expression, and plays a protective role against skin aging. In a study on the apoptosis of HaCaT cells with normal or overexpressed HSP27 under UVB irradiation, RES regulated HSP27 expression, upregulated Bax expression, and downregulated B-cell lymphoma 2 (Bcl-2) expression to control the activation of p65 and caspase-3 [84]. The survival rate of HaCaT cells pretreated with different doses of RES was significantly improved (up to 94%). Kim et al. showed that oral or topical administration of RES could reduce UVB-induced epidermal thickness by ~55% [85]. At the cellular level, RES reduces MMP-1 and MMP-9 expression, activates the antioxidant signaling pathway mediated by the intracellular Nrf2/HO-1 signaling pathway, and protects cells from UV damage. Further, RES can target TLR signaling pathways such as TLR2, TLR3, and TLR4 [86,87]. The accumulation of advanced glycation end products (AGEs) in 3D skin models is associated with reduced epidermal stratification and the reorganization of collagen in the dense layer of cells in the upper dermis. Using in vitro 3D skin model experiments, Markiewicz et al. demonstrated that the use of RES derivatives in dermal fibroblasts reduces glycosylation-induced AGEs and increases collagen expression and epidermal stratification [88]. Shin et al. found that protocatechuic acid exhibits high antioxidant activity in vitro, induces type Ⅰ collagen synthesis in human dermal fibroblasts and skin explants, and inhibits MMP-1 secretion by UVA-irradiated human dermal fibroblasts [89]. Lee et al. used UVB/H2O2-induced human dermal fibroblast damage as an in vitro model and UVB/ H2O2-induced photoaging in C57BL/6J nude mice as an in vivo model and reported that galangal alleviates loss of cell viability induced by UVB/H2O2 and exerts anti-photoaging and antioxidant effects by promoting the sirtuins1/peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α)/Nrf2 signaling pathway [90].
3.3
Terpenoids
Terpenoids are a remarkable class of natural products derived from linear polyprenyl diphosphate precursors [91]. A research team examined the antioxidant properties of terpenes to elucidate the structure-activity relationship governing their radical-scavenging capacity, particularly the contribution of conjugated double bonds [92]. The results demonstrated that terpenes containing π-bond conjugation exhibited markedly stronger chain-breaking antioxidant activity in both DPPH·, and ABTS+· assays, with α-terpinene, ocimene, pulegone, and farnesene identified as the most potent scavengers. Xu et al. investigated protostane-type terpenoids isolated from Alisma orientalis to evaluate their antiproliferative activities and elucidate key structure-activity relationships [93]. Based on the results, they concluded that the degree of C-16 oxidation and the double bond between C-13 and C-17 may be significant in anti-proliferative activities.
Buranasudja et al. revealed that the triterpenoid components in Centella asiatica callus extract (CACE) exhibit strong antioxidant and anti-aging properties, wherein they attenuate H2O2-induced cytotoxicity in human dermal fibroblasts [94]. In addition, CACE inhibited the induction of MMP-9 after H2O2 exposure. Kim et al. evaluated the anti-aging effects of tissue cultured from wild trigonelline and its major ginsenoside in TNF-α stimulated human dermal fibroblasts by inhibiting MMP-1 secretion and increasing collagen type Ⅰ alpha 1 chain (COL1A1) secretion [95]. Notably, trigonelline 1 and trigonelline 3 inhibited the MMP-1 secretion in human dermal fibroblasts stimulated by TNF-α, and trigonelline 2 could promote COL1A1 secretion, but the effect was not significant. Liu et al. found that the ginsenoside Rk1 significantly reduced OS by inhibiting ROS overproduction and enhancing the activity of antioxidant enzymes, and that Rk1 inhibited the phosphoinositide 3-kinase (PI3K)/protein kinase B (Akt)/NF-κB signaling pathway both in vitro and in vivo [96]. Ginsenoside F1 reduces IL-6 secretion in a dose-dependent manner, reaching the maximum inhibition rate (~57%) at 30 µg/mL. It also reduced IL-8 mRNA and protein expression and inhibited the SASP production by interfering with DNA-damage response signaling (the sustained DNA damage response signal required to maintain SASP) [97]. Ginsenoside prevents UVB-induced skin photoaging in multiple ways. Specifically, Rb1, Rb3, Rg3, Rk1, and Rh2 inhibit UVB-induced ROS to prevent or reduce oxidative skin damage and inflammatory responses [98]. Liu et al. used HaCaT keratinocytes exposed to UVB irradiation as an in vitro model and BALB/c nude mouse back skin exposed to UVB irradiation as an in vivo model, and found that the ginsenoside Rk1 reduced UVB-induced inflammatory responses by downregulating the secretion of multiple pro-inflammatory cytokines and could also inhibit collagen degradation by reducing MMP-3 and MMP-9 expression. Therefore, ginsenoside Rk1 may be an effective natural anti-photoaging agent [96]. Further, Li et al. used a BALB/c hairless mouse model to show that ginsenoside improved symptoms of UVB-induced epidermal barrier dysfunction, such as increased epidermal thickness and transepidermal water loss (TEWL), reduced SC water content, and inhibited phosphorylation of c-Jun N-terminal kinase (JNK), p38, and Rb3 in HaCaT cells [99].
3.4
Alkaloids
Alkaloids are naturally occurring organic compounds containing nitrogen within a heterocyclic ring, typically exhibiting basic properties. Each alkaloid molecule contains at least one nitrogen atom, which is often tertiary and less commonly quaternary. The heterocyclic rings can fuse with each other or with other ring systems, resulting in alkaloids with polycyclic or macrocyclic structures. Račková et al. evaluated the antioxidant activities of alkaloids from Mahonia aquifolium [100]. The study focused on how structural features, particularly phenolic hydroxyl groups, electron-delocalization capacity, and lipophilicity, govern their radical-scavenging and antiperoxidative properties. The results demonstrated that hydroxylated alkaloids exhibited substantially stronger antiradical and membrane-protective activities than berberine, with efficacies comparable to stobadine and superior to Trolox, highlighting the synergistic contribution of phenolic -OH groups and favorable lipophilicity to antioxidant potency. A comprehensive analysis of 17 classes of isoquinoline alkaloids was performed to delineate key structure-activity relationships. The review revealed that quaternary nitrogen and a C2-C3 methylenedioxy substitution markedly enhance antimicrobial and antiviral potency, particularly within the N-methyltertrahydroprotoberberine, protoberberine, and benzophenanthridine subclasses [101].
Li et al. established an in vivo OS model using mouse skin tissues exposed to UVR [102]. Subsequent experiments revealed that caffeine, an alkaloid, inhibits its main cellular target, the adenosine A2A receptor, and increases the protein level of Sirtuins3. Caffeine further protects the skin from OS-induced aging by activating 5′-adenosine monophosphate-activated protein kinase (AMPK), which activates autophagy and promotes telomerase reverse transcriptase (TERT) expression at the mRNA and protein levels to extend telomere length and prevent cell aging [103]. In an experimental model of normal human dermal fibroblasts cultured in vitro, the alkaloid centcyanine activated the KL gene expression and the related protein, Klotho, in normal human dermal fibroblasts and slowed down their replication and aging process [104]. Naikoo et al. explored the protective effects of trigonelline against UVB-induced photodamage in human dermal fibroblasts (Hs68 cells) and BALB/c mice and found that the survival rate of UVB-exposed cells treated with trigonelline was significantly higher than that of untreated irradiated cells [105]. Further, trigonelline treatment successfully reduced OS, restored Ca2+ homeostasis, re-established endoplasmic reticulum function, and prevented apoptotic cell death. Freitas et al. obtained four benzylisoquinoline alkaloids by separating and purifying Ocotea paranapiacabensis [106]. Subsequently, an in vitro anti-glycation test and measurement of the fluorescence of total AGEs revealed that the alkaloids in O. paranapiacabensis exhibited a strong inhibitory effect on AGEs and protected the skin from aging.
3.5
Vitamins
Vitamins are essential organic compounds required in small amounts for normal physiological functions, many of which exhibit potent antioxidant properties. Their chemical structures, including hydroxyl, conjugated double bonds, and lipophilic characteristics, critically determine their ability to scavenge free radicals and protect biomolecules from oxidative damage [107].
As a form of vitamin B3, niacinamide is a water-soluble vitamin. Kwak et al. used two cell models, rapidly aging human breast cancer MCF-7 cells and senescent human fibroblasts, and found that nicotinamide treatment reduced oxidative damage in senescent cells [108]. Further, nicotinamide inhibits ROS production, reduces lipofuscin accumulation, inhibits senescence-associated β-galactosidase (SA-β-gal) activity, exerts anti-aging effects, and controls mitochondrial and cellular superoxide levels, with a stronger effect on cytoplasmic superoxide levels. Nicotinamide treatment suppressed the stress-induced elevation of cytokine and prostaglandin synthesis, restored lamin B1 levels that are known to decline during aging and are reduced by UVB exposure, and pretreatment with 5% nicotinamide reduced UV-induced erythema [109]. Shao et al. demonstrated that topical retinol (vitamin A1) significantly increased epidermal thickness by stimulating keratinocyte proliferation and upregulating c-Jun transcription factor [110], stimulated the proliferation of endothelial cells and the production of ECM (type Ⅰ collagen, fibronectin, and elastin) by activating dermal fibroblasts, and maintained ECM homeostasis by regulating the TGF-β/connective tissue growth factor (CTGF) signaling pathway. Further, Scott et al. demonstrated that oral administration of high doses of vitamin D3 shortly after UVB exposure reduced inflammation and induced epidermal barrier repair [111], thereby rapidly reversing UVB exposure-induced skin damage. Gref et al. covalently coupled vitamin C with squalene to form a new vitamin C-squalene bioconjugate and tested its bioactivity in skin explants using an ex vivo model [112]. Ten days after application, vitamin C-squalene significantly increased the thickness of the skin epidermis, preferentially promoted type Ⅲ collagen production in skin explants, and promoted glycosaminoglycan production.
3.6
Polysaccharides
Polysaccharides are widely distributed in plants, microorganisms (including fungi and bacteria), algae, marine organisms, and animals. The biological activities of various polysaccharides are closely related to their chemical structures, with molecular-level features playing a decisive role in mediating their functional effects [113]. Polysaccharides with lower molecular weight tend to display enhanced antioxidant activity, as the increased number of free hydroxyl groups allows them to more effectively donate hydrogen atoms and neutralize free radicals [114]. In addition, the relationship between glycosidic-bond type and activity suggests that polysaccharides with β-glycosidic linkages generally exhibit higher biological activity [115], whereas those with α-glycosidic linkages are often inactive due to hydrolysis by α-glucoamylase in the human body under certain conditions.
Fucoidan-rich polysaccharides extracted from Sargassum horneri were prepared and compositional characteristics evaluated, and their anti-aging potential was examined using antioxidant assays, UVB-induced human fibroblasts, and a short clinical application test [116]. Sargassum horneri fucoidan extract displayed marked DPPH radical-scavenging activity, enhanced procollagen synthesis, suppressed UVB-induced MMP-1/3 expression in fibroblasts, and clinically improved epidermal barrier function with reduced TEWL, supporting its potential as a cosmetic anti-aging ingredient. A novel branched β-(1,3)-D-glucan polysaccharide (DAG-1) was extracted from Durvillaea antarctica using subcritical water extraction and ultrafiltration, and characterized by GC-MS and 1D/2D NMR [117]. DAG-1 exhibited anti-skin-aging effects in D-gal-induced mice by preserving superoxide dismutase (SOD) activity, reducing malondialdehyde (MDA) accumulation, and maintaining hydroxyproline levels. In vitro, it alleviated fibroblast senescence, modulated TNF-α and IL-6 production, and potentially regulated mast cell activity via KIT proto-oncogene receptor tyrosine kinase (c-KIT)/KITLG (KIT ligand, also known as stem cell factor) signaling. Hu et al. isolated and structurally characterized a purified polysaccharide-1 (P1) from Sargassum fusiforme, which possesses a defined β-ManA/α-GulA backbone and terminal α-L-fucose linkage [118]. In UVB-irradiated HaCaT cells, P1 markedly suppressed the expression of MMP-1/-3/-9 and reduced ROS as well as pro-inflammatory cytokines (TNF-α, IL-6, IL-1β), thus attenuating collagen degradation, OS, and inflammation associated with photoaging. Although marine-derived bioactive compounds hold great potential, their application is often limited by supply and regulatory constraints, which may be alleviated through nanotechnology-enabled stabilization and delivery strategies [119].
3.7
Bioactive peptides
Bioactive peptides typically consist of 2–20 amino acid residues per molecule, although some may contain more than 20 residues, with molecular masses generally below 6000 Da [120]. Bioactive peptides are latent within the primary sequences of their parent proteins and remain inactive until they are released through enzymatic, chemical, or microbial hydrolysis, which allows them to exert diverse physiological and functional activities [121]. It is worth noting that these peptides exhibit pronounced scavenging activities toward reactive oxygen and nitrogen species, largely owing to the enrichment of polar and aromatic amino-acid residues in their sequences. This biochemical characteristic underpins their potential utility as functional dietary components or therapeutic candidates for mitigating aging-associated processes [122].
Certain bioactive peptides are capable of modulating multiple biological processes within both the epidermal and dermal compartments of the skin, thereby influencing cutaneous function through the enhancement of collagen synthesis and partial restoration of ECM components, ultimately contributing to the amelioration of wrinkles, fine lines, and overall skin tone [123]. Castro-Jacome et al. prepared peptide extracts from white sorghum (Sorghum bicolor) kafirins by alcalase hydrolysis [124], followed by ultrafiltration to yield two fractions of < 1 kDa (PE-1) and 1–3 kDa (PE-3). In UVB-exposed human organotypic skin cultures, both extracts markedly attenuated oxidative damage by restoring SOD and glutathione peroxidase (GPx) activites, maintaining or increasing catalase (CAT) activity, and suppressing pro-inflammatory cytokines and matrix-degrading enzymes, with PE-1 exhibiting superior protective efficacy comparable to glutathione. He et al. investigated the pentapeptide Ala-Tyr-Ala-Pro-Glu (AYAPE), previously identified from Isochrysis zhanjiangensis, in UVB-induced HaCaT keratinocytes and H2O2-treated human dermal fibroblasts, and demonstrated that AYAPE alleviates UVB-induced photoaging by reducing DNA damage, intracellular ROS, MAPK/AP-1 activation, and MMP-1 production, while also attenuating oxidative-stress-driven fibroblast senescence through p53-dependent mechanisms, thereby suggesting its potential as an anti-aging peptide [125].
3.8
Biogenic amines
Biogenic amines are low-molecular-weight organic compounds derived primarily from amino acids, characterized by one or more amino groups attached to an aliphatic, aromatic, or heterocyclic carbon backbone. Their chemical structures, including the number and position of amino groups, degree of saturation, and presence of functional substituents, play a critical role in determining their biological activities [126]. Electron-donating groups and conjugated systems can enhance free radical scavenging, while specific amine configurations may interact with cellular receptors or enzymes involved in OS response, inflammation and cellular senescence pathways [127].
Li et al. established a macrophage-senescent skin cell co-culture system using THP-A-derived macrophages and tert-butyl hydroperoxide-induced senescent HaCaT and HF-1 cells [128], and assessed senescence markers by SPiDER-βgal staining and qPCR to evaluate the effects of carnosine (CAR). CAR activated the Akt2 signaling pathway, upregulated cluster of differentiation 36 (CD36) and receptor for advanced glycation end products (RAGE), enhanced macrophage phagocytic capacity, and thereby promoted the clearance of senescent keratinocytes and fibroblasts, indicating its potential to reverse skin aging. Girardi et al. established an ex vivo human skin model exposed to solar-simulated radiation (300–750 nm) to evaluate the photoprotective effects of topically applied L-CAR (0.2% and 2%), assessing ROS, AGE formation, and gene expression related to OS, inflammation, pigmentation, and photoaging [129]. L-CAR effectively reduced ROS and AGE accumulation and mitigated the modulation of 11 selected biomarker genes, demonstrating its capacity to protect skin from solar radiation-induced damage. Radrezza et al. employed a quantitative proteomic approach to analyze skin proteome changes in hairless mic exposed to UVA and to evaluate the preventive effects of CAR [130]. CAR pre-treatment mitigated UVA-induced alterations in 252 proteins, preserving pathways related to integrin-linked kinase, calcium signaling, fibrogenesis, cell migration, filament formation, and mitochondrial function, highlighting its protective effect against UVA-induced proteomic perturbations. Kim et al. investigated the effects of Streptococcus-secreted metabolites on human skin using metagenomic analyses [131], isolated bacterial strains, and treatments with their culture supernatants on skin cells and ex vivo skin. Streptococcus-derived spermidine enhanced skin structure and barrier function by upregulating collagen and lipid synthesis, leading to improved elasticity, hydration, and desquamation, indicate ng a microbiome-mediated anti-aging effect.
4.
Transdermal DDSs for natural products
Natural products have gained widespread attention because of their low toxicity, few side effects, wide range of biological activities, mild medicinal effects, and low development costs. However, some natural products exhibit easy degradation, poor hydrophilicity and lipophilicity, and difficulty penetrating the skin barrier, resulting in low bioavailability. Therefore, many studies have focused on DDSs to increase the quantity and rate of active ingredient delivery to target sites to improve drug efficacy.
4.1
Skin barrier and transdermal DDS
The cell layer connecting the SC, the outermost layer of the skin, and the epidermis constitutes the main resistance to transdermal transport (Fig. 4) [132]. The SC is the main component of the skin barrier and comprises dead cells and keratinocytes with a highly insoluble cornified envelope and keratin filaments, creating a "brick wall" structure [133]. Further, the smaller intercellular spaces because of the desmosomes and tight junctions in the SC limit the permeability of macromolecular drugs and proteins [134]. In addition, the keratinized envelope and intercellular lipid layer of keratinocytes are fat-soluble, thereby greatly limiting the penetration of water-soluble drugs into the skin. Only drugs with appropriate lipid solubility bypass keratinocyte diffusion through the lipid layer [135]. Consequently, the design of effective transdermal delivery systems must consider both the lipid solubility of drugs and strategies to overcome the SC's densely packed keratinized structure. Tight junctions of the epidermis are secondary barriers to the skin [136], and the normal life cycle of epidermal cells is critical for skin barrier function. New epidermal cells gradually mature and form keratinocytes, contributing to the continuous renewal of SC and the maintenance of its integrity. Therefore, penetrating the barrier function of the tight junctions in the SC and epidermis is the biggest challenge in transdermal drug delivery systems (TDDS) to deliver drugs to or across the skin and reach the target tissue [137]. This highlights the importance of developing rationally engineered delivery systems for bioactive natural compounds to achieve targeted anti-skin-aging effects. Topical skin preparations, such as ointments and portions made from animal, mineral, or plant extracts, have been commonly used since 3000 BC [138]. In 1950, the first transdermal drug delivery device, which allowed drugs to enter the bloodstream by penetrating the epidermis, was invented in the United States [139]. In 1979, the U.S. Food and Drug Administration approved the first transdermal preparation (Transderm-SCOP) [140], which could continuously release scopolamine over a 72 h period to prevent and treat nausea and vomiting caused by motion sickness. Consequently, the concepts of TDDS and transdermal therapeutic systems emerged. TDDS can enhance drug permeability, improve therapeutic effects, and reduce dosage and side effects, thereby expanding the scope of treatment and improving treatment selectivity, which are key aspects of drug development and treatment optimization.
Figure 4
Figure 4.
Schematic illustration of skin structure. (A) The skin consists of the epidermis, dermis, subcutaneous tissue, and appendages. (B) The epidermis includes, from top to bottom: SC, stratum granulosum, stratum spinosum, stratum basale, and the basement membrane. The SC is made up of anucleate corneocytes; The granulosum layer contains flattened keratinocytes; The spinosum includes keratinocytes and LCs; The basale consists of keratinocyte progenitor cells, melanocytes, and Merkel cells. (C) The SC has a brick-and-mortar structure, with dislocated corneocytes (15%–30%) embedded in a lipid matrix and connected by desmosomes. Created in https://BioRender.com.
4.2
Transdermal penetration promotion of natural products to treat skin aging
4.2.1
Physical penetration promotion
Microneedles (MNs) are small devices that can penetrate the SC and reach the depth of the dermis [141]. Microperforations via MNs can form hydrophilic microchannels in the SC, allowing drugs to quickly penetrate the SC barrier and reach the dermis [142]. Chiu et al. developed a MNs formulation based on poly-γ-glutamate that maintains the stability of epigallocatechin gallate (EGCG) and ensures its effective delivery to the skin to treat atopic dermatitis [143]. Notably, the amount of EGCG delivered to live skin by MNs (38.7 µg/cm2) was significantly higher than that of the untreated group (24.2 µg/cm2). Further, once-weekly microneedling treatments are at least as effective as the daily topical application of EGCG and l-ascorbic acid (as a stabilizer) solutions and significantly reduce the administration frequency and dosage required. Drugs are encapsulated in biodegradable polymers in dissolvable MN (DMNs) [144]. DMNs bypass the skin barrier by forming temporary microchannels, enabling effective delivery of nanocarriers into the skin [145]. DMNs do not leave hazardous needle waste and are not reusable, thereby improving their safety. Yang et al. used a single dissolvable MNs patch and applied a layer of horse oil around the adenosine (Ad)-loaded DMNs area to form an adenosine-loaded DMNs (HOS-Ad- DMNs) patch and compared its efficacy with that of an Ad-only dissolvable MNs patch [146]. Notably, the HOS-Ad DMNs patch significantly improved the dermal density, elasticity, and hydration of the skin and reduced wrinkles. Yerneni et al. used mild sonication to encapsulate curcumin and albumin into extracellular vesicles (EVs) and integrated the resulting Centella asiatica- EVs into the tip loading of DMNs [147]. Centella asiatica-EVs were five times more stable than natural curcumin in vivo, and Centella asiatica-EVs loaded into dissolvable MNs maintained their original beneficial cellular uptake and anti-inflammatory activities for at least 12 months. Fibroblast growth factors (FGFs) promote skin tissue remodeling and epithelial regeneration, but their penetration into the skin is limited by poor absorption. Yang et al. prepared a hyaluronic acid (HA) DMNs patch containing FGF-2 and FGF-21, which easily penetrated the skin of mice, releasing approximately 38% of the drug into the skin within 10 min and significantly improving skin inflammation and wrinkles caused by UVR within 2 weeks [148]. Sun et al. developed nanocomposites combining positively charged melittin (MEL) and negatively charged indocyanine green (ICG), embedded in dissolving MNs (MEL/ICG-HA@DMNs), to precisely bypass the SC and deliver therapeutics to superficial melanoma lesions [149]. The electrostatic complexation of MEL and ICG markedly reduced MEL's hemolytic activity while retaining membrane-disruptive function, and incorporation into MNs enhanced localized drug concentration and photothermal efficacy. Despite their potential, MN-based delivery systems face several unresolved challenges, including variability in skin penetration, limited macromolecule loading, and the need for precise control of release kinetics. Long-term safety, risk of local irritation or immune responses, manufacturing consistency, and regulatory standardization also remain major barriers to clinical translation for anti-skin-aging applications.
The iontophoresis method, which pushes charged molecules into the skin under electrodes of the same polarity using a small current to increase permeability [150], is often used with small molecules and charged drugs. Silva et al. studied the penetration of gallic acid after applying gel-type topical preparations to pig skin via cathodic iontophoresis and found that iontophoresis promoted the retention of gallic acid in the deeper layers of the skin and increased antioxidant activity in the skin after 20 min [151]. Nevertheless, several unresolved challenges continue to limit the broader application of iontophoresis. The narrow therapeutic window between effective and tolerable current densities raises concerns regarding skin irritation, particularly during repeated or long-term treatments. In addition, iontophoresis is largely restricted to small, charged molecules, and its efficiency is influenced by interindividual variability in skin resistance as well as formulation- and device-dependent parameters, complicating standardization and clinical translation.
Electroporation is an active delivery system that causes a short-term increase in cell membrane permeability using high-voltage (usually > 100 V) short-pulse electric fields that form tiny pores in the SC lipid bilayer, facilitating drug penetration into the skin. Zhang et al. explored the feasibility of electroporation-mediated topical delivery of l-ascorbic acid using a cream formulation and crystal suspension applied to a human cadaver and fresh surgical skin, respectively [152]. Notably, electroporation-mediated topical delivery was more effective on fresh skin than on cadaver skin and enhanced the penetration of l-ascorbic acid into the skin. However, several key challenges associated with electroporation-based transdermal delivery remain unresolved. The requirement for high-voltage pulses raises safety concerns, including patient discomfort, potential tissue damage, and difficulties in precisely controlling pore formation and recovery. Moreover, variability in skin properties, limited selectivity of pore creation, and the lack of standardized protocols hinder reproducibility and impede the clinical translation of electroporation for routine dermatological and anti-skin-aging applications.
4.2.2
Lipid-based nanocarriers
4.2.2.1
Liposomes (LPs) and niosomes
LPs are nano-sized lipid vesicles that encapsulate drugs and improve drug stability and transdermal efficiency by interacting with the skin lipid layer. Saewan et al. developed coffee berry extract (CBE)-loaded nanoliposomes to enhance stability, skin penetration, and anti-aging efficacy. By optimizing lipid composition and reducing particle size, the formulation achieved improved encapsulation efficiency, thermal stability, and sustained release, enabling deeper dermal delivery with reduced cytotoxicity [153]. Similarly, melatonin-loaded elastic liposomes incorporating sodium deoxycholate exhibited enhanced deformability, resulting in increased SC permeation and shortened lag time in vitro. Liu et al. co-encapsulated apigenin and doxycycline into flexible liposomes using sodium cholate and Tween-80 as edge activators, which enhanced vesicle deformability and skin penetration compared with conventional liposomes [154]. These structural modifications facilitated SC traversal and improved deep dermal delivery of both hydrophobic and hydrophilic cargos. Wang et al. [155] developed RES and CAR co-loaded LPs, further modified with chitosan and catechol-chitosan coatings, to enhance transdermal penetration and achieve synergistic anti-aging effects. By optimizing the RES: CAR mass ratio to 2:1, the engineered LPs exhibited excellent physicochemical stability, sustained-release behavior, and improved delivery of active compounds into the skin.
Compared with liposomes, niosomes are composed of non-ionic surfactants rather than phospholipids, conferring improved chemical stability, lower production cost, and enhanced suitability for transdermal delivery applications. Jin et al. developed optimized liposomes (GSL-7) and niosomes (GSN-4) for transdermal delivery of total ginsenosides to enhance anti-photoaging efficacy [156]. Formulation optimization significantly improved encapsulation efficiency, stability, and skin permeation, leading to increased transdermal flux and cutaneous retention. As a result, GSL-7 effectively alleviated UV-induced OS and inflammation in cellular and animal photoaging models. Li et al. designed a hyaluronic acid–modified, steareth-2-based niosomal system (EGT@HA-nio) that combines high mechanical deformability with active cellular targeting to overcome both the SC barrier and inefficient intracellular delivery [157]. By exploiting the intrinsic softness of the steareth-2 bilayer for intercellular penetration and HA-mediated targeting to circumvent OCTN-1 transporter dependence, the system enables direct mitochondrial accumulation of ergothioneine. Consequently, EGT@HA-nio exhibited superior in vivo efficacy, markedly attenuating UV-induced oxidative damage, restoring epidermal architecture, and preventing collagen degradation, thereby highlighting its potential as a versatile platform for penetration-limited therapies.
4.2.2.2
Ethosomes (ESs) and transfersomes (TRs)
Liposomal vesicle formulations can be broadly categorized into rigid vesicles, such as conventional LPs, and elastic vesicles, including ESs and TRs. A key limitation of traditional LPs is their restricted permeation through the SC, prompting the development of modified LPs with enhanced skin penetration. ESs, enriched with ethanol, function as reservoir systems enabling sustained drug delivery to target sites. As the addition of a high concentration of alcohol lowers the phase transition temperature of the liposomal membrane, the transdermal flux of ESs is better than that of conventional LPs, with a superior transdermal absorption effect. TRs, or deformable LPs, incorporate edge activators or surfactants, which, in combination with phospholipids, confer high membrane flexibility, allowing them to traverse pores up to five times smaller than their own diameter [158]. Yücel et al. compared rosmarinic acid (RA)–loaded ESs and conventional liposomes for transdermal anti-aging delivery [159], showing that optimized ESs achieved significantly higher transdermal flux and skin permeation. This improvement was attributed to ethanol-induced membrane fluidity and vesicle deformability, enabling deeper dermal deposition than conventional liposomes. Qin et al. developed ferulic acid (FA) ESs (FA-ES) co-modified with astragaloside Ⅳ and ceramide IIIB [160], whose structural and compositional optimization enhanced solubility and transdermal delivery. FA-ES reduced toxicity and improved skin hydration, repair, and antioxidant effects in zebrafish, while promoting absorption, lifespan, and stress resistance in C. elegans. Miatmokov et al. evaluated the impact of surfactant type on TRs loaded with amniotic mesenchymal stem cell metabolite products (AMSC-MP) for anti-aging therapy [161]. Incorporation of the anionic surfactant sodium cholate produced TRs with the smallest size (~144 nm), highly negative zeta potential, and superior skin penetration compared to cationic or nonionic counterparts. These compositional optimizations enhanced vesicle deformability and surface charge, enabling effective delivery of high-molecular-weight AMSC-MP into the deep dermis and significantly increasing collagen density in UV-aged skin. Harmita et al. demonstrated that TRs-based formulations significantly improved the stability and transdermal delivery of N-acetylcysteine compared with non-vesicular creams [162]. The incorporation of Tween-80 as an edge activator enhanced vesicle deformability, enabling more efficient and stable dermal antioxidant delivery. Malviya et al. developed novel vesicular carriers, TRs (SA-TRs) and ESs (SA-ETs), to enhance the dermal delivery of sinapic acid (SA) [163]. By incorporating edge activators or high ethanol content, these carriers exhibit superior elasticity and deformability, overcoming the limited permeation of conventional LPs. Comparative studies showed that both SA-TRs and SA-ETs markedly increased skin permeability, with SA-ETs achieving the highest flux (7.6 ± 0.6 µg cm−2 h−1) and a 4.2-fold enhancement, highlighting ESs as a promising transdermal antioxidant delivery platform. Collectively, these studies demonstrate that the strategic compositional and structural modifications of ESs and TRs, through ethanol enrichment or incorporation of edge activators, significantly enhance vesicle deformability, transdermal flux, and dermal deposition of bioactive compounds, overcoming the penetration limitations of conventional LPs and offering effective platforms for skin anti-aging therapy.
4.2.2.3
Solid lipid nanoparticles
Within a certain particle size range, solid lipid nanoparticles (SLNs) can inhibit the penetration of drugs into the skin while enhancing the penetration and accumulation of drugs in the upper skin layer to ensure local retention and action of drugs [164]. Kotb et al. investigated the anti-photoaging effects of hydro-distilled frankincense essential oil (FO) and its optimized SLNs formulation (FO-SLNs) in a UVB-irradiated rat model [165]. FO-SLNs markedly enhanced skin penetration, restored antioxidant capacity, suppressed inflammatory and aging markers, and improved skin histology compared with unformulated FO and vitamin A palmitate. These effects are attributed to the solid lipid nanoparticle design, in which Compritol® 888 ATO and Tween-80 improve solubility, stability, and sustained release of the hydrophobic oil, thereby overcoming the delivery limitations of conventional formulations and ensuring targeted dermal bioavailability. Aanisah et al. developed SLNs-based hydrogels to improve the solubility, skin penetration, and photoprotective efficacy of quercetin and luteolin from safflower extract [166]. The optimized formulation achieved high encapsulation efficiency, sustained release, enhanced dermal retention, and good biocompatibility, demonstrating its potential for preventing photoinduced skin aging. Overall, SLNs, through optimized lipid matrices, surfactants, and surface modifications, effectively enhance the solubility, stability, dermal penetration, and local retention of hydrophobic bioactive compounds. These studies highlight SLNs as versatile and safe carriers that overcome the limitations of conventional LPs and creams, offering promising strategies for anti-aging and photoprotective topical applications.
4.2.2.4
Nanostructured lipid carriers (NLCs)
NLCs, which effectively improve skin hydration, are considered superior DDSs. Accordingly, NLCs and enriched PFC® perfluorocarbon hydrogels have been developed as dual-method carriers for the topical skin delivery of natural products to improve the stability and skin permeability of bioactive agents. Eiras et al. developed a vitamin E-loaded NLCs hydrogel (HG-NLCVE) to enhance skin hydration and improve formulation consistency for topical application [167]. Characterization confirmed appropriate physicochemical properties, and in vitro assays demonstrated excellent biocompatibility and negligible irritancy. These results indicate that HG-NLCVE overcomes the low-viscosity limitation of conventional NLCs dispersions, supporting its potential for cosmetic and dermatological applications. Navabhatra et al. employed an artificial neural network (ANN) with K-fold cross-validation to optimize NLCs encapsulating a 5-O-caffeoylquinic acid–rich Cratoxylum formosum leaf extract (CFE-NLCs) [168]. The optimized formulation, with precisely balanced solid lipids, liquid lipids, and emulsifiers, exhibited enhanced physicochemical stability, a biphasic release profile for sustained delivery, and improved permeation into deeper epidermal layers compared with the free extract. These compositional and structural refinements effectively address the limitations of conventional plant extract formulations, including poor chemical stability, limited dermal absorption, and potential irritation. To explore how NLCs promote the skin penetration of highly lipophilic β-carotene across the SC barrier, Maretti et al. investigated the enhancement of β-carotene skin permeation using NLCs to overcome the limitations of conventional formulations [169]. They developed two nanosized NLCs (NLC1 and NLC2) and micro structured lipid carriers, evaluating morphology, loading efficiency, and in vivo penetration via tape stripping in human skin. NLC2 (~200 nm) significantly increased β-carotene delivery into deeper SC layers compared with the free compound and larger carriers, demonstrating the importance of optimized nanoscale design for transdermal transport of lipophilic actives. Collectively, these studies demonstrate that NLCs, through optimized composition, nanoscale architecture, and functional modifications, can significantly enhance the stability, skin penetration, and bioactivity of lipophilic compounds. Such advancements highlight the versatility and effectiveness of NLCs-based systems for topical and dermocosmetic applications, including hydration, photoprotection, and antioxidant delivery.
4.2.2.5
Cubosomes and hexosomes
Cubosomes and hexosomes are non-lamellar lipid nanoparticles with three-dimensional bicontinuous cubic or inverse hexagonal liquid crystalline structures. Cubosomes are formed by dispersing inverse bicontinuous cubic phases (Q2) into an aqueous medium, featuring a curved bicontinuous lipid bilayer (~3.5 nm) and two nonintersecting water channels with symmetries such as Im3m, Pn3m, or Ia3d [170]. Hydrophilic compounds localize near emulsifier polar heads or within water channels, hydrophobic compounds reside in the lipid bilayer, and amphiphilic compounds occupy the interface [171]. Hexosomes are derived from inverse hexagonal phases (H2), exhibiting densely packed, columnar water-filled cylindrical micelles with similar compartmentalization for hydrophilic, hydrophobic, and amphiphilic cargos [172]. These non-lamellar nanoparticles offer higher loading capacity, larger surface area, and enhanced chemical and mechanical stability compared to conventional micelles and vesicles [173]. Furthermore, cubosome–hexosome transitions can be modulated by temperature, ionic strength, or pH, enabling the design of stimuli-responsive nanocarriers for controlled and targeted delivery [174]. Fornasier et al. developed non-lamellar lipid nanoparticles, including cubosomes and hexosomes based on monoolein, stabilized with Span 80 and incorporating sodium taurocholate to enhance skin penetration for photodynamic therapy against metastatic melanoma [175]. By replacing conventional phospholipid bilayers with three-dimensional bicontinuous cubic (Pn3m) or inverse hexagonal (HII) phases and incorporating biocompatible surfactants, these nanoparticles exhibit optimized membrane curvature and deformability, overcoming the rigidity and limited drug-loading of traditional LPs. The formulations achieved high co-encapsulation efficiencies (~95%) for chlorin e6 and coenzyme Q10 and demonstrated potent photocytotoxicity against Me45 melanoma cells, particularly with cubosomes, highlighting their potential as structure-engineered nanocarriers for dermal oncotherapy. Despite their well-defined structural advantages and demonstrated efficacy in dermal drug delivery, the application of cubosomes and hexosomes in skin anti-aging remains limited, indicating substantial potential for further exploration and development in this field.
4.2.3
Polymer-based nanocarriers
4.2.3.1
Micelles
Pluronics comprise the copolymer poly(ethylene oxide) and poly(propylene oxide) that spontaneously form nanoscale structures above the critical micelle concentration. Pluronic nanogels as a delivery system can control drug release, enhance absorption, and maintain good biocompatibility, biodegradability, and nontoxicity [176]. Punicalagin is a natural product with antioxidant activity and poor stability. Garcia et al. prepared pluronic micelles encapsulating punicalagin and added Span80 to the formulation, which reduced their sol-gel transition temperature (P < 0.05), enhancing the skin permeability (~80–315 µg/cm2) of punicalagin [177]. Wichit et al. explored the ability of polymer micelles prepared from polyethylene glycol-conjugated phosphatidylethanolamine to stabilize all-trans retinoic acid (ATRA) under different storage conditions [178]. At a 750 Da molecular weight of polyethylene glycol, the micelle (PEG750–DPPE) could encapsulate ATRA up to 82.7% and significantly delay its oxidation, thereby improving its stability. These studies demonstrate that pluronic and polymer-based micellar systems effectively enhance the stability, skin permeability, and controlled release of labile bioactive compounds, highlighting their potential as safe and efficient topical delivery platforms.
4.2.3.2
Lecithin/chitosan nanoparticles (LCNs)
LCNs are nanoscale carriers formed by the self-assembly of negatively charged lecithin and positively charged chitosan, used to encapsulate bioactive compounds for enhanced stability, solubility, and transdermal delivery. Dong et al. developed LCNs via the self-assembly of positively charged chitosan and negatively charged lecithin, and loaded them with a baicalein-phospholipid complex (BPC) to enhance lipophilicity and encapsulation efficiency (84.5%) of the poorly soluble flavonoid [179]. The resulting BPC-LCNs exhibited a compact core–shell structure with high positive zeta potential (+33.0 mV), enabling sustained drug release, excellent colloidal stability, and significantly improved ex vivo and in vivo skin retention and permeation compared with conventional formulations. Badie et al. prepared and evaluated RES-self-assembled liquid crystalline nanoparticles [180]. Notably, the LCNs encapsulation for RES was ≥73.5%, and the release time was up to 24 h. Moreover, RES-self-assembled LCNs showed a 6-fold increase in transdermal penetration capacity and higher stability than those of the suspension. Overall, these studies demonstrate that such nanostructured lipid-based systems can achieve high encapsulation efficiency, sustained release, and markedly enhanced skin penetration and retention, offering promising platforms for topical delivery of lipophilic antioxidants.
4.2.3.3
Dendrimers
Dendrimers are highly branched, tree-like synthetic macromolecules with a well-defined 3D architecture, used as nanocarriers to improve the solubility, stability, and targeted delivery of bioactive compounds. Singh et al. investigated the use of poly(amidoamine) (PAMAM) dendrimers to encapsulate RES and develop effective topical cream and hydrogel formulations [181]. Molecular encapsulation, hydrogen bonding, and electrostatic interactions within the dendrimer's 3D architecture enhanced RES stability and significantly improved topical permeability. Preclinical studies in rat models demonstrated that dendrimer-RES formulations exhibited superior bioavailability, favorable pharmacokinetic profiles, and enhanced safety compared with free RES, highlighting their potential as effective anti-aging topical carriers. Belhadj et al. investigated poly-l-lysine dendrimers to enhance the stability and efficacy of vitamin C in dermocosmetic formulations [182]. Complexation with second-generation dendrimers (DGL2) increased vitamin C lipophilicity by over 300%, improved chemical stability in aqueous and emulsion systems under stress, and enhanced in vivo skin hydration by 66.2%. These findings demonstrate that poly-l-lysine dendrimers are effective, biocompatible nanocarriers that overcome the traditional limitations of vitamin C instability and poor skin penetration. Collectively, these findings highlight that dendrimer nanotechnology, whether PAMAM or poly-l-lysine, provides effective, biocompatible platforms to overcome the conventional limitations of antioxidant instability and poor skin penetration, supporting their potential in anti-aging and dermocosmetic applications.
4.2.4
Inorganic/hybrid nanocarriers
4.2.4.1
Metal nanoparticles
Metal nanoparticles are nanoscale particles composed of metallic elements, typically ranging from 1 nm to 100 nm, exhibiting unique physicochemical, optical, and catalytic properties distinct from their bulk counterparts. Wu et al. developed eco-friendly gallic acid-coated gold nanoparticles (GA-AuNPs) [183], which were characterized as spherical with an average diameter of ~40 nm and a negative zeta potential. GA-AuNPs effectively mitigated high glucose-induced OS and inhibited MMP-1-mediated type Ⅰ collagen degradation in dermal fibroblasts by suppressing ROS-dependent p38 MAPK/ERK and NF-κB signaling pathways. Lai et al. developed an innovative OS microenvironment (OSM)-responsive therapeutic strategy in which trivalent gold precursors are selectively biosynthesized in situ into functional fluorescent gold nanoclusters (FGNCs) within damaged skin [184]. By exploiting the pathological OSM as an intrinsic trigger, this approach circumvents key limitations of conventional nanocarriers, including nonspecific distribution and low delivery efficiency, enabling localized synthesis and activation of therapeutic agents. The biosynthesized FGNCs effectively attenuated inflammatory responses by suppressing the NF-κB/TNF-α signaling axis, highlighting a safe, precise, and facile therapeutic modality for OS–associated skin disorders.
Zhang et al. synthesized size-tunable platinum nanoparticles (PtNPs) using an eco-friendly Nymphaea tetragona flower extract method, yielding well-dispersed particles of ~4 and ~2 nm [185]. These PtNPs exhibited strong antioxidant activity, inhibited tyrosinase and UVB-induced melanin production in melanocytes, and promoted type Ⅰ collagen synthesis in dermal fibroblasts via activation of the TGF-β/Smad signaling pathway. Younis et al. synthesized magnesium oxide nanoparticles (RcNps) via a green method using Rosa floribunda charisma extract as both reducing and capping agent [186]. These RcNps demonstrated superior antioxidant, anti-aging, and antibiofilm activities compared to the crude extract. Their polyhedral morphology, nanoscale size (35.25–55.14 nm), and bioactive surface functionalization enhance skin penetration and enable targeted dermal delivery, addressing limitations of conventional nanocarriers.
4.2.4.2
Metal-organic frameworks (MOFs)
MOFs are crystalline porous materials composed of metal ions or clusters coordinated to organic ligands, forming highly ordered, tunable three-dimensional structures with large surface areas and versatile chemical functionalities. Li et al. designed and synthesized a novel exosome-loaded MOFs hydrogel composite to mitigate skin aging by targeting OS [187]. By integrating a luminescent Ba(II)-based MOFs with a three-dimensional porous hyaluronic acid/carboxymethyl chitosan hydrogel, the system combines the MOFs structural stability and catalytic potential with the polysaccharide matrix's biocompatibility and high drug-loading capacity. In vitro, the composite induced dose-dependent upregulation of SOD and reduction of MDA in human dermal fibroblasts, demonstrating its efficacy against oxidative damage and potential as an advanced transdermal delivery platform. Xiao et al. synthesized and characterized a series of zinc-based MOFs, identifying zeolitic imidazolate framework-8 (ZIF-8) as exhibiting the broadest and highest UVA/UVB diffuse reflectance due to its tailored bandgap achieved via precise ligand selection and framework engineering [188]. This MOFs-based sunscreen surrogate demonstrated superior safety and efficacy compared to conventional inorganic filters (TiO2 and ZnO), effectively minimizing ROS generation and reducing skin penetration through its hydrophobic surface and stable crystalline structure. In a pig skin model closely resembling human skin, ZIF-8 provided comparable or enhanced photoprotection, highlighting its potential as a tunable, next-generation platform for transparent high-performance UV filters. These studies illustrate the versatile potential of MOFs-based composites and frameworks as multifunctional platforms for skin therapeutics and photoprotection, combining structural tunability, biocompatibility, and targeted functional performance.
4.2.5
Nucleic acid self-assembled nanocarriers
Tetrahedral framework nucleic acids (tFNAs) have emerged as leading biological delivery vectors owing to their high programmability, minimal immunogenicity, excellent biocompatibility, and intrinsic biodegradability. tFNAs are highly effective biological carriers capable of traversing cellular barriers and modulating cellular behavior [189]; through rational static structural design, functional molecules can be incorporated into their framework to construct passive, cell or tissue-targeted DDSs [190]. Owing to their efficient cellular internalization, tFNAs function as versatile carriers for small molecules and oligonucleotides, conferring broad anti-inflammatory, antioxidant, antibacterial, and immunomodulatory activities with strong potential in barrier tissue repair and regeneration [191]. Li et al. engineered bioswitchable tetrahedral DNA nanostructures for microRNA delivery [192]. BDS-A anchors miRNAs via sticky-end hybridization, while BDS-B encapsulates them within the tetrahedral framework. These designs enhance cellular uptake, preserve nanostructural integrity, and enable RNase H-triggered release, overcoming poor stability, uncontrolled release, and limited tissue penetration of conventional carriers. Building on this, the BiRDS system encapsulates miRNA inhibitors, maintains a small size (~10 nm), and undergoes controlled 3D-to-2D transformation upon internalization. BiRDS demonstrates superior serum stability, efficient uptake, enhanced transdermal penetration, and effective anti-aging activity in skin models [193]. Yue et al. developed a binary anti-aging nanoplatform (siR-TDNbox) by engineering tetrahedral DNA nanostructures (TDNs) to encapsulate siRNA targeting Raptor, a key mechanistic target of rapamycin complex 1 (mTORC1) component [194]. Leveraging the intrinsic anti-inflammatory and antioxidant properties of the DNA framework, the platform not only protects and delivers siRNA but also mitigates age-associated inflammation, overcoming the single-function limitation of conventional carriers. siR-TDNbox exhibited synergistic effects, delaying cellular senescence, inhibiting the mTORC1 pathway, and extending lifespan, representing a novel strategy for combined senotherapeutic intervention. Li et al. developed a transdermal delivery platform by employing tFNAs as nanocarriers for α-arbutin, using a thermal loading method to achieve high encapsulation efficiency (76.85%) while preserving drug activity [195]. Leveraging the biocompatibility and skin-penetrating capability of the tetrahedral structure, this system overcomes the poor penetration and low loading limitations of conventional carriers. The tFNAs @α-Arb complex demonstrated exceptional efficacy, achieving 95.65% melanin inhibition in vivo through effective skin deposition and downregulation of key melanogenic genes, thereby enhancing the therapeutic potential of α-arbutin for hyperpigmentation treatment. In summary, these studies highlight tFNAs as highly versatile and multifunctional nucleic acid nanoplatforms capable of programmable cargo loading, efficient transdermal and cellular delivery, and synergistic regulation of aging and pigmentation-related pathways, underscoring their broad potential in advanced skin therapeutics.
5.
Conclusion
Skin aging arises from the intricate interplay of OS, inflammation, cellular senescence, mitochondrial dysfunction, and glycation, processes that are further intensified by environmental and lifestyle factors. Natural bioactive compounds offer a unique advantage in modulating these interconnected pathways due to their multi-target and pleiotropic biological activities. Accumulating evidence supports their capacity to attenuate photoaging, suppress inflammatory signaling, preserve ECM integrity, and restore redox homeostasis in aging skin. However, biological efficacy alone is insufficient for successful translation. The therapeutic and cosmetic potential of natural compounds is strongly influenced by their physicochemical properties and delivery efficiency. Advanced transdermal and nanocarrier-based delivery systems have emerged as critical enablers, capable of enhancing stability, penetration, bioavailability, and site-specific action (Fig. 5).
Figure 5
Figure 5.
Schematic overview of a transdermal delivery framework for natural anti-aging compounds. This figure illustrates a comprehensive strategy for addressing the key pharmaceutical challenges of natural bioactive ingredients, such as low solubility, chemical instability, and limited skin penetration. Natural compounds, including flavonoids, polyphenols, alkaloids, vitamins, and polysaccharides, are encapsulated within rationally designed nanocarriers, comprising lipid-based systems (e.g., liposomes, ESs, TRs, SLNs, NLCs, cubosomes), polymer-based carriers, inorganic/hybrid platforms (e.g., metal nanoparticles, MOFs), and nucleic acid–assembled structures (e.g., tFNAs). These nanocarriers are incorporated into topical formulations such as solutions, creams, ointments, and gels. Transdermal delivery efficiency is further enhanced by physical penetration-enhancement techniques, including MNs, electroporation, and iontophoresis, which transiently modulate the skin barrier to facilitate deeper and more targeted penetration of encapsulated actives. This integrated approach enables stabilized, targeted, and efficient transdermal delivery of natural anti-aging compounds, providing a versatile platform for advanced skin therapeutics. Created in https://BioRender.com.
Comparative analysis indicates that no single carrier platform is universally optimal; rather, delivery materials must be rationally matched to the physicochemical characteristics of the active compound and the underlying aging mechanism. Importantly, several challenges remain unresolved, including inconsistencies in experimental models, limited long-term safety data, insufficient understanding of skin-layer-specific distribution, and the translational gap between laboratory-scale formulations and scalable, regulatory-compliant products. Notably, despite extensive preclinical evidence, clinical studies specifically evaluating natural compounds and advanced delivery systems for skin anti-aging remain scarce, with most available data derived from in vitro or animal models. Future research should therefore prioritize mechanism-guided carrier design, stimuli-responsive and skin-microenvironment-adaptive systems, alongside standardized evaluation frameworks that integrate biological efficacy, delivery performance, and clinical endpoints. In conclusion, by critically integrating skin aging biology with delivery material innovation, this review moves beyond descriptive summarization and provides a forward-looking conceptual framework for the rational development of natural anti-skin-aging therapeutics. Such an integrated approach is essential to accelerate the translation of promising natural compounds from experimental research into clinically and commercially viable solutions for maintaining skin health and delaying aging.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
This work was financially supported by National Natural Science Foundation (No. 82304393), Beijing Natural Science Foundation (Nos. L242027 and L256067, China), CAMS Innovation Fund for Medical Sciences (No. 2025-I2M-XHXX-093), National Key R&D Program (No. 2023YFC2706100), and Beijing Nova Program (No. 20250484812).
P. Singh, R.K. Kesharwani, R.K. Keservani, Antioxidants and vitamins: Roles in cellular function and metabolism, Sustained Energy for Enhanced Human Functions and Activity, Elsevier, 2017, pp. 385–407.
P. Li, W. Zhang, S. Peng, et al., ACS Appl. Mater. Interfaces 17 (2025) 49227–49239. doi: 10.1021/acsami.5c10110
Figure 1
Compare aging skin and young skin at the tissue, internal structure, and cellular levels. (A) ECM, composed of collagen, elastin, and reticular fibers, provides structural support for the dermis. Aging or UV exposure leads to ECM degradation and reduced synthesis, causing wrinkles and sagging. (B) SC, the outermost epidermal layer, consists of anucleate keratinocytes and intercellular lipids, forming a "brick wall" barrier. Aging slows SC turnover, reduces moisture, and weakens barrier function, leading to dry and sensitive skin. (C) Aged skin exhibits wrinkles, thinning, flattened dermal-epidermal junctions, dermal papillae atrophy, decreased cell density, collagen breakdown, and elastin loss. (D) Young skin shows a smooth surface, tight dermal-epidermal junctions, dense cellularity, and well-organized collagen and elastin fibers. Created in https://BioRender.com.
Figure 2
(A) The solar spectrum consists of electromagnetic rays of varying wavelengths that penetrate the skin to different depths. (B) UVR damages the skin through multiple mechanisms. UVB induces DNA damage and promotes skin cancer. Keratinocytes release alpha‑melanocyte‑stimulating hormone (α-MSH) and endothelin‑1 (ET‑1), stimulating melanocytes to produce melanin, resulting in pigmentation and age spots. ROS generation triggers inflammatory responses by promoting the release of TNF-α, IL-1β, and IL-6 from keratinocytes, fibroblasts, and LCs, leading to sunburn and skin inflammation. ROS also upregulates MMPs (MMP-1, MMP-3, MMP-9), causing collagen degradation, wrinkles, and accelerated skin aging. Created in https://BioRender.com.
Figure 3
Sources of ROS and the signal pathways involved. Exogenous ROS include: UVR, air pollution, cigarette smoke, and heavy metal exposure. Endogenous ROS include the electron respiratory transport chain in mitochondria, NADPH oxidase, endoplasmic reticulum stress, and peroxisomes. The signal pathways involved in ROS mainly include: AP-1 signaling pathway, TGF-β/Smad signaling pathway, NF-κB signaling pathway, Nrf2/ARE signaling pathway, AGEs-RAGE signaling pathway, mTOR signaling pathway, p53/p21/p16 signaling pathway, etc. Solid arrows indicate activation, and dotted arrows indicate inhibition. Created in https://BioRender.com.
Figure 4
Schematic illustration of skin structure. (A) The skin consists of the epidermis, dermis, subcutaneous tissue, and appendages. (B) The epidermis includes, from top to bottom: SC, stratum granulosum, stratum spinosum, stratum basale, and the basement membrane. The SC is made up of anucleate corneocytes; The granulosum layer contains flattened keratinocytes; The spinosum includes keratinocytes and LCs; The basale consists of keratinocyte progenitor cells, melanocytes, and Merkel cells. (C) The SC has a brick-and-mortar structure, with dislocated corneocytes (15%–30%) embedded in a lipid matrix and connected by desmosomes. Created in https://BioRender.com.
Figure 5
Schematic overview of a transdermal delivery framework for natural anti-aging compounds. This figure illustrates a comprehensive strategy for addressing the key pharmaceutical challenges of natural bioactive ingredients, such as low solubility, chemical instability, and limited skin penetration. Natural compounds, including flavonoids, polyphenols, alkaloids, vitamins, and polysaccharides, are encapsulated within rationally designed nanocarriers, comprising lipid-based systems (e.g., liposomes, ESs, TRs, SLNs, NLCs, cubosomes), polymer-based carriers, inorganic/hybrid platforms (e.g., metal nanoparticles, MOFs), and nucleic acid–assembled structures (e.g., tFNAs). These nanocarriers are incorporated into topical formulations such as solutions, creams, ointments, and gels. Transdermal delivery efficiency is further enhanced by physical penetration-enhancement techniques, including MNs, electroporation, and iontophoresis, which transiently modulate the skin barrier to facilitate deeper and more targeted penetration of encapsulated actives. This integrated approach enables stabilized, targeted, and efficient transdermal delivery of natural anti-aging compounds, providing a versatile platform for advanced skin therapeutics. Created in https://BioRender.com.

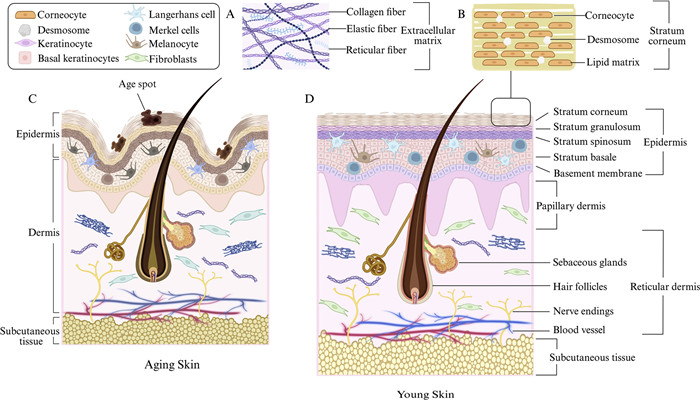
 DownLoad:
DownLoad:
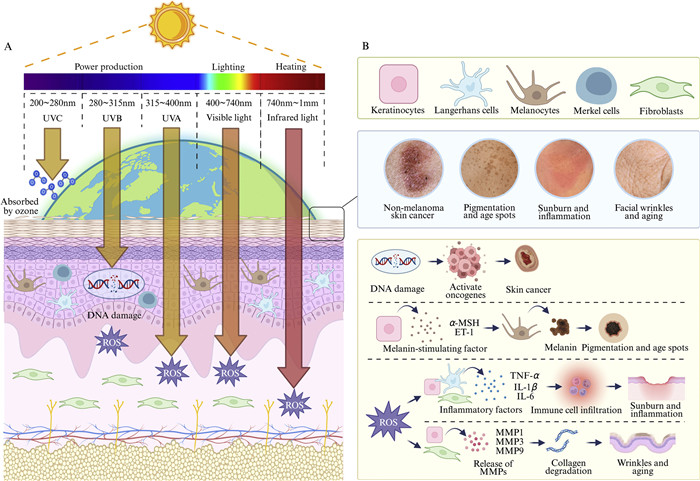
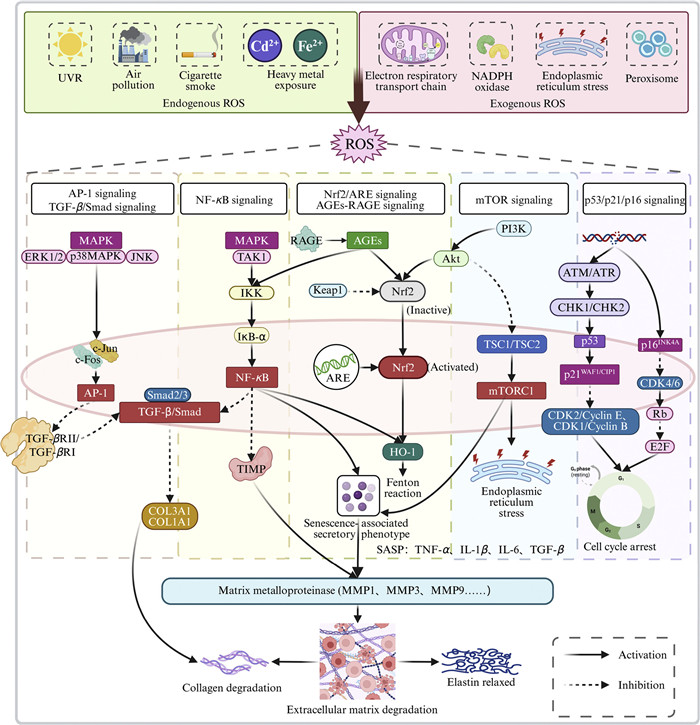
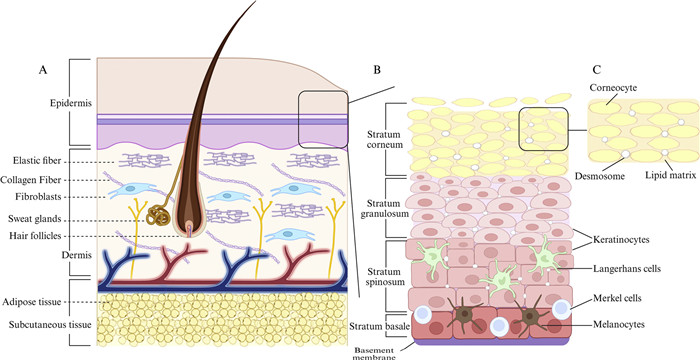
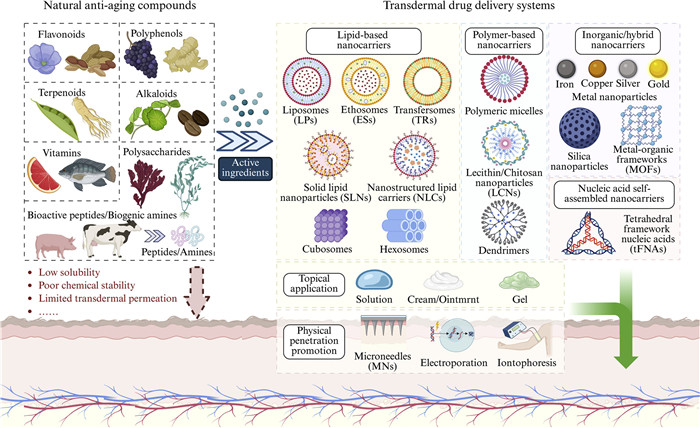

 下载:
下载:






 下载:
下载:
