Dual efficacy of extracellular polymeric substances in microalgae-fungi consortia system: Advances and challenges in wastewater treatment and biomass recovery mechanisms
Citation:
Hanglong Ma, Zhihong Yin, Qi Wang, Yuhong Li, Lei Tian, Jing Yang, Liandong Zhu, Shaoqi Zhou. Dual efficacy of extracellular polymeric substances in microalgae-fungi consortia system: Advances and challenges in wastewater treatment and biomass recovery mechanisms[J]. Chinese Chemical Letters,
2026, 37(7): 112174.
doi:
10.1016/j.cclet.2025.112174
Dual efficacy of extracellular polymeric substances in microalgae-fungi consortia system: Advances and challenges in wastewater treatment and biomass recovery mechanisms
English
Dual efficacy of extracellular polymeric substances in microalgae-fungi consortia system: Advances and challenges in wastewater treatment and biomass recovery mechanisms
College of Resources and Environmental Engineering, Key Laboratory of Karst Georesources and Environment, Ministry of Education, Guizhou University, Guiyang 550025, China
b.
Guizhou Karst Environmental Ecosystems Observation and Research Station, Ministry of Education, Guiyang 550025, China
c.
School of Resource and Environmental Sciences, Hubei Key Laboratory of Biomass-Resources Chemistry and Environmental Biotechnology, Wuhan University, Wuhan 430079, China
Received Date:
12 July 2025 Accepted Date:
25 November 2025 Revised Date:
03 October 2025 Available Online:
15 July 2026
Abstract:
As a new wastewater treatment technology, the microalgae-fungi consortia system (MFCS) has attracted much attention because of its high efficiency, environmental protection and energy saving features. Extracellular polymeric substances (EPS) play a key role in MFCS treatment of wastewater and microalgae harvesting. In this review, the definition and formation process of MFCS, thoroughly examines the composition, properties, and influencing factors of EPS, as well as systematically explains the mechanism of action of EPS in wastewater treatment and microalgae harvesting. EPS surface functional groups have negative charge, adsorption, flocculation, and hydrophilicity/hydrophobicity properties. Microalgae and fungi form MFCS through EPS electrostatic attraction and protein bridging. EPS achieves adsorption, aggregation, and degradation of pollutants through targeted binding, chelation, coordination mechanisms, hydrophobic interactions, and complexation degradation mechanisms via functional groups. EPS achieves microalgae harvesting through electrostatic neutralization, surface protein interaction, and polysaccharide adhesion. However, current research still faces the following challenges, the selection of algal strains in the formation process of MFCS is still unclear. The mechanism of EPS removal of new pollutants and internal signaling pathways are still unclear. FSH takes too long, while FPH requires higher costs during harvesting. In the future, MFCS should be explored in combination with actual engineering applications, and different combinations of algae species and strains should be explored. Using genetic engineering to explore the synergistic mechanism of microalgae and fungi EPS in pollutant treatment. Improve the process conditions for harvesting microalgae using fungi, optimizing the two methods of microalgae harvesting, and conduct experiments on the efficient harvesting of microalgae. This review systematically explored the mechanism of action of EPS in MFCS, which is important for wastewater treatment and microalgal biomass harvesting.
The rapid development of the economy and society has caused many environmental problems, among which wastewater treatment has become increasingly serious. Wastewater pollutants are complex, they are rich in the conventional nutrients C, N, and P, but mixed with many heavy metals and emerging pollutants. Traditional physical, chemical, and biological treatment methods are faced with limited capacities, equipment investments, and high operating costs [1,2]. Microalgae biotechnology for wastewater treatment is a means of reducing pollution and carbon while efficiently using biomass [3]. However, the use of single microalgae for wastewater treatment faces the disadvantages of low performance when treating high concentrations of pollutants, with consequently high harvesting costs [2,4]. Enhancing the performance and harvesting efficiency of microalgae biotechnology for wastewater treatment is currently a major challenge.
By constructing a microalgae-fungi consortia system (MFCS), the two mutually beneficial symbionts form a stable co-culture, which can greatly improve the pollutant removal efficiency, enhance system stability, and increase the resource recovery efficiency [2,5]. Microalgae and filamentous fungi interact with each other through EPS on their surfaces to form a stable and mutually beneficial symbiotic system with a stable structure that maximizes the wastewater treatment capacity of the microalgae and fungi [2]. For example, most filamentous fungi demonstrate good adsorption of heavy metal ions, and the presence of fungi improves the tolerance of an MFCS to heavy metals [2]. One study showed that the adsorption efficiency of an MFCS for Cd(Ⅱ) was 98.89%, with a maximum adsorption capacity of 37.3 mg/g, which was higher than that of a single microalgal-fungal system [6]. MFCS also provide greater removal of the conventional nutrients C, N, and P. A study by Yang et al. reported that the chemical oxygen demand (COD) removal efficiency of an MFCS was significantly greater than that of a single Chlorella system, with a removal efficiency of 70.68%. The TN removal efficiency of the MFCS was significantly higher than that of the single system, with the MFCS removing 67.09% of TN, compared to 44.39% and 18.20% for Chlorella and Trichoderma monoculture systems, respectively [7]. Mi et al. used a fungal-microalgae complex (FMC) composed of Cordyceps militaris and Navicula seminulum to effectively remove various antibiotics from aquaculture wastewater. The treatment efficiency of triclosan (TCS) was 82.6%, while the treatment efficiency of by individual Navicula seminulum and Cordyceps militaris systems were 67.4% and 56.6%, respectively. The FMC group had a processing efficiency of 77% for lithium nitrite (LIN), which was also much higher than that of the individual processing groups. Microalgae remove antibiotics by adsorption, accumulation, and biodegradation, while fungi increase the removal efficiency by producing laccase. Furthermore, a variety of functional groups in the FMC alter the structure of antibiotics by hydroxylation, glycosylation, demethylation, and hydrolysis, rendering the degradation products more volatile or removing them from the aqueous phase [5]. In addition, in a symbiotic system, filamentous fungi act as efficient flocculants for microalgae harvesting, which is a low cost, highly efficient and simple harvesting method that causes little damage to microalgae, and has wide adaptability compared with other harvesting methods such as flotation, sedimentation, filtration, and flocculation. The tryptophan and humic acid on the surface of filamentous fungi can directly capture and immobilize microalgae on the hyphae and between the hyphae through an EPS protein bridging mechanism, which captures the microalgae and creates a globule without damaging the microalgal cells [2,8,9]. Luo et al. used Pleurotus ostreatus HEI-8 to harvest Chlorella sp. NCU C01, achieving a harvest efficiency of 64.86% after 2.5 h [10]. Similarly, Arghya Bhattacharya et al. achieved a 99% harvesting efficiency of Chlorella pyrenoidosa in 4 h using Aspergillus Fumigatus [11]. Microalgae and fungi interact with each other through EPS to form MFCS, which exert powerful ability in wastewater treatment. Furthermore, the fungi can be used as a natural flocculant to harvest microalgae with a low energy consumption.
A statistical analysis of the literature showed that the research on MFCS treatment of wastewater has been increasing yearly or stable (Fig. 1a). From the perspective of generating MFCS for wastewater treatment EPS serves as a bridge connecting microalgae and fungi, and are therefore key to forming an MFCS. It EPS also enhance the wastewater treatment capacity and assist fungi in harvesting microalgae (Fig. 1b). Keyword clustering and co-occurrence analysis was performed on the MFCS wastewater treatment process literature results using CiteSpace scientometric software. The high Q value of 0.8547 in the keyword clustering indicated that EPS had a significant structure in the keyword clustering of wastewater treatment and flocculation harvesting. A high S value of 0.8993 indicated that the keyword clustering map had high persuasiveness (Fig. 1c). The results indicated that there is a high level of attention to EPS in MFCS within the fields of wastewater treatment and flocculation harvesting (Fig. 1d). In summary, EPS have a crucial role in MFCS used in wastewater treatment and flocculation recovery of microalgae biomass. In recent years, an increasing number of scientists have expressed interest in the field of MFCS wastewater treatment. However, current research focuses on the impact of environmental factors on the secretion content of EPS in the formation of MFCS. Pollutant removal focuses on the macro-level pathways of biological adsorption, accumulation, and degradation. Focusing on microalgae harvesting, research on the charge neutralization mechanism between microalgae and fungi [12,13]. Tian et al. systematically explained macro-level interactions such as gas exchange, metabolite transfer, and EPS secretion, ultimately forming the MFCS [12]. Wang et al. described in detail the regulation of MFCS system stability by environmental factors such as temperature and pH [2]. The removal of pollutants mainly focuses on the bioadsorption, accumulation and degradation mechanisms of MFCS [2,12]. In terms of microalgae harvesting, Alam et al. pointed out that differences in surface charges between fungi and microalgae drive flocculation [14]. Singh and Nazari et al. also demonstrated that this biological flocculation results from a charge neutralization mechanism [13,15]. In summary, current research has yet to establish a holistic mechanistic framework for MFCS with EPS as the central hub, particularly lacking an integrated analysis of EPS mediated microscopic interactions, pollutant transformation pathways, and environmental response mechanisms across the processes of MFCS formation, pollutant removal, and microalgae harvesting.
Figure 1
Figure 1.
(a) WOS literature study on microalgae, fungi, EPS, and wastewater treatment in the last five years. (b) MFCS formation, wastewater treatment, microalgae harvesting process. (c) Keyword clustering map, and (d) keyword co-occurrence map. The figure is originally created by the authors using Microsoft PowerPoint and Citespace.
Therefore, the main contents of this review include the following. (ⅰ) Starting from the formation of MFCS the structural characteristics of EPS and the factors affecting their production are explored; (ⅱ) The mechanisms of action of EPS on different pollutants treated by MFCS was analyzed, including those of conventional pollutants, heavy metals and new pollutants; (ⅲ) The mechanisms of EPS in the process of harvesting microalgae from MFCS and the influence of external environmental factors on the harvesting process are summarized; (ⅳ) The challenges of EPS in for wastewater treatment and the harvesting process of MFCS are analyzed trends and future developments are predicted. The purpose of this study is to provide a useful reference and guidance for applying EPS in wastewater treatment and harvesting processes with MFCS.
2.
Definition and formation of MFCS
2.1
Definition of MFCS
Microalgae and fungi are fixed through electrostatic adsorption of EPS, and the bidirectional circulation of substances forms MFCS. In an MFCS, microalgae are immobilized on the surface of fungal mycelial particles through the electrostatic attraction of secreted EPS, while the fungi synthesizes amino acids through N cycling and CO2O2 cycling to provide the necessary metabolic support for microalgae photosynthesis [16]. MFCS have been shown to enhance biomass yield, and have high potential for applications in environmental remediation and resource recovery. Mi et al. explored an MFCS of Cordyceps militaris and Navicula seminulum for the sustained treatment of aquaculture water, and found that the MFCS had a higher biomass content than to a single microalgal or fungal system [5]. MFCS have also shown advantages in the elimination of heavy metal contaminants, antibiotics, and pathogenic Vibrio bacteria [5].
2.2
Formation of MFCS
In the process of forming an MFCS, fungal mycelia are used to harvest microalgae. There are two different cultivation methods, Fungal Spore-assisted Harvesting (FSH) and Fungal Pellet-assisted Harvesting (FPH) (Fig. 2). An FSH system is a co-culture of filamentous fungal spores with microalgae. The spores gradually develop into filamentous fungi in the medium, while the microalgae continue to grow and multiply. After a period of time the microalgae become entrapped on the surface of the fungi or entangled and wrapped inside the mycelial sphere by the hyphae, eventually forming an MFCS [8]. In FPH, conidia are directly co-cultured with logarithmically growing microalgae [2,8]. First, fungal conidia form mycelial deposits of about 2–3 cm in liquid medium. Then, the mycelia are co-cultured with microalgae in logarithmic growth. After co-culturing for a certain period of time under specific conditions, the MFCS is obtained by filtering and isolation [2]. Microalgae and fungi interact with each other through bridging during the formation of an MFCS. The extracellular proteins in EPS are critical for the MFCS bridging process. The interactions among functional groups (e.g., amino, amide, phosphate, hydroxyl, and aldehyde groups) the microalgae and fungi increase the rate of photosynthesis and the content of photosynthetic pigments in microalgae. This improves the efficiency of photosynthesis in the microalgae, which require significant energy metabolism and material exchange [16]. The synthesis of amino acids in an MFCS facilitates an increase in extracellular protein production, and the cycling of CO2 and O2 provides gas exchange in the MFCS, which satisfies the metabolic activity of the microalgae and fungi as a mutually beneficial symbiotic system. For example, Amrit Lal co-cultivated Aspergillus sp. with Chlorella sp. MJ11/11 to form an MFCS, which resulted in higher yields of biofuels [17]. Zhang et al. used Chlorella vulgaris with Aspergillus niger Ted S-OSU through from an FSH culture to form an MFCS [18].
Figure 2
Figure 2.
Formation process of MFCS by FPH and FSH. The figure is originally created by the authors using Microsoft PowerPoint.
3.
Structure-defining compositional properties of EPS
3.1
Definition and classification of EPS
EPS are high molecular weight polymers secreted by microorganisms that play a key role in the formation and function of microbial aggregates. The concept of EPS was first proposed by Flemming et al., who described them as "extracellular polysaccharides" with the main emphasis on their polysaccharide components [19]. Subsequently Wingender et al. used EPS as a collective term to cover a wide range of macromolecules in microbial aggregates, including polysaccharides (PS), proteins (PN), nucleic acids, lipids and other polymers [20]. Nielsen et al. further expanded the definition of EPS by stating that those polymers that are located outside the cell wall, although not directly attached to the surface of the cell membrane, are also categorized as EPS [21]. Geesey, however, regarded EPS as functional substances secreted by microorganisms during the process of microbial aggregation formation [22]. Salama et al. further refined the definition of EPS by considering them as organic polymers produced by microorganisms that are capable of binding to other particles in biofilm systems and forming adhesions to their substrates [23]. Taken together, EPS is widely recognized as a polymer composed of high molecular weight secretions, cell lysates, macromolecular hydrolysis products secreted by microorganisms, and adsorbed organic matter. In MFCS, EPS plays a crucial role in maintaining system stability and function. Microalgae secrete EPS during growth, that are mainly composed of PS and PN. These EPS are attached to the surface of the microalgal cell wall, which helps to maintain cell morphology, and are involved in extracellular enzyme secretion, defense under stressful environments, and intercellular interactions. EPS secreted by fungi consists mainly of PS and are usually attached to the surface of the mycelium or surrounded by the mycelium, thus providing attachment sites for microalgae that promote MFCS formation [24].
EPS contain three main types of components (Fig. 3), namely soluble extracellular polymeric substances (S-EPS), loosely bound extracellular polymeric substances (LB-EPS) and tightly bound extracellular polymeric substances (TB-EPS) (Fig. 3). S-EPS is freely soluble in the fluid surrounding the cell, whereas B-EPS is tightly bound to the cell surface, although there are no specific differences between the two in terms of composition. Bound extracellular polymeric substances (B-EPS) can be further divided into LB-EPS and TB-EPS, which differ in their protein and polysaccharide content and produce structural and functional differences. LB-EPS is located in the outer layer of TB-EPS, and have a looser structure and a main function of helping the cell to adsorb nutrients. TB-EPS is tightly bound to the outer layer of the cell and form the external structure of the cell. This categorization reflects the differences in the chemical composition of EPS, and reveals their functional diversity in microbial aggregates [25].
Figure 3
Figure 3.
Structural definition, compositional features, and characteristics of EPS. The figure is originally created by the authors using Microsoft PowerPoint.
EPS is a complex multi-component mixture with a variety of organic components (e.g., PN, PS, humic acids, nucleic acids, glyoxylates, and lipids) and inorganic components (Fig. 3). Of these, PN and PS dominate, usually accounting for 70%−80% of the total EPS composition [26]. However, the composition of EPS is not constant, but instead varies significantly depending on the species of organisms, stage of growth, and the culture environment. For example, Xiao et al. systematically analyzed the EPS of certain algal genera in the phyla Chlorophyceae [27]. It has been shown that the PS of Chlorella phylum were mainly composed of hexoses, pentoses, glyoxalates, methyl pentoses, methyl hexoses and sulfates. In contrast, the EPS of the diatom phylum, contained fucose, contains fucose, rhamnose, mannose, glucose, xylose, arabinose, galactose and glucuronic acid. The EPS of the red algal phylum were alternately linked with monosaccharides such as glucose, xylose, galactose and glucuronic acid [27]. In addition, the PN in the EPS was mainly composed of aromatic proteins, the types and contents of which varied depending on the microbial species and environmental conditions. For example, Wang et al. analyzed the EPS of a unicellular microalgae (Ankistrodesmus falcatus var. acicularis) by three-dimensional fluorescence spectroscopy (3D-EEM), and identified several components, including aromatic proteins (Ⅰ and Ⅱ), xanthohumic acid-like organisms (Ⅲ), soluble microbial by-products (Ⅳ), and humic acid-like organisms (Ⅴ). Of these, humic acid-like protein was the only one detected in LB-EPS [3]. Liu et al. further investigated the EPS of microalgal biofilms using 3D-EEM technology and found that the protein composition mainly consisted of, polycarboxylate-type humic acids, and tyrosine and tryptophan amino acids [28]. These studies showed that the composition and content of EPS were not constant and were linked to parameters such as the sample type, collection method, culture environment and culture time.
PS and PN are the core components of EPS, and their contents and ratios directly affect the functional properties of EPS. For example, Wang et al. found that the high PS content in S-EPS was mainly responsible for the cohesion and adhesion of suspended cells in a culture medium. The PN is extensively involved in the formation of the polysaccharide matrix, which is closely related to the formation and stability of TB-EPS. The level of PN content determines whether microalgae can form integrity aggregates rapidly [3]. Melo et al. further showed that the content of TB-EPS correlated with the microbial concentration and was significantly affected by the total sugar and protein content [29]. Furthermore, Jin et al. similarly mentioned that the higher the microbial activity was, the more the EPS secreted more PS, and the content of PN and PS determined the flocculation capacity of the microalgae [30]. These studies revealed a close relationship between the EPS components and their functions, and provided an important theoretical basis for optimizing the application of MFCS.
3.3
Characterization of EPS
EPS exhibit a variety of unique physicochemical properties like negative surface charge, adsorption, flocculation, and hydrophilicity, rendering it important in environmental remediation and microbial aggregate formation. The negative charge of EPS mainly comes from functional groups in its polysaccharide fractions, such as carboxyl (-COOH) and phosphodiester (-O-P(O)(OH)-O-) groups [2]. Under suitable pH conditions, these groups ionize by releasing H+, giving EPS a negative charge (Fig. 3). Jesus et al. (2019) found that glucuronic acid, pyruvic acid, acyl groups, or amino acid carboxyl groups in the protein fraction also contribute to this charge [31]. This property affects EPS stability and enables interactions with environmental cations like metal ions [31]. EPS’s adsorptive capacity stems from surface functional groups (-COOH, -NH2, -OH, etc.) that form stable complexes with metal ions. Negatively charged coordination groups bind to heavy metals, while EPS’s porous structure and large specific surface area enhance adsorption [2]. For instance, extracellular proteins (PN) and polysaccharides (PS) of Chlamydomonas reinhardtii specifically bind zinc and copper, respectively. FTIR and XPS analyses showed that under 5 mg/L zinc exposure, the peak intensities of protein-related functional groups (e.g., C=O in amide Ⅰ, C-N/N-H in amide Ⅱ) on the microalgae cell surface decreased and shifted. For copper, functional group shifts were mainly associated with carbohydrates (e.g., C-H, COOH) [32]. indicating EPS adsorption is closely linked to its chemical composition and structure.
The flocculating properties of EPS stem from their bilayer structure and rich functional groups (e.g., -COOH, -NH, -OH, -CO), which enable them to form a barrier for microorganisms to protect against the effects of changes in water quality and quantity and exposure to toxic substances. Furthermore, the large surface area of EPS and the aggregation of functional groups promote dispersed particles to aggregate and form larger flocs, thus enhancing the stability of the system. The flocculation properties of EPS have important applications in wastewater treatment and microalgae collection processes. The hydrophilicity of EPS depends on the different functional groups of PN and carbohydrates in its EPS, as the hydroxyl groups (-OH) in EPS is able to form hydrogen bonds with water molecules, rendering EPS with some hydrophilicity [2]. However, the compact structure and strong intermolecular forces of the PN in EPS are capable of locking up the bound water, thus exhibiting hydrophobic properties [33]. For example, in their analysis of EPS PS and PN in the genus Limnothrix, Han et al. reported that PN were mainly composed of hydrophobic amino acids, while carbohydrates contained a higher proportion of hydrophilic groups [34]. This balance between hydrophilicity and hydrophobicity enables EPS to perform their functions under different environmental conditions.
4.
Factors affecting the production of EPS
4.1
Biological factors
The production of EPS is influenced by a variety of biological factors, primarily the species of the organisms, their growth stages, and the interactions among organisms (Fig. 4). There are significant differences in the composition of EPS produced by different microalgal species. For example, the PS of common green microalgae such as Chlorella sp., D. salina, Ankistrodesmus angustus, and B. braunii do not have the same composition as those of red algae. In addition, glycoproteins are more common in the EPS of green algae. However, many non-covalently bound proteins are more common in red algal cells, for example the major amino acids of the extracellular PN of Chlorella vulgaris include aspartic acid (asp), glycine (gly), glutamic acid (glu), alanine (ala), and serine (ser) [27]. The EPS of fungi consists mainly of PS attached to the surface of the fungal mycelium or surrounding the mycelium. In addition to PS, it also includes substances such as PN, fats, oxalic acid, and melanin. The content of EPS fractions produced by different species of fungi were shown to vary significantly [24]. For example, Gutiérrez et al. studied six species of fungi, including Pleurotus eryngii and the Pleurotus sajorcaju, and found that their EPS was mainly based on glucan dominant chained structural substances, with sugars accounting for 75% of the total mass with a low fraction of protein content [35]. Xu et al. found that the EPS of the fine-legged Penicillium sp. sugars accounted for 70.3%−85.7% of the total mass and PN accounted for 14.3%−29.7%. In addition, Leung et al. (2009) reported that sugars in the EPS of Cordyceps sinensis accounted for 54.9%−70.1% of the total EPS mass and PN accounted for 26.2%−29.3% [36]. Therefore, the type of microorganism significantly affects the composition of the EPS.
Figure 4
Figure 4.
Factors influencing EPS production in MFCS. The figure is originally created by the authors using Microsoft PowerPoint.
The growth stage of microorganisms has a significant effect on their EPS production. For example, the EPS content of microalgae in the logarithmic growth stage increased faster than in other stages, and the EPS content decreased slightly and stabilized when it reached the growth equilibrium stage. Cristiane Santos de Jesus et al. found that EPS production was closely related to the kinetics of cell growth [31]. Similarly, Zhang et al. studied the self-flocculation of microalgae of S. quadricauda by collecting EPS after the microalgae had cultured to a stable stage [18]. Li et al. and Xie et al. succeeded in increasing the number of EPS-binding sites by co-cultivating microalgae at the logarithmic growth stage with filamentous fungi to achieve efficient flocculation [37,38]. In addition, microbial interactions can also significantly affect EPS production. Jin et al. explored the simultaneous nitrification and denitrification processes of microalgae and activated sludge and found that microalgae and activated sludge worked together to significantly increase the production of EPS [30]. Similarly, Bertsch et al. conducted monoculture and co-culture experiments by comparing Lactobacillus rhamnosus strains (ATCC 9595, R0011, and RW-9595 M) with Saccharomyces cerevisiae, and found that RW-9595 M had higher EPS production under co-culture conditions [39]. In summary, the microbial species, growth stage, and inter-organism interactions all have a significant impact on the production of EPS. Typically, microalgae and bacteria produce the largest amount of EPS during the stabilization period, and the specific composition of the EPS is closely related to the microbial species. Importantly, the interactions between organisms can further promote the production of EPS, which therefore also provides a potential pathway to regulate the production of EPS.
4.2
Abiotic factors
4.2.1
Light conditions
The light condition is one of the important abiotic factors affecting the production of microalgae EPS, which mainly includes light quality, light intensity and wavelength (Fig. 4).
(ⅰ) Different light qualities have significant effects on the EPS production and composition of microalgae. Some studies have shown that the EPS production of Chlorella vulgaris under green light and blue light was significantly higher than that under white light. For example, Zhang et al. found that after four growth cycles culturing Chlorella vulgaris, the highest EPS yield of 20.48 mg/g was achieved under green light and the EPS protein and polysaccharide contents reached 126.26 mg/g and 214.72 mg/g, respectively. Cells cultured under blue and green light secreted higher amounts of PN and PS in their EPS compared with secretion under white light. A 3D-EEM analysis showed that EPS, which are mainly composed of soluble protein-like compounds and humic acid-like compounds, had the lowest accumulation of these two type of compounds under white light and the highest under green light [40]. In addition, light quality determines the relative abundance of specific functional groups in EPS. For example, blue and green light irradiation was shown to promote the production of O=C-OH and O=C-OR functional groups, which are associated with PS, carboxylates, and glucuronides [40]. Thus, light quality affects the yield of EPS, and has a regulatory effect on their chemical composition.
(ⅱ) Light intensity is an important determinant of EPS synthesis by photosynthetic autotrophic microalgae. In general, a higher light intensity can significantly increase EPS production before reaching the light saturation point. However, there are differences in the adaptability of different microalgal species to light intensity [41]. For example, Porphyridium sordidum had the highest EPS production at a light intensity of 80 µmol m-2 s-1, whereas Chlorella EPS production decreased with increasing light intensity (50, 120, 180 µmol m-2 s-1). Similarly, Cyanobacterium aponinum, Anabena spp, and Nostoc sp, showed different patterns of change in EPS production under specific light intensities. Light intensity affects both the total yield of EPS and their composition. Increased light intensity was found to increase the protein content of EPS [41]. It was reported that, the effect of glycine on the yield of bound EPS was regulated by light intensity. For example, Shen et al. found that at 250 µmol m-2 s-1 light intensity (mixing time of 3 days, glycine concentration of 0.1 g/L), the B-EPS yield reached 47.3 mg/g, while at a 125 µmol m-2 s-1 light intensity (mixing time 3 days, glycine concentration 1 g/L), the B-EPS yield was 44.2 mg/g [42]. Light intensity regulates the amount and composition of EPS by affecting the charge distribution of extracellular PS. For example, as cyanobacterial cells were exposed to increasingly stronger light, the amount of polysaccharides (RPSs), podoplanar polysaccharide (CPS), and total EPS increases; whereas the weaker the light, the higher the ratio of podoplanar polysaccharide (CPS) to extracellular polymer (EPS), and the lower the ratio of released polysaccharide (RPS) to EPS [43].
(ⅲ) A mix of wavelengths also has a significant effect on EPS productivity. For example, EPS production increased with time when flagellates were cultured under different light conditions, and there were differences in EPS production when cells were cultured under different mixed wavelengths of light. The EPS production of cells cultured under light at a red/blue ratio of 15:7 and a red/blue/green ratio of 12:5:5 was 1.71 and 1.68 times higher than that of cells cultured under white light, respectively [44]. Correspondingly, the wavelength-shifting method also had a significant effect on EPS yield. For example, under (9 days of exposure to white light followed by 9 days of exposure to red/blue/green light at a ratio of 12:5:5, the EPS content reached its highest value, which increased by 85.61% compared to the control, suggesting that fine tuning the light conditions could increase the total EPS yield [44]. In summary, light conditions (including light quality, light intensity and wavelength mix) have important effects on the production and composition of microalgal EPS. Green and blue light have been found to significantly increase EPS production and its protein and polysaccharide contents, while light intensity regulated EPS synthesis by affecting photosynthesis and the charge distribution of EPS components. Also, the use of mixed wavelengths and wavelength shift methods further indicate that EPS productively may be effectively improved by optimizing light conditions.
4.2.2
Temperature
Temperature is a key abiotic factor in the regulation of EPS synthesis in microalgae, and strongly species-specific, as evidenced by the differential regulation of EPS production, composition and synthesis pathway (Fig. 4). The EPS synthesis response to temperature in different microalgae is diverse and can be classified into positive temperature-responsive, optimal temperature-matched and extreme temperature-induced. In some positive temperature responsive microalgae. EPS production increased significantly with increasing temperature, For example, Nahla Mezhoud et al. found that the highest EPS production at the highest temperature (40 ℃) compared to the lowest light intensity (20 µmol m-2 s-1) in their study of a thermophilic Graesislla strain. Elevated temperature (20 ℃ to 40 ℃) was shown to be a major driver of EPS synthesis, indicating that high temperature may promote EPS secretion by activating protective metabolic mechanisms [45]. Some microalgae are optimal temperature-matched, with optimal temperature profiles for EPS secretion that overlap with their optimal growth temperatures. For example, B. braunii had its highest EPS production rate at 25–30 ℃, which was consistent with the optimal temperature range for algal growth [41]; Other microalgae are extreme temperature-inducing types, with the potential to produce more EPS at extreme temperatures due to the presence of enzymes that have different optimal temperatures for microbial growth during EPS production. For example, the EPS production of Anabaena sp. ATCC 33047 at 40–45 ℃ was significantly higher than at 30–35 ℃ [46]. Temperature affects the mechanism of action of EPS, probably on intracellular enzymes. During EPS production, certain enzymes are active, and unlike the temperature at which microorganisms optimally grow, extreme temperatures stimulate these intracellular enzymes to produce EPS to cope with extreme environments. Temperature regulation of EPS in microalgae is characterized by a complex dosage effect and species specificity. In industrial production, culture conditions should be optimized according to the temperature response of the target algal species. For positive temperature-responsive species, a gradient warming strategy can be used, whereas for extreme temperature-inducing species, a balance between EPS production and cell survival is required.
4.2.3
pH
pH is a key environmental factor regulating the synthesis and structure of EPS, with an influence throughout the dynamic processes that affect EPS morphology and conformation, polysaccharide secretion efficiency, and protein secondary structure (Fig. 4). The chain structure of EPS exhibits significant conformational changes (such as stretching and curling) under different pH conditions, which are mainly regulated by electrostatic interactions and the state of dissociation of functional groups. For example, Wang et al. confirmed that the slight stretching of the chain structure of EPS under alkaline conditions (pH 7.0–10.5) facilitates diffusion of the extracellular matrix and exposure of functional sites by exploring EPS secretion by Bacillus megaterium TF10. Under acidic conditions (pH 1.3–7.0) protonation enhanced intramolecular hydrogen bond formation, which caused the EPS chain structure to undergo random curling and incomplete chain extension. Greater disruption of carboxyl groups and amide groups; and a disordered, randomly curled conformation were present at a higher pH [47]. PS are preferentially secreted under weakly alkaline conditions. Most algae produce higher levels of EPS under alkaline conditions. For example Scytonema javanicum PS production was significantly higher at pH 7.5 than under acidic conditions [46]. The highest EPS production reported in one study was observed in Tetradesmus obliquus and Anabaena flos-aquae at pH 8.0 and 9.0, respectively [46]. Structural changes in proteins vary according to pH conditions. A pH change affects the secondary structure of EPS proteins by reconfiguring the charge distribution and disrupting hydrogen bond networks. For example, Zhang et al. reported that the FTIR spectra of TB-EPS at pH 4.5 and 10.5 showed that the amide Ⅰ vibration shifted to 1637 cm−1, which was identified as a β-fold. The amide Ⅲ vibration at pH 10.5 was attributed to random curling, while it changed to an α-helix (1304 cm−1) at pH 4.5. The results showed that a lower or higher pH altered the secondary structure of proteins [48]. Overall, the optimal pH range for microalgal EPS synthesis was 7.0–10.5, within which EPS showed extended conformations, efficient polysaccharide secretion, and stabilized functional protein structures. Too high or too low a pH affects the morphology of EPS and disrupts the protein structure of EPS.
5.
Mechanism of action of EPS in wastewater treatment using MFCS
5.1
Mechanism of pollutant removal from wastewater by EPS-mediated MFCS
5.1.1
Conventional nutrients (C, N and P)
As a structural matrix for microalgal-fungal aggregates, EPS enable physical adhesion between microorganisms through unique N—H, C═O, and C—O-C functional groups in its PN and PS fractions. This intermolecular force induces microbes to form a stable three-dimensional aggregate structure, which significantly enhances pollutant binding efficiency through electrostatic interactions, ion exchange, and chelation mechanisms, and provides a microenvironmental system conducive to nutrient delivery and storage [49]. Table 1 demonstrates the removal efficiency of MFCS for conventional nutrient elements including C, N, and P.
Table 1
Table 1.
Summary table of the removal performance of MFCS in sewage treatment.
Mechanism of EPS action on carbon metabolism in MFCS. The ability of the MFCS to remove C is flanked by changes in the amount of COD in the effluent (Fig. 5). Carbonic anhydrase produced by microalgae is able to absorb carbonate or convert it to CO2 while generating O2 and biomass for microbial metabolism and growth via the Calvin–Benson–Bassham cycle. Fungi produce laccase to degrade macromolecular organic matter into a soluble carbon source [1,5]. The O2 and proteins produced in MFCS are then consumed in a complementary manner for the conversion of carbon consumption. Fungi utilize O2 and proteins secreted by microalgae to break down large organic molecules into small soluble substances. Microalgae utilize small molecular dissolved substances produced by fungi to generate O2 and proteins through photosynthesis. Throughout the process, O2 and protein are constantly consumed and produced. Wu et al. found that the co-culture system of S. dimorphus and Aspergillus oryzae exhibited the highest TOC fixation rate (85.8%), suggesting that the fungus promoted carbon uptake by microalgae [49]. Gas exchange between fungi and microalgae facilitate the growth of each other and to some extent promotes the utilization of C in wastewater. Yang et al. treated molasses wastewater with microalgae, fungi and MFCS with COD removal efficiencies of 25.96%, 59.00% and 70.68%, which suggested that co-cultures of algae and fungi were superior to monocultures in terms of nutrient removal [7]. In MFCS, fungi may secrete enzymes to degrade organic matter and release carbon sources for microalgae to use. EPS, as substances secreted by microorganisms, may transfer and store carbon sources in the secretion process. EPS facilitate microbial contact with carbon sources and promote their uptake and utilization (Fig. 5). For example, in the treatment of swine wastewater, microalgae and fungi converted organic carbon in the wastewater into biomass through the action of EPS to achieve carbon removal and conversion [49].
Figure 5
Figure 5.
EPS-mediated removal mechanisms of conventional nutrients (C, N and P) in MFCS. The figure is originally created by the authors using Microsoft PowerPoint.
(ⅰ) Mechanism of EPS action on N metabolism in MFCS. The removal of N by MFCS is mainly reflected in the transformation of different states of N by microalgae (Fig. 5). Wu et al. in their investigation of MFCS for the treatment of pig wastewater, reported that microalgae were key in the uptake and metabolism of inorganic nitrogen sources such as NH4+-N and NO3–-N [49]. NH4+ was first taken up by microalgal cells and subsequently converted into amino acids through a series of enzymatic reactions. In this process, glutamine synthetase (GS) catalyzed ammonia and glutamic acid to form glutamine, which was then converted back to glutamic acid by glutamine 2-oxoglutarate aminotransferase (GOGAT), which also released ammonia. The ammonia can be reused to synthesize other amino acids, which are in turn used in protein or DNA synthesis. In contrast, NO3--N conversion required a stepwise reduction process, first to NO2--N and then to NH4+-N, a process that relied on the corresponding reductases, such as nitrate reductase (NR) and nitrite reductase (NiR) [49]. In MFCS, EPS may facilitate the process of nitrogen assimilation and transformation through their own functional groups. EPS enhance nitrogen metabolism through dual mechanisms mediated by their protein-polysaccharide components. First, these components provide microbial attachment sites, thus directly facilitating nitrogen uptake and assimilation. Second, the components establish inter-microbial signaling networks and material exchange channels that synergistically optimize nitrogen metabolic pathways in MFCS. This coordinated action drives the conversion of ammonia-nitrogen pollutants into either immobilized microbial biomass or gaseous nitrogen, achieving efficient nitrogen removal [5].
(ⅱ) Mechanism of EPS action on P metabolism in MFCS. MFCS remove P through assimilation by fungi (Fig. 5). For example, Mi et al. constructed a co-culture system of C. militaris and N. seminulum with microalgae to treat wastewater containing PO43- pollutants. C. militaris alone and the co-culture group of C. militaris and N. seminulum significantly affected P uptake, and the greatest P uptake was achieved by the MFCS [5]. The intrinsic mechanism of this phenomenon is a result of the EPS on the surface of microalgae that provide oxygen, amino acids, PN, PS, and other substances for fungal growth, thus achieving a greater P removal effect. In an MFCS, the presence of fungi can provide a larger specific surface area for microalgae attachment, which facilitates microalgae proliferation [16]. In their study of total phosphorus removal by S. dimorphus, Wu et al. pointed out that functional groups such as quaternary and tertiary ammonium were carried by PN in EPS. These complexed with negatively charged phosphates, leading to the rapid adsorption of phosphorus. Moreover, EPS are key in the formation and stabilization of MFCS. MFCS increases the contact area between microorganisms and phosphorus, which contributes to microbial uptake and assimilation of phosphorus, resulting in wastewater phosphorus removal [49].
5.1.2
Heavy metal removal mechanism
EPS in MFCS remove heavy metals through adsorption. Microalgae cell walls contain cellulose, hemicellulose, proteins, and other polymers [1]. A variety of anionic groups and binding sites are also present in the cell wall, which effectively adsorb and chelate cationic metals (Fig. 6). In addition, some functional fungi are able to capture and store metal ions through mycelium, and convert inorganic and methylated metal ions into volatile states [5,50]. EPS use electrostatic interactions, ion exchange chelation, and ligand mechanisms in the treatment of heavy metal pollutants, which ultimately leads to the adsorption of heavy metal substances [1]. For example, Mi et al. found that MFCS compared to a single system had a larger surface area, a more stable morphological structure, and stronger metal binding based on the degradation of mercury, cadmium, arsenic, and chromium [5]. Wang et al. further revealed through adsorption experiments that cadmium exposure specifically induced extracellular protein enrichment in TB-EPS, with distinct protein subtype variations observed under stress by other metals. This metal-responsive modulation directly validated the critical role of EPS-associated proteins in heavy metal ion adsorption, in which the proteins effectively reduced heavy metal toxicity through physicochemical sequestration [6]. In summary, the heavy metals are adsorbed by EPS through electrostatic interaction and ion exchange, while the microalgae cell wall provides binding sites that work together with the fungal hyphae to capture and efficiently remove heavy metals (Fig. 6). Table 1 demonstrates the removal efficiency of heavy metal pollutants by MFCS.
Figure 6
Figure 6.
EPS-mediated heavy metal removal mechanisms in MFCS. The Figure is originally created by the authors using Microsoft PowerPoint.
(ⅰ) Antibiotic removal mechanisms. In MFCS, microalgae and fungi are able to adsorb and accumulate pollutants through their cell walls and EPS. The adsorption of antibiotics by microalgae EPS is essentially achieved through targeted binding between the various functional groups and antibiotic molecules via specific forces, including electrostatic attraction and hydrophobic effects [51]. Electrostatic adsorption originates from the complementary charges between functional groups and pollutants. The amino groups (-NH₃+) in proteins attract negatively charged antibiotics, while the phosphate groups (-O-PO32–) in polysaccharides bind to positively charged antibiotics [51]. Han et al. found that negatively charged groups (carboxyl, hydroxyl, phosphate groups) on the surface of Chlamydomonas reinhardtii FACHB-265 provide adsorption sites for tetracycline [52]. Wu et al. further confirmed that the protein content in EPS from Chlorella pyrenoidosa FACHB-9 was positively correlated with the degradation efficiency of enrofloxacin (EFX) and ciprofloxacin (CFX), both of which validated the electrostatic enrichment mechanism [51]. The hydrophobic effect of EPS achieves specific adsorption through van der Waals forces. The amino (-NH2) and sulfhydryl (-SH) groups of proteins (PN) bind to tetracycline through hydrophobic interactions, while the dense network formed by PS and PN encapsulates the antibiotic, significantly inhibiting the internal diffusion of tetracycline [53]. Components such as PS, PN and lipids on the surface of EPS provide abundant adsorption sites that help capture and immobilize contaminants (Fig. 6). For example, Mi et al. found that the symbiotic system of C. militaris and N. seminulum was a highly efficient in degrader of four antibiotics, triclosan (TCS), lincomycin (LIN), sulfamethoxazole (SMX) and trimethoprim (TMP), in which EPS were critical in pollutant adsorption, and fungi secreted laccase to contribute to antibiotic degradation [5]. Additionally, EPS acted as a photosensitizer in the treatment of tetracycline (TC) through photocatalytic degradation of the antibiotic [54]. For example, Bai et al. found the presence of a photosensitizer in EPS in experiments evaluating the degradation of tetracycline hydrochloride by Chlorella vulgaris, with directional coupling of photosynthetic electron extraction with TC induction [54]. During the photodegradation process, the algal EPS content was significantly increased by the effects of the antibiotics. Furthermore, protein-like tryptophan, fulvic acid-like substances, and humic acid-like substances in EPS were shown to induce photocatalytic degradation of antibiotics [54]. Han et al. also reported that in the treatment of TC by Chlamydomonas reinhardtii FACHB-265, negatively charged groups on the surface of the microalgae provided adsorbate sites for tetracycline binding. As the main functional groups of EPS are carboxyl, hydroxyl, and phosphorylated groups, EPS adsorbed positively charged tetracycline through electrostatic interactions [52]. In summary, the PS, PN and lipids of EPS act as adsorption accumulators in antibiotic treatment, and as a photosensitizer for the photocatalytic degradation of antibiotics.
(ⅱ) Microplastic removal mechanisms. Microalgae show a significant particle size and concentration dependence in microplastic treatment. It is generally believed that microplastics at a high concentration and with a small particle size are more toxic to microalgae. However, the actual situation needs to consider the specific mechanism of action of algae species on microplastics. Studies have shown that small particle sized microplastics promote microalgal growth, while microplastics with a large particle size inhibit microalgal growth. Song et al. (2023) found that microalgal growth was promoted when exposed to 20-nm and 100-nm polystyrene microplastics (PS MPs), but was significantly inhibited when exposed to 1 μm PS-MPs [55]. Zhou et al.) indicated that 1 μm PS-MPs with a concentration of 5 mg/L and a particle size range of 0.2–5 μm were the most toxic to Microcystis aeruginosa at an exposure of 12 h [56]. Analyzed from the perspective of microplastic concentration, large particle sizes and high concentrations of microplastics can have a shading effect on microalgae and reduce the photosynthetic activity of microalgal cells. High concentrations of PS-MPs of a specific particle size altered the transcription of genes involved in cell division [56]. Aromatic proteins and humic acids in EPS directly mediate the adsorption and aggregation of MPs through hydrophobic functional groups (benzene rings, alkyl chains) [55,57]. The hydrophobic domains of aromatic proteins (such as benzene rings) can adsorb PS microplastics and promote cell self-aggregation. Scanning electron microscope (SEM) images show 100 nm PS particles encapsulated between microalgal cell aggregates, providing visual confirmation of the aromatic protein-mediated hydrophobic encapsulation process [55]. Humic acid induces microplastic aggregation through alkyl chains. Shiu et al. also found that EPS increases hydrophobicity and viscosity through aromatic residues [57]. Humic acid in EPS induces aggregation of MPs through its alkyl chains [58]. Its dilute water level points include long-chain fatty acids (such as 11-amino-undecanoic acid) identified in EPS [58]. Shiu et al. also found that aromatic residues in EPS can enhance the hydrophobicity and viscosity of the system [57]. In addition, Li et al. confirmed through 3D-EEM that 3 μm PS promotes the secretion of more aromatic proteins (tryptophan, tyrosine) by Chlorella, and EPS acts as a flocculant to promote the aggregation of MPs-microalgae complexes [59]. Song et al. also reported a significant increase in humic acid (Region V) after exposure to 20 nm PS, which mediated microplastic aggregation through alkyl chains [55]. Interest is the ability of microalgal EPS to mitigate the toxicity of PS-MPs. For example, Microcystis aeruginosa cells stimulated by 20 nm PS-MPs, secreted large quantities of PS, humic acids, aromatic proteins, and fulvic acids into the EPS. Among them, humic acid contributed to enveloping the microplastic surface, aromatic proteins induced self-aggregation, and fulvic acid accelerated algal growth [55]. These EPS components worked together to enhance the oxidative stress and antioxidant capacity of the algal cells, thereby mitigating the adverse effects of PS-MPs on algal cells. At the same time, aromatic proteins mediate the encapsulation and aggregation of microplastics by algal cells, while humic acid induces the self-aggregation of microplastics through alkyl chains. The EPS acted as a flocculant in this process, promoting the aggregation of microalgae with PS-MPs [59]. Similarly, Jiao et al. concluded in their experiments using Microcystis aeruginosa PCC7806 and PS-MPs that small particle size microplastics may increase the concentration of humic-like substances and enhance the oxidative stress capacity of microalgae [60]. Furthermore, microalgae were shown to develop an integrated defense against MP stress to mitigate small-sized MP toxicity, which is consistent with the conclusion of Song et al. [55]. Using 3D-EEM fluorescence spectroscopy, humic acid-like substances were found to be produced from TB-EPS of Microcystis aeruginosa PCC7806 [60]. In summary, microalgae EPS can reduce the toxicity of polystyrene microplastics. When microalgae are stimulated by a large number of secretion of proteins of EPS, together with the enhancement of algal cell stress resistance. EPS acts as a flocculant to promote aggregation during the treatment of microplastics. Small particle size microplastics can also add humic acid-like substances that enhance the oxidative stress of microalgae to reduce their toxicity (Fig. 7).
Figure 7
Figure 7.
EPS-mediated removal mechanisms of emerging contaminants in MFCS. The figure is originally created by the authors using Microsoft PowerPoint.
(ⅲ) Persistent Organic Pollutants (POPs) removal mechanisms. POPs, such as those found in cyanide, pesticide, dye, and antibiotic industrial waste streams, are generally characterized by persistence, high toxicity, and complex compositions. Biosorption is an important means of treating difficult-to-degrade organic compounds, particularly POPs. The adsorption of POPs by EPS is primarily driven by hydrophobic interactions and the complexation of polar functional groups. Concurrently, the protein and polysaccharide networks encapsulate POPs, and enzymatic reactions both inside and outside the cells degrade them. Hydrophobic interactions are commonly observed in the interactions between EPS and POPs, mediated by the hydrophobic regions (alkyl chains, aromatic rings) of proteins and the nonpolar segments of polysaccharides in EPS [61,62]. Chen et al. used 3D-EEM analysis to show that at high (ibuprofen) IBU concentrations, the signal from small aromatic proteins (Region I) in EPS was enhanced, indicating that the benzene ring structure of IBU was integrated into the EPS components [61]. Similarly, Satpati et al. also pointed out that the hydrophobic components of proteins in EPS bind to the aromatic rings of PAHs through van der Waals forces [63]. The polar functional groups in EPS, such as carboxyl groups (-COOH), hydroxyl groups (-OH), and phosphate groups (-O-PO32–), can adsorb and aggregate with PAH molecules through hydrogen bonds and electrostatic interactions. Proteins and polysaccharides form gel-like networks that encapsulate PAHs within their structures, limiting their migration. Ultimately, PAHs are hydroxylated through enzyme-catalyzed reactions (such as monooxygenases and dioxygenases), increasing their water solubility and bioavailability [63]. Not only that, EPS also participates in biotoxicity regulation. Under PBDEs stress, microalgae (Chlorella sp.) rapidly secrete EPS, with proteins increasing by 51%−169% and polysaccharides increasing by 169% within 6 h, forming a physical barrier to reduce the entry of pollutants into cells [64]. Extracellular proteins are the main components of PBDEs adsorption. Lv et al. found that within 6 h of exposure to BDE-47, extracellular proteins increased by 51%, while the bioaccumulation of BDE-47 reached its peak. Similarly, exposure to high concentrations of BDE-209 resulted in a significant increase in extracellular proteins that were positively correlated with BDE-209 accumulation [64]. On the one hand, PBDEs (such as BDE-47 and BDE-209) are hydrophobic organic compounds that bind to the hydrophobic regions of proteins in EPS (such as alkyl chains and aromatic rings) through van der Waals forces. On the other hand, the polar functional groups of proteins (-NH2, -COOH) chelate and adsorb PBDEs [64].
(ⅳ) Removal mechanisms of endocrine disrupting chemicals (EDCs). The removal of EDCs mainly relies on the dual mechanisms of adsorption and degradation of EPS. EPS adsorbs EDCs through the functional groups (-OH, -COOH, -NH2) of polysaccharides and proteins, and achieves degradation through photocatalytic oxidation and enzymatic reactions [65]. During the adsorption process, hydrophobic forces play a dominant role. Sole et al. found that the hydrophobic properties of the alginate matrix significantly increased the BPAF adsorption rate to 80% [66]. Zhou et al. also pointed out that OPE exposure causes an increase in the PN/PS ratio of EPS, and that the hydrophobic groups (alkyl/aromatic rings) and charged groups provided by PN enhance the adsorption of EDCs [67]. Yang et al. found that under NP and BPA stress, the content of hexoses (MAN, GAL, GLC) in EPS of Tetrahymena increased, and arabinose in tightly bound EPS enhanced pollutant binding through a hydrophobic furan conformation [68]. The degradation process is mainly achieved through photocatalysis and enzymatic hydrolysis working together. EPS produces reactive oxygen species (ROS, such as •OH) under light exposure, oxidizing and degrading EDCs (such as E2 and EE2) [69]. Among these, furanoic acid-like substances (C1) act as photosensitive components, with their fluorescence intensity increasing with rising OPE concentrations. They degrade 41.53%−54.50% of OPEs via ROS [67]. In general, high concentration of EDCs can damage cells, negatively affect microalgae growth, and even cause cell death. However, low concentrations of EDCs can be used as a source of nutrients by microalgae and enhance the activity of certain enzymes, and may be removed through photolysis, biodegradation, biosorption, and accumulation [65,66]. PN, PS, and lipids in the microalgae EPS were shown to promote partial photolysis of EDCs [65], similar to a study by Bai et al. [54]. The hydroxyl, phosphate, amino acid, and carboxyl groups of EPS can promote biosorption and bioaccumulation. Proteins of EPS were found to help form an external digestive system for processing EDCs, promoting their degradation by microalgae, while PS contributed to pollutant adsorption [68]. Microalgae metabolism was shown to be accelerated during photosynthesis and respiration when stimulated by NP and BPA, which increased EPS production to cope with EDCs stimulation. Therefore, microalgae enhanced the biosorption and accumulation of EDCs through EPS functional groups such as hydroxyl and carboxyl groups (Fig. 7), thereby promoting the EDCs degradation process.
Summarizing, the mechanisms of action of EPS in MFCS include providing active sites to increase biological adsorption and accumulation, reducing the toxic effects of pollutants on microorganisms, and ultimately promoting the removal of POPs. Table 1 demonstrates the removal efficiency of MFCS for emerging pollutants.
5.2
Practical applications of MFCS in wastewater treatment
In wastewater treatment applications, MFCS can improve the tolerance of microalgae to highly concentrated and complex pollutants in wastewater [70]. Most filamentous fungi also have good adsorption properties for heavy metal ions, which can further enhance wastewater treatment [2]. During the treatment of wastewater, pollutants are first adsorbed on the EPS of the microalgae, then slowly transported to the algal cells where the pollutants are adsorbed and utilized by the microorganisms through metabolic processes, ultimately converting them into microalgae biomass [71]. Microalgae can be used for front-end treatment to improve the efficiency of the wastewater treatment process, and for end-of-pipe treatment to ensure the discharge of safe, treated water.
For example, Hemalatha et al. successfully combined a batch reactor (SBR), bioelectrochemical treatment (BET) and microalgae treatment to overcome the drawbacks of incomplete treatment of effluent from a single process and to obtain high value-added products such as biomass, carbohydrates and lipids [72]. Sheetal Kishor Parakh et al. linked activated sludge in an SBR with a photobioreactor (PBR) through a hydrophobic, microporous polypropylene hollow fiber membrane to establish a link between the two reactors, thus achieving the benefit of both SBR and PBR systems. The exchange of CO2 and O2 between the microalgae and activated sludge reduced the cost of external aeration in the SBR process, and at the same time reduced the COD in the effluent. Incorporating microalgae balanced the N and P in the SBR process, which enhanced the efficiency of N and P removal in the SBR and prevented changes in the microbial community [73]. A combination of microalgae and the SBR promoted the simultaneous nitrification and denitrification process (SND) in the SBR compared to the conventional SBR process. Jin et al. found that the concentrations of NH4+-N, NO3--N, and NO2--N were lower in MB-SBR than in C-SBR by comparing the performance of a microalgal batch-sequencing batch reactor (MB-SBR) with that of a conventional sequencing batch reactor (C-SBR), which was attributed to microalgae forming a continuous anoxic microenvironment that promoted denitrification. Microalgae can stimulate bacteria to release EPS to form microalgal-bacterial flocs, which create a continuous anoxic microenvironment and promote denitrification as the flocs gradually increase in size. Moreover, microalgae were shown to increase the species richness of activated sludge and render the biological structure of the MB-SBR more stable. The two communities of bacteria and microalgae in the MB-SBR cooperated with each other, which may have contributed to the system’s ability to perform SND [30]. Therefore, integrating the microalgae and SBR process compensated for the shortcomings of a single process and enhanced the efficiency of wastewater treatment. It also generated biomass, lipids, and other high value-added products. The combination can also enhance wastewater treatment efficacy and resource recovery potential, providing a new direction for sustainable wastewater treatment technologies.
6.
Harvesting mechanisms of EPS in MFCS
6.1
Mechanism of EPS flocculation in MFCS
EPS are mainly composed of polysaccharides and proteins. These are the products of microbial metabolism, which allow cells to withstand harsh environments, and act as an good efficient flocculant [74]. The EPS flocculation mechanism is shown in Fig. 8. Its mechanism of action for microalgae collection can be described according to three important aspects: structure, charge, and molecular weight.
Figure 8
Figure 8.
Flocculation mechanisms of EPS in MFCS. The Figure is originally created by the authors using Microsoft PowerPoint.
(ⅰ) The EPS structure is affected by its composition, which is rich and diverse and includes PS, PN, nucleic acids, and other biomolecules. EPS realize efficient flocculation through the synergistic effects of multiple components and their conformational changes in the flocculation process (Fig. 8). The PS in EPS exert synergistic effects due to their long main chains and functional side chains that provide a flocculated chain structure. For example, Yuan et al. found that the EPS polysaccharide backbone of Bacillus megaterium TF10 consisted of rhamnose, glucose, and galactose, while the side chains contained groups such as glucuronic acid, acetylamino sugar, and protein [74]. The long main chain provided skeletal support for the floc, while a large number of binding sites were formed by the carboxyl group of glyoxylate, the amino group of acetylamino sugars, and other groups, which enhanced the adsorption capacity. Protein structural changes facilitated a smooth flocculation process. The secondary structure of the EPS PNs changed significantly during the flocculation process, with the proportion of random curls increasing from 24.08% to 30.24%, the proportion of α-helical structures decreasing from 31.41% to 25.86%, and the proportion of β-antiparallel structures increasing slightly from 0.54% to 1.31%. These conformational changes rendered the protein molecules more flexible and easier to bind to other substances, thus promoting flocculation [74]. The flocculation process was supported by the action of functional groups, in that the functional groups of the EPS, such as -OH, -COO-, C-H, and -NH2, showed characteristic vibrations during flocculation. Ahmad et al. (2016) observed that hydroxyl and carboxyl groups coordinated with Zn2+ ions to form intercellular bridges, increasing the flocculation efficiency up to 86.7% for S. quadricauda [9]. In addition, proteins in the amide Ⅰ (1700–1600 cm−1), amide Ⅱ (1600–1500 cm−1), and amide Ⅲ (1300–1200 cm−1) protein bands, and the 1200–900 cm−1 band associated with PS and nucleic acids, were found to help regulate flocculation functions through mechanisms such as hydrogen bonding and electrostatic interactions [74]. In summary, the flocculation functions of EPS are dependent on the physical adsorption network of polysaccharide chains, the dynamic fitness of protein conformations, and chemical bridging among functional groups. The three synergistically form a multilevel structural response mechanism to ultimately realize efficient flocculation efficiency.
(ⅱ) From a charge perspective, the flocculation process of EPS is essentially a result of the synergistic effect of their surface charge properties and cation-mediated charge neutralization and bridging mechanisms (as shown in Fig. 8). PN, PS and other macromolecules in EPS carry ionizable functional groups such as carboxyl, phosphate and hydroxyl groups. At a near-neutral pH, the carboxyl groups and other groups dissociate to confer EPS with a negative charge [75]. The negative electrical characteristics of EPS are neutralized by cations (such as Zn2+, Ca2+, Mg2+) through electrostatic action, reducing the electrostatic repulsive force and realizing flocculation. For example, Ahmad et al. used Zn2+ to neutralize the negative charge of Scenedesmus quadricauda EPS and induce flocculation [9]. Yu et al. (2009) found that Ca2+ induced the adsorption of negatively charged TB-EPS onto kaolin particles by cation bridging, which achieved a flocculation efficiency of 54.1% [76]. Cations act as bridges to EPS molecules or particles while neutralizing a charge, and cells are subsequently adsorbed to the bioflocculant chain to form bridging structures. TB-EPS in activated sludge bridges negatively charged particles via Ca2+ to form stabilized flocs [9,76]. In addition, cationic polymers neutralized the negative surface charge of mature cells of Chlorella vulgaris, and enhanced the bridging effect through a long-chain structure, resulting in a flocculation efficiency of 97% [77]. The electronegativity of EPS originates from carboxyl, phosphoryl, and hydroxyl ionizable groups in the molecular structure, and flocculation is achieved through cation-mediated electrostatic interactions.
(ⅲ) The molecular weight of EPS affects their structure, as high molecular weight flocculants have higher flocculant activity and larger flocs [74,76]. High molecular weight EPS contribute to a significant increase in the size of microalgae-fungal aggregates. For example, EPS secreted by Penicillium sp. AHP141 increased the harvesting efficiency of Chlorella sp. DT025 by enhancing the surface roughness of microalgal cells and forming larger floc structures [78]. PN, PS, and humic acid substances of EPS were shown to be the core drivers of flocculation through a fluorescence-based characterization of a co-culture system of Aspergillus niger HW8–1 and Chlorella sp. HQ [8]. In addition, the high molecular weight components of fungal EPS (e.g., xanthates, humic acid analogs) enhance the surface adsorption of microalgal cells through long-chain structures and abundant functional groups (e.g., -OH, -NH). For example, proteins, complex amino acids, and tryptophan in Aspergillus niger X-5 EPS were shown to be involved in the flocculation process of Scenedesmus sp. Z-4. A 3D-EEM analysis showed that these components dynamically responded to flocculation activity [79]. The molecular weight properties of fungal EPS work in conjunction with functional groups (e.g., hydroxyl groups, amino acids) to promote the flocculation process (Fig. 8). For example, cation bridging and hydrogen bonding enabled Scenedesmus sp. Z-4 to form stable flocs with the EPS of Aspergillus niger X-5 [79]. The EPS of Penicillium sp. AHP141 enhanced the strength of physical binding with microalgal cells due to its richness in polar groups [78]. Fig. 8 shows the harvesting efficiency of different MFCS. In summary, high molecular weight organics (e.g., PN, PS) in EPS enhance the microalgae flocculation efficiency through functional group interactions, size modulation, and interface interactions, to enhance microalgal flocculation efficiency. Table 2 demonstrates the environmental factors and harvesting efficiency of fungal-assisted microalgae harvesting.
Table 2
Table 2.
The harvesting efficiency of different MFCS.
6.2
Influence of external environmental factors on MFCS harvesting efficiency
Microalgae are captured by filamentous fungi through a triple mechanism of mycelial wrapping, EPS bridging, and direct cell wall adsorption. This process provides significant advantages, such as not requiring an investment in additional equipment, separation achieved through simple filtration, and significantly reduced harvesting costs. This type of harvesting method keeps the microalgal cells intact, while the fungi and microalgae form a mutually beneficial symbiotic system, which synergistically produces high value-added products, such as high-value oils and bioactive substances. During the harvesting process, external environmental factors can affect the efficiency of fungal harvesting of microalgae, such as the culture method, temperature, pH, incubation time, rotational speed, and nutrient composition.
The form of microalgae captured by fungi in different culture modes varies. There are two main modes of harvesting microalgae using fungi: FSH and FPH. In FSH, microalgae-fungal aggregates are formed that are more robust compared to those produced by FPH [8]. A scanning electron microscope (SEM) image analysis by Pei et al. of cultures formed using both methods showed that a large number of algal cells were attached to the mycelial network in the FSH aggregates, while only a few microalgal cells were seen on the FPH aggregates [79]. This probably stemmed from the difference in culture mode, in which fungal spores developed into hyphae before encapsulating microalgal particles in the FSH mode. In FPH, the fungi are cultured first and then separated and added to the algal medium after they produce fungal mycelia clusters. At this point, the organic acids, PN, fungal surface EPS enzymes, fungal cell wall humic acids, and amides Ⅰ and Ⅱ collectively trap the microalgae. A comparison of SEM images of two cultures, of Chlorella sp. HQ and Aspergillus niger HW8–1 by Wang et al. similarly found that many microalgae were attached to the mycelial network of FSH particles, and only a small amount of algae was observed within the FPH particles [8]. This was consistent with the findings of Pei et al. [79]. In the FSH mechanism, the LB-EPS and TB-EPS of the fungal clusters were modified to contain many soluble microbial products in addition to aromatic proteins in regions Ⅰ and Ⅱ. With the FPH mechanism, aromatic proteins were involved in the EPS and a large amount of negatively charged proteins and humic acids were released from the fungal clusters. EPS contains large amounts of PN, PN, humic acid-like compounds, and glyoxal, which confer the fungal cells a high adsorption capacity [8].
Other environmental factors, such as temperature, pH, incubation time, rotational speed, and nutrient composition affect the efficiency by which fungi harvest different microalgae [80-82]. It was shown that the microalgal flocculation efficiency significantly increased at higher temperatures. Chen et al. found that the FSH method achieved up to a 99% flocculation efficiency of Chlorella sp. at 40 ℃, and high temperatures accelerated the formation of Penicillium sp. particles, thus shortening the harvesting time. The FPA method achieved a 98.26% of algal cells at 34 ℃, indicating that temperature affected the efficiency of microalgae harvesting using different fungi [83]. At high temperatures, the activity of microalgae was found to be reduced and weaker; however, high temperatures promoted fungal particle formation, which is conducive to MFCS formation [83]. pH is also a key influencing factor. Prajapati et al. showed that Aspergillus lentulus efficiently harvested Chroococcus sp. cells in environments with a pH of 8.6–9.2, and that nearly 100% of microalgal cells were harvested within 24 h [84]. Lower pH favored harvesting efficiency. The attraction between Chlorella sp. and Pleurotus ostreatus hyphae was enhanced, with harvesting efficiencies of up to 64.86% within 150 min [10]. This was because under acidic conditions, EPS present a curled state, while under alkaline conditions, they present a stretched chain structure, which is more favorable for presenting the protein binding sites required for flocculation. In a high pH environment, a large number of Ca2+, Mg2+, and PO43− ions in the wastewater will react to form positively charged precipitates, such as calcium phosphate and magnesium hydroxide. These precipitates can induce microalgae self-flocculation through electrical neutralization [83]. Incubation time and agitation rates also affect fungal harvesting of microalgae. Pleurotus ostreatus formed pellets when incubated at an agitation speed of 100 rpm, which harvested Chlorella sp. with a high efficiency (Table 2) [10]. In a co-culture of Aspergillus oryzae and Chlorella pyrenoidosat, Chlorella pyrenoidosa cells were gradually adsorbed by fungal particles over time, at 72 h, free algal cells could were completely harvested by fungal particles [85]. A suitable incubation time, logarithmic growth, vigorous metabolism, secreted metabolites, and enzymes all promote interactions with microalgae to effect harvesting. Excessive incubation times, nutrient depletion, and metabolic waste accumulation can inhibit fungal growth and harvesting efficiency. If the incubation time is too short, the fungi will not be able to pelletize [84]. Additionally, appropriately increasing the stirring speed can increase the contact area and frequency of contact between fungi and microalgae, thus promoting their interaction. However, a mixing speed that is too fast may produce a water current shear force and destroy the fungal hyphae and microalgae cell structure, reducing the harvesting efficiency [83]. Finally, the nutrient composition also affects the harvesting efficiency. Prajapati et al. found that Aspergillus lentulus had the best harvesting efficiency of Chroococcus sp. in the presence of 10 g/L glucose, 0.5 g/L NH4NO3, and 0.5 g/L K2HPO4 [84]. This is because when nutrients are sufficient, the metabolic activity of microalgae and fungi are enhanced, which promotes the synthesis and accumulation of the appropriate substances [8].
In general, the use of fungi to harvest microalgae should consider the culture mode, regulation of environmental parameters, and optimization of operating conditions. First, FSH or FPH should be selected according to the needs of the target products. Second, precise control of temperature, pH, incubation time, stirring speed, and nutrient ratios must be considered. Ultimately, the process technology should strike a balance between cost and increasing the value added to achieve low-cost and high efficiency harvesting of microalgae.
7.
Challenges and prospects
Microalgae and fungi form a stable system that enhances wastewater treatment with resistance to pollutants. Filamentous fungi can be used as a natural flocculant in microalgae harvesting to capture the microalgae, which protects the morphology of microalgal cells and produces a very high harvesting efficiency. However, there are still limitations in the application of microalgal-fungal systems that restrict their practical application to some extent.
The components of the EPS of different algal and fungal species vary significantly, resulting in a lack of universal screening criteria for constructing symbiotic systems (Fig. 9). Once an MFCS has been generated, it is not necessarily clear whether the algal-fungal system is safe or will consume or damage the target biomass. In the future, we should strengthen our understanding of the EPS composition and structure of various microalgae and fungi and study their growth characteristics under different environmental conditions to determine the optimal combination of algal species and strains. And with the help of advanced biotechnological means, such as gene sequencing, proteomics, we should conduct in-depth studies to elucidate the signaling pathways and regulatory mechanisms within the polymers of the algal system. By analyzing the chemical composition and structural characteristics of EPS and the growth conditions of microalgal fungi, we can explore whether toxic interactions between microalgae and fungi occur during the growth process.
Figure 9
Figure 9.
Challenges and prospects. The figure is originally created by the authors using Microsoft PowerPoint.
The ability of MFCS to treat pollutants needs to be further explored. Firstly, the problem of microalgae light energy limitation and functional site masking caused by mycelial wrapping. The symbiosis formed by microalgae and fungi strengthens the overall capacity of the system and makes it more stable, but some of the microalgae in the symbiotic culture is adsorbed on the surface of the mycelium, and some of the microalgae is wrapped inside the filamentous fungi. When using pollutant-adapted microalgae, the microalgae wrapped inside the fungi are unable to remove the pollutants, and thus the overall system is not able to remove the pollutants. Similarly, with microalgae bound to the fungal surface, the number of protein binding sites of the fungi are reduced, and the pollutant removal efficiency of pollutant-adapted fungi may be reduced compared to that of the pure fungal group. Fungus-wrapped microalgae may reduce light transmission, with the microalgae receiving less solar energy for photosynthesis. The growth rate of microalgae is inhibited with low transmission of light, which reduces the biomass of microalgae. Second, MFCS have not been well-studied for use with new pollutants (e.g., POPs and EDCs) in wastewater, and there is a lack of mechanistic investigations of EPS in MCFS for these two new pollutants. There is a lack of research on the functional group mechanisms of EPS in FITR and XPS in studies dealing with EDCs and POPs. It is also unclear whether there is a specific synergistic mechanism between fungal EPS and microalgal EPS in the treatment of pollutants. The specific mechanism by which EPS secreted by fungi treats pollutants has not yet been quantified. The dynamic patterns of changes in the composition and structure of EPS during the removal of pollutants by MFCS are still unclear. Future research should focus on how to improve the ability of symbionts to manage pollutants and develop innovative symbiotic structures and cultures. Enabling microalgae or fungi to function better in symbiotic systems may solve issues with the limited capacity of microalgae or fungi. For example, creating novel reactor designs or light regulation strategies to adjust the light intensity and wavelength in real time according to microalgae growth needs and light conditions may ensure that microalgae can perform sufficient photosynthesis. At the same time, we will increase the research on the degradation of new pollutants in water by the algal-fungal system, integrate genomics and proteomics technologies, analyze the EPS-mediated metabolic signaling pathway. Further exploration of the mechanism of action of EPS on POPs and EDCs in MFCS. Using multi-omics integration technology (transcriptomics combined with Fourier infrared spectroscopy) to analyze the role of key components of EPS (such as C—N and C—H groups). Using genetic engineering to explore the synergistic mechanism of microalgae and fungi EPS in pollutant treatment. Establish a database of pollutant components and EPS functional group interactions to predict EPS specificity mechanisms and quantify the dynamic changes of EPS during the treatment process (Fig. 9).
There are also many challenges in harvesting microalgae from fungi. The two important harvesting methods, FSH and FPH, can realize efficient harvesting, but challenges remain. For example, FSH is harvested by co-culturing filamentous fungal spores with microalgae. This is a lengthy process, usually 24–48 h, which increases production costs and may lead to contamination or metabolic changes in microalgae during prolonged cultivation that affect their quality [83]. FPH has demonstrated good harvesting performance under acidic pH conditions. However, cultivating fungal particles prior to FPH requires additional time and cost, and the cultivation conditions are demanding [83]. Moreover, the contribution weights of EPS in flocculation processes such as electrostatic neutralization, surface protein interaction, and polysaccharide adhesion during microalgae harvesting have not been quantified. EPS secreted by different algal systems varies in composition and content, and there is no universal method for harvesting microalgae. Process conditions of the fungal harvesting of microalgae should be improved in the future. Fungal strains can be screened and improved to enhance their capture ability and adaptability to microalgae, shorten the harvesting time, and reduce costs. For example, efficient fungal strains can be screened from the environment or existing strains can be modified using genetic engineering techniques. Additionally, the advantages and disadvantages of FSH and FPH cultivation methods should be studied in depth. Optimize the cultivation conditions according to the actual needs, and determining the best mixing ratio, cultivation time and temperature of different algae and bacteria species under different cultivation methods. A standardized harvesting process and quality control system should be established to ensure that the harvesting process is efficient, stable, and sustainable to achieve industrialized fungal harvesting.
8.
Conclusion
This study reviewed the definition and formation of MFCS, the properties of and EPS and the factors that influence these properties, and the mechanisms of EPS action in wastewater treatment and microalgae harvesting. Microalgae and fungi were shown to form a mutually beneficial symbiotic system through electrostatic attraction between functional groups and protein bridging mechanism in EPS. EPS were found to participate in the removal process of C, N, and P from wastewater through signaling, substance exchange, and functional group capture, and to realize the adsorption of heavy metals through electrostatic interactions, ion exchange, and chelating-ligand mechanisms. EPS also help MFCS degrade new pollutants through adsorption accumulation and enhanced oxidative stress. During harvesting, the physical adsorption network of polysaccharide chains, dynamic fitness of protein conformation, and chemical bridging of functional groups of EPS together promote efficient flocculation of microalgae and fungi. Although EPS show great potential in MFCS, they still face certain challenges. Future research should focus in the following directions: (i) Mechanisms of action of EPS in MCFS on POPs and EDCs; (ii) signaling pathways and regulatory mechanisms within the algal-fungal system; and (iii) optimization of microalgae harvesting conditions, superior strain screening, and the harvesting potential of fungi. Future research should be combined with practical engineering applications, which will further promote scalable MFCS. In summary, the study of the mechanisms of EPS in MFCSs provides new ideas for solving the problem of wastewater treatment and resource recovery, which has important environmental and economic significance.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
This work was supported by the National Natural Science Foundation of China (Nos. 42307347 and 42567062), Guizhou Provincial Basic Research Program (Natural Science) MS [2025](611), Guizhou University High-Level Talent Research and Platform Construction Funds (No. 600195233301), the Science and Technology Innovation Team of Education Agency in Guizhou Province (Qian Jiao Ji[2023]056).
Figure 1
(a) WOS literature study on microalgae, fungi, EPS, and wastewater treatment in the last five years. (b) MFCS formation, wastewater treatment, microalgae harvesting process. (c) Keyword clustering map, and (d) keyword co-occurrence map. The figure is originally created by the authors using Microsoft PowerPoint and Citespace.
Figure 3
Structural definition, compositional features, and characteristics of EPS. The figure is originally created by the authors using Microsoft PowerPoint.
Figure 5
EPS-mediated removal mechanisms of conventional nutrients (C, N and P) in MFCS. The figure is originally created by the authors using Microsoft PowerPoint.

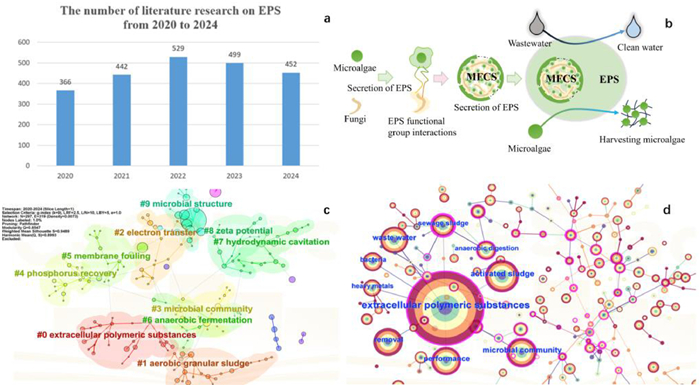
 DownLoad:
DownLoad:
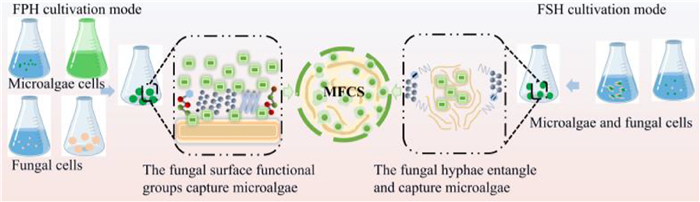
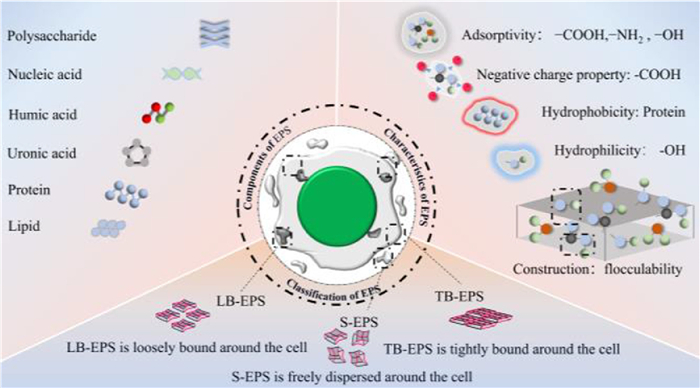
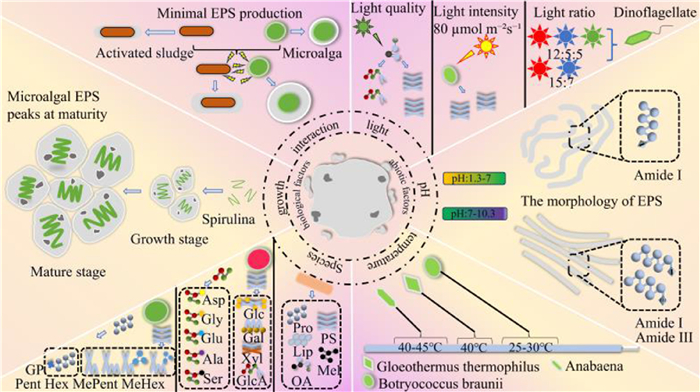
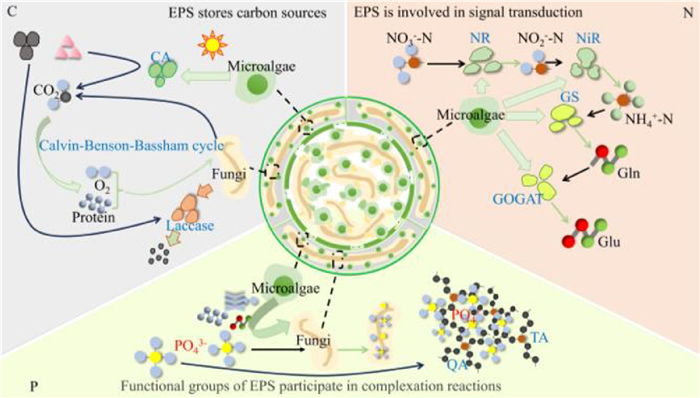
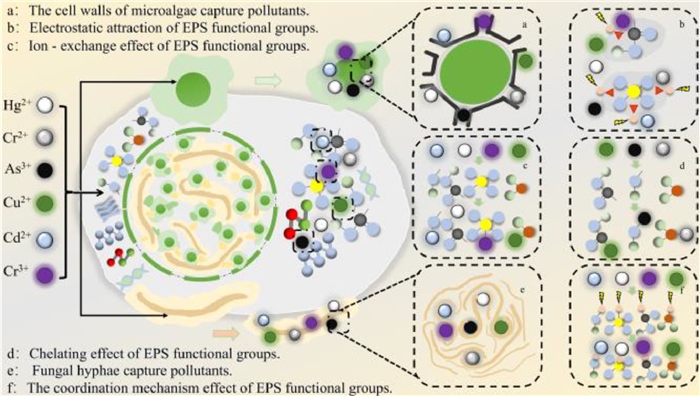
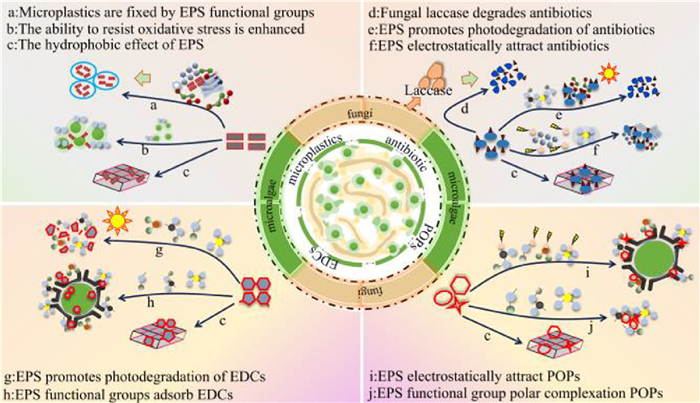
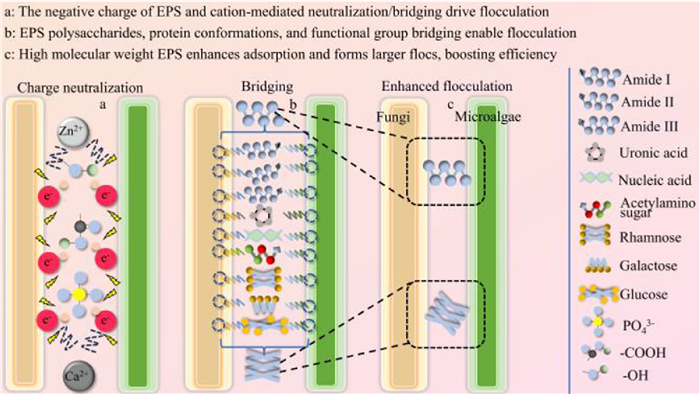
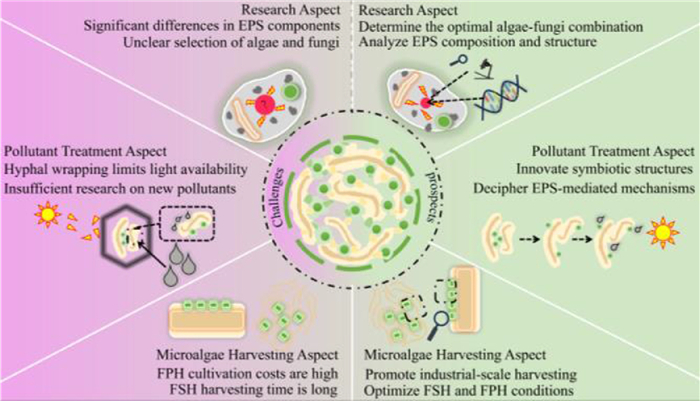

 下载:
下载:










 下载:
下载:
