
Figure 1.
Structures of commercial antibacterial agents.
Antibacterial agrochemicals: Recent research progress and outlook
Shaobo Wang , Honggui Yang , Chunni Zhao , Deyu Hu , Baoan Song

Plant bacterial diseases caused by bacteria pathogens are a major constraint on crop production and cause significant losses on a global scale [1]. Mansfield et al. listed ten plant pathogenic bacteria that have caused great harm to agricultural production, including Xanthomonas oryzae pv. oryzae (Xoo), Ralstonia solanacearum (Rs), Xanthomonas axonopodis pathovars, Pectobacterium carotovorum, Erwinia amylovora, Pseudomonas syringae pathovars, Xanthomonas campestris pathovars, Agrobacterium tumefaciens, Xylella fastidiosa and Dickeya [2]. For example, Xanthomonas infects about 400 plants and causes huge economic losses worldwide [3-5]. Among them, the rice bacterial leaf blight caused by Xoo can reduce rice production by 10%−50%, posing a significant threat to global food security [6-8]. Another example, Rs is a "species complex" and one of the most destructive phytopathogenic bacteria in the world [2,9]. The host range covers > 50 families and > 200 species [9]. Due to the wide geographic distribution and host range of Rs, it is difficult to quantify the economic impact of the disease it causes. Potato alone causes about $ 1 billion in global losses [2,10].
At present, the main methods to control plant bacterial diseases include plant host resistance, biological control and chemical control [1,11]. Current resistant varieties can effectively control bacterial diseases, but resistance has deteriorated due to the diversity of pathogenic strains and the ability of bacteria to express effectors that inhibit certain host defense responses [12]. Biological control mainly uses microbial control, but live microbial preparations are susceptible to environmental conditions and crops, showing instability of the effect [13,14]. Chemical control is a relatively quick and effective control method (Fig. 1). Currently, copper-containing bactericides and agricultural antibiotics are mainly used to control bacterial diseases [15]. However, reliance on copper-containing bactericides leads to phytotoxicity, the spread of copper-resistant strains, and soil accumulation and adverse effects on soil biota [16-18]. The continued use of antibiotics has led to significant resistance. What is more, concerns about the potential impact of antibiotics on human health have led the European Union to restrict the use of antibiotics [19-22]. Furthermore, the scarcity of other types of antibacterial agents, unsatisfactory control effects and the development of bacterial resistance make it greatly difficult to control crop bacterial diseases [23,24]. Therefore, it is an urgent issue to search for new, efficient and antibacterial agrochemicals in the field of pesticide science.
In recent years, researchers have focused on metal materials [25,26], peptides [27,28], microbial metabolites [29,30], separation from natural substances [31,32], and modification of lead compounds in the control of plant bacterial diseases. So far, there is no review report on structural modification of antibacterial agrochemicals. In this review, the antibacterial agrochemicals in recent 10 years are divided into oxadiazole/thiadiazole, triazole, isopropanolamine, onium salts, quinazolines, flavonoids, amides, esters, metal complexes, and others according to the active skeleton. The research progress of antibacterial agrochemicals is summarized in terms of structure-activity relationships (SAR) and mechanisms of action. Finally, the future development trend and application of antibacterial agrochemicals were prospected.
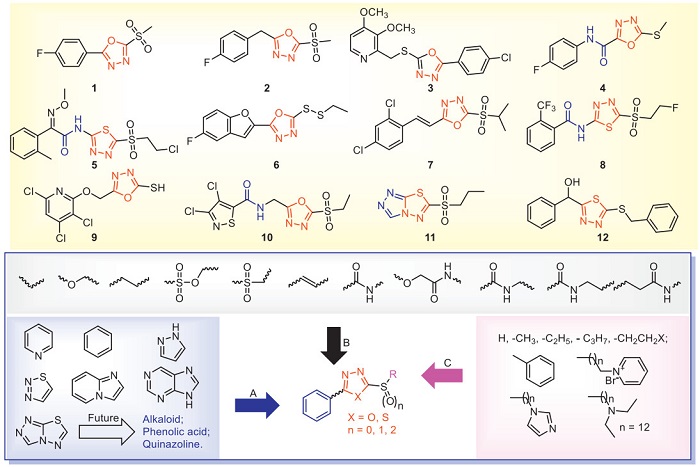
Oxadiazole/thiadiazole is a nitrogen-containing heterocyclic compound that can induce the host to defend against plant pathogens [33]. The successful commercialization of thiadiazole compounds such as thiodiazole copper, zinc thiazole and bismerthiazol also proves that thiadiazole compounds have excellent application prospects and research value in the design and synthesis of new and efficient antibacterial agents. Liang et al. found that bismerthiazol reduces the virulence of Xoo by inhibiting the histidine utilization pathway and quorum sensing, thereby achieving the purpose of antibacterial [33]. In addition, the degradation product of bismerthiazol contains 2-amino-1,3,4-triadiazole and N, N'-methylene bis(2-amino-1,3,4-thiadiazole), which have good antibacterial activities. However, these two degradation products are banned due to teratogenic side effects in humans and animals [34]. Recently, many compounds containing oxadiazole/thiadiazole have been developed to study their antibacterial activities in order to search for potential antimicrobial agents. The antibacterial activities of some representative oxadiazole/thiadiazole compounds are shown in Table 1 [35-50]. Compounds 1, 6, 7 and 11 increase the activities of superoxide dismutase (SOD) and peroxidase (POD) in rice, thereby enhancing the resistance of rice to Xoo. In addition, compounds 1 and 11 increase the chlorophyll content of rice and inhibit the increase of malondialdehyde content in rice, thereby reducing the damage of Xoo to rice. This suggests that the structure of oxadiazole/thiadiazole increases the content of defense enzymes and chlorophyll, and improves the antibacterial ability of the host. In addition, at a concentration of 20 µg/mL, the inhibitory rates of compound 1 on the expression of extracellular polysaccharide (EPS) related genes gumB, gumG, gumM and xanA were 94.88%, 68.14%, 86.76% and 79.21%, respectively [35]. Chen et al. used "activity-based protein profiling (ABPP)" technology to find that the specific target of compound 1 against Xoo and X. axonopodis pv. citri (Xac) was dihydrolipoamide S-succinyltransferase (DLST). This result was further confirmed through in vitro interaction analysis and Western blot experiments [36,37]. Gao et al. used a label-free proteomics quantitative analysis method to screen the differential protein of compound 2 that acts on citrus canker. They found that compound 2 has a significant regulatory effect on the metabolism of pyrimidine [39]. Compound 3 regulates the differential proteins of the rice purine metabolism pathway, and the down-regulated differential proteins are adenylosuccinate lyase and N5-(carboxyamino)imidazole ribonucleotide mutase [41]. Compound 4 disrupts Xanthomonas oryzae pv. oryzicola (Xoc) growth and pathogenicity by interfering with pathways associated with bacterial virulence. Translational regulator CsrA (CsrA) and virulence regulator (Xoc3530) were identified as target proteins by ABPP. Further knocking out the CsrA or Xoc3530 gene in Xoc resulted in a significant reduction in motility and pathogenicity of the mutant strains [42]. Compound 5 can regulate the differential proteins in the glycolysis/gluconeogenesis pathway. The upregulated proteins are fructose-1,6-bisphosphatase, and the dihydrolipoamide acetyltransferase component of pyruvate dehydrogenase complex, dihydrolipoyl dehydrogenase, and aldehyde dehydrogenase ALDH2b [43]. Compound 6 obstructs the physiological and biochemical processes of pathogenic bacteria by interfering with proteins in the glycolytic-gluconeogenesis pathway [44]. Compound 7 can regulate the differential proteins of the rice glutathione metabolism pathway, and the differential proteins upregulated are glutamate-cysteine ligase A and glutathione reductase [45]. Compound 9 regulates the metabolism of histidine, starch and sucrose in rice [47]. Compound 11 regulates differential proteins in rice photosynthetic pathways and upregulates proteins associated with photosystem Ⅱ and photosystem Ⅰ [49]. Compound 12 inhibited hpa1 transcription and hypersensitive response (HR) without affecting bacterial growth [50]. These results fully indicated that oxadiazole/thiadiazole structures destroy the morphology of bacteria, reduce the EPS of bacteria, increase the permeability of cell membrane, and thus reduce the pathogenic ability of bacteria. In the study of the mode of action on the host, oxadiazole/thiadiazole compounds can not only enhance the activities of SOD, POD and chlorophyll in plants, but also strengthen multiple regulatory pathways, thus enhancing the resistance to bacteria. Currently, great progress has been made in the research on the mechanism of action of oxadiazole/thiadiazole compounds in vitro. For example, new targets such as DLST, CsrA or Xoc3530 have been discovered. However, in the pathway of crop protection, further research on the targets of their antibacterial effects in vivo is of great significance for the development of antibacterial agents.
 DownLoad:
CSV
DownLoad:
CSV
| Compd | Bacterial | In vitro antibacterial activities/ EC50 (µg/mL) | Protective activities at 200 µg/mL (%) | Mechanism of action | Ref. |
| 1 | Xoo | 9.9 | / | DLST | [35,36] |
| Rs | 8.3 | / | [37,38] | ||
| 2 | Xoo | 1.1 | 43.5 | Metabolism of pyrimidine | [39,40] |
| Xoc | 7.1 | / | |||
| 3 | Xoo | 4.8 | / | Purine metabolism | [41] |
| 4 | Xoo | 5.3 | 44.3 | CsrA and Xoc3530 | [42] |
| Xoc | 7.6 | 47.5 | |||
| 5 | Xoo | 1.8 | 51.3 | Glycolysis/gluconeogenesis | [43] |
| Xoc | 2.1 | 45.9 | |||
| 6 | Xoo | 0.3 | 40.3 | Interferes with proteins in the bacterial glycolysis-gluconeogenesis pathway | [44] |
| Xoc | 0.6 | 42.0 | |||
| Xac | 10.4 | / | |||
| 7 | Xoo | 0.4 | / | Glutathione metabolism | [45] |
| Xoc | 0.2 | 35.4 | |||
| 8 | Xoo | 0.4 | 55.8 | / | [46] |
| Xac | 6.7 | / | |||
| Rs | 5.1 | / | |||
| 9 | Xoo | 13.4 | 56.0 | Histidine metabolism and starch and sucrose metabolism | [47] |
| Rs | 33.5 | / | |||
| Xac | 24.6 | / | |||
| 10 | Xoo | 0.8 | / | / | [48] |
| Xoc | 2.2 | / | |||
| 11 | Xoo | 0.6 | 49.4 | Photosynthetic | [49] |
| Xoc | 1.6 | 49.7 | |||
| 12 | Xac | / | / | Inhibited hpa1 transcription and hypersensitive response (HR) | [50] |
| Xoc: Xanthomonas oryzae pv. oryzicola, Xac: X. axonopodis pv. citri. | |||||
The modified active structural motifs of oxadiazole/thiadiazole compounds with antibacterial activities are shown in Fig. 2. According to the structural characteristics, oxadiazole/thiadiazole compounds are divided into four parts: A (aromatic ring), B (flexible chain), C (substituent) and sulfur bond. In part A, the antibacterial activity of aromatic rings is generally higher than that of alkyl or cycloalkanes. The aromatic rings are mainly heterocyclic rings containing N atoms and benzene rings. Among them, nitrogen-containing heterocycles are beneficial to improve water solubility, and improve the solubility of oxadiazole/thiadiazole compounds in water, thereby increasing antibacterial activity. Therefore, when designing part A, we can introduce structures such as quinazolines, alkaloids or phenolic acids to ensure that the activity is enhanced on the basis of increased water solubility. In part B, it is mainly the flexible chain connecting the aromatic ring and oxadiazole/thiadiazole, usually the chain of 0 to 3 carbon atoms has better antibacterial activity, and the continuous extension of the carbon chain is not conducive to the activity. The carbon atoms can be replaced with amide, amine, sulfonic acid and sulfone groups. We believe that the rational design of flexible chains has a significant influence on the antibacterial activity of oxadiazole/thiadiazole compounds. Therefore, when designing the structure, structures like isopropanolamine and esters can be introduced to improve the antibacterial activity. The introduction of small alkanes and halogenated alkanes in part C is more beneficial to the activity. The introduction of benzene and heterocyclic rings generally weakens the activity. In addition, the introduction of long-chain onium salts can also increase antibacterial activity. The sulfur between the oxadiazole/thiadiazole and part C has a vital biological activity for the compound. The antibacterial activity of the sulfone is significantly higher than that of the sulfoxide and thioether. We tried to separate oxadiazole/thiadiazole from sulfone with one or two carbon atoms, and found that the antibacterial activity decreased, indicating that the connection between oxadiazole/thiadiazole and sulfone is a necessary group for antibacterial activity. Although oxadiazole/thiadiazole compounds have excellent in vitro antibacterial activity, they do not have outstanding in vivo activity. We analyzed that the possible reasons might be the poor water solubility of oxadiazole/thiadiazole compounds and the relatively low stability after the splicing of the sulfonyl group and oxadiazole/thiadiazole. Therefore, when introducing the oxadiazole/thiadiazole structure, we should consider as much as possible how to increase water solubility and how to improve stability, which is beneficial to increasing the in vivo activity.
Triazole compounds are five-membered aromatic compounds containing three nitrogen atoms, which have broad-spectrum and high-efficiency biological activities, and are one of the current research hotspots [51]. Similarly, triazole compounds have made remarkable achievements in agricultural fungicides, and their mechanisms of action are to inhibit the 14α-demethylase involved in the biosynthesis of fungal sterols [52]. In contrast, triazole compounds have only begun to develop in agrochemical antibacterial applications. Table 2 and Fig. 3 show the antibacterial activity of some representative triazole structures [53-59]. Compounds 13 and 15 destroyed the cell membrane structure of Xoo and Xac, respectively, and achieved the effect of inhibiting Xoo and Xac, respectively. In addition, compound 13 affects bacterial growth by interfering with bacterial EPS and membrane permeability [53]. Compound 14 significantly downregulated the expression of Xoo's type Ⅲ secretion system and transcription activator-like effector-related proteins [54]. Compound 20 may trigger the pathogen's apoptosis and interfere with the balance of the pathogen's redox system, thereby inhibiting its normal growth and reproduction [59]. This suggests that the triazole structure interferes with the type Ⅲ secretion system and redox system, reduces EPS, disrupts cell morphology, and ultimately reduces the pathogenic capacity of bacteria. However, the mechanism of the current compounds with triazole structures is limited to the phenotypic analysis of pathogens by the agents and the detection of physiological and biochemical indexes. Their molecular targets and regulatory mechanisms have not yet been elucidated.
DownLoad:
CSV
| Compd | Bacterial | In vitro antibacterial activities | Protective activities at 200 µg/mL (%) | Mechanism of action | Ref. |
| 13 | Xoo | EC50 = 18.8 µg/mL | 50.8 | Destroyed the cell membrane structure | [53] |
| 14 | Xoo | EC50 = 5.0 µg/mL | 54.2 | Downregulated the expression of related proteins in the type Ⅲ secretion system and transcriptional activator of Xoo | [54] |
| 15 | Xac | EC50 = 38.2 µg/mL | 61.1 | Enhance the permeability of bacterial membrane and change the morphology of bacterial cells | [55] |
| 16 | Xoo | EC50 = 18.8 µg/mL | 56.2 | / | [56] |
| 17 | Erwinia amylovora | MIC = 0.0004 mmol/L | / | / | [57] |
| Erwinia carotovora | MIC = 0.0004 mmol/L | ||||
| X. campestris | MIC = 0.0008 mmol/L | ||||
| 18 | Xac | EC50 = 10.0 µg/mL | / | / | [58] |
| Xoo | EC50 = 7.2 µg/mL | 49.5 | |||
| 19 | Psa | EC50 = 4.6 µg/mL | 79.4 | / | [59] |
| 20 | Xoo | EC50 = 3.4 µg/mL | 53.2 | Interfere with the balance of the pathogen's redox system | [59] |
| Xac | EC50 = 2.9 µg/mL | / | |||
| Psa: Pseudomonas syringae pv. actinidiae. | |||||
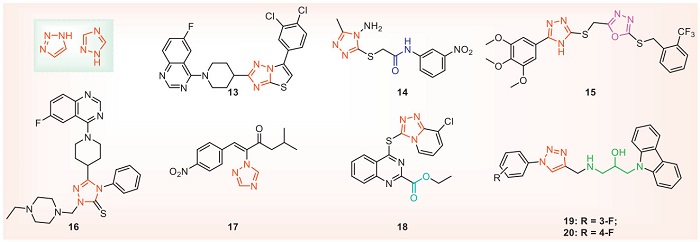
In the design of triazole antibacterial agents, the triazole part can be combined with hydrogen bonds, which is conducive to binding biomolecular targets and increasing solubility. Therefore, it is usually used as an active fragment to be spliced with natural products or lead structures with antibacterial activity. For example, compound 15 was obtained by concatenating oxadiazole and triazole, and compounds 16 and 18 were obtained by introducing quinazoline. Both carbon and nitrogen atoms of triazole can be structurally derived. The introduction of sulfur atoms and flexible chains such as isopropanolamine on the carbon atoms may help improve the antibacterial activity. At the same time, when designing new structures, the addition of triazole or triazole-containing condensed ring structures can be considered to improve antibacterial activity and water solubility. In addition, it is also a good choice to introduce triazole skeleton with antifungal activity into the structure of antibacterial activity. For example, compounds 19 and 20 introduce triazole containing isopropyl alcohol into carbazole, and the antibacterial activity is significantly increased.
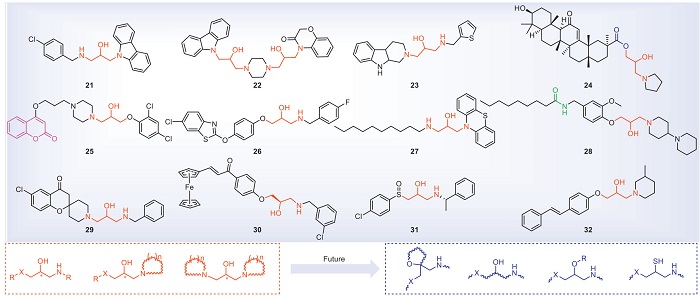
Structures containing isopropanolamine are easy to be derivatized and are a kind of advantageous lead structures. Especially in the field of antifungal research, in-depth studies have been carried out, and commercial drugs such as the pharmaceutical fluconazole (Diflucan) and the pesticide triadimefon have been developed. In recent years, compounds containing isopropanolamine have attracted wide attention due to their good antibacterial activity against plant pathogenic bacteria. The structures of the isopropanolamine compounds with antibacterial activities are shown in Fig. 4, and antibacterial activities are shown in Table 3 [60-71]. At 25 µg/mL, compound 21 can wrinkle or rupture cell membranes and interact with Xoo's DNA. Through proteomics technology, it is found that compound 21 mainly affects Xoo's starch and sucrose metabolic pathways and biotin metabolism [60]. Studies have shown that compounds 23 and 24 may disrupt the balance of the bacterial redox system, thereby inducing different apoptotic behaviors on the pathogens Xoo and Xac [62,63]. Compound 25 inhibits diffusion signaling factor (DSF) biosynthesis, biofilm formation, EPS, and related quorum sensing (QS) pathway proteins. In addition, compound 25 reduces bacterial flagella aggregation, leading to decreased swimming performance [64]. Compound 26 inhibits swimming motility, flagella assembly, and T3SS induced HR. It is further speculated that the RpfF protein in the QS pathway may be its potential target. The results of molecular docking show that the compound is embedded in the pocket of the RpfF protein and binds to Thr A: 243, Ile C: 180, and Cys B: 177 [65]. Compound 27 acts on the bacterial cell membrane and interacts with DNA in a groove binding mode [66]. The results of fluorescence titration indicate that compound 28 has excellent binding affinity with the potential target protein RpfF. Its binding constant (KA) with Xoo RpfF protein is 104.87 L/mol. Molecular docking further demonstrates that compound 28 is embedded in the pocket of the RpfF protein and binds to Glu A: 161, Gly A: 138, Gly A: 86, and Tyr A: 106. In addition, compound 28 can affect the synthesis of DSF signal, biofilm formation, EPS production, cellulase and amylase activities, as well as motility of Xoo, thus achieving antibacterial effects [67]. Compound 29 interferes with the mRNA expression levels of acetyl-CoA carboxylase, acyl carrier protein and fatty acid biosynthesis family genes related to fatty acid synthesis process, reduces the lipid content of membrane, increases the permeability of membrane, and finally destroys the integrity of bacterial membrane [68]. At a concentration of 7.08 µg/mL, the supramolecular nanospheres containing compound 30 exhibited an inhibition rate of 55.85% against the biofilm of Xoo. In addition, this material can alter the membrane permeability of Xoo and disrupt the redox system, leading to the leakage of bacterial electrolytes, oxidative stress and cellular dysfunction, as well as reducing the production of EPS, bacterial motility, extracellular cellulase and amylase activities and bacterial pathogenicity. Moreover, at a concentration of 15 µg/mL, the supramolecular nanospheres containing compound 30 were safe for non-target organisms such as zebrafish and earthworms [69]. The supramolecular antibacterial material containing compound 31 had a significant destructive effect on the mature biofilm of Xac. After the Xac cells had aged for 24 h, at a concentration of 25 µg/mL, the eradication rate was as high as 91.9% [70]. The supramolecular antibacterial material containing compound 32 possessed excellent biofilm eradication ability against Xoo. After the Xoo cells had aged for 48 h, the antibacterial material achieved a biofilm eradication rate of 82.8% at 15.68 µg/mL. Furthermore, this material also inhibited the production of EPS, the transcription of swimming-related genes, bacterial quorum sensing and cellulase and amylase activities [71]. In summary, isopropanolamine compounds can achieve antibacterial effects through various pathways, such as disrupting cell morphology, influencing bacterial DNA, the redox system, the biosynthesis of DSF, biofilm formation, EPS synthesis, QS pathway proteins, T3SS, extracellular enzymes, fatty acid synthesis, and reducing the aggregation of bacterial flagella. In terms of target discovery, phenotypic experiments have suggested that the possible potential targets of compounds 26 and 28 are the RpfF protein in the QS pathway. However, more experiments are needed to verify these targets.
 DownLoad:
CSV
DownLoad:
CSV
| Compd | Bacterial | In vitro antibacterial activities/EC50 (µg/mL) | Protective activities at 200 µg/mL (%) | Mechanism of action | Ref. |
| 21 | Xoo | 1.3 | 48.7 | / | [60] |
| Xac | 2.0 | / | |||
| Psa | 3.8 | / | |||
| 22 | Xoo | 5.0 | 48.2 | / | [61] |
| Xac | 4.7 | / | |||
| Psa | 8.5 | / | |||
| 23 | Xoo | 1.7 | 51.9 | / | [62] |
| Xac | 4.1 | / | |||
| Psa | 2.4 | 65.5 | |||
| 24 | Xoo | 4.8 | 51.8 | / | [63] |
| Xac | 3.2 | / | |||
| 25 | Xoo | 6.8 | 43.6 | Inhibit DSF biosynthesis and related QS pathway gene expression | [64] |
| Xoc | 8.9 | / | |||
| Xac | 3.1 | / | |||
| 26 | Xoo | 1.5 | 48.7 | RpfF | [65] |
| Xac | 2.0 | / | |||
| Psa | 5.7 | / | |||
| 27 | Xoo | 0.8 | 52.0 | Destroy cell morphology, and interacts with DNA in a groove binding mode | [66] |
| Xac | 4.8 | / | |||
| Psa | 2.3 | 80.0 | |||
| 28 | Xoo | 9.9 | 42.6 | RpfF | [67] |
| Xac | 7.0 | ||||
| 29 | Xoo | 1.3 | 43.3 | Interferes with genes involved in fatty acid synthesis and damages the integrity of bacterial cell membranes | [68] |
| Xac | 1.4 | / | |||
| Psa | 8.5 | / | |||
| Cmm | 8.2 | / | |||
| 30 | Xoo | 1.8 | 48.9 | Inhibit the formation of biofilms, alter cell membrane permeability, reduce the production of EPS, and decrease bacterial motility | [69] |
| 31 | Xac | 1.9 | 81.4 | Destruction of cell membrane | [70] |
| 32 | Xoo | 2.0 | 50.8 | Destroy cell membrane and inhibit EPS production | [71] |
| Cmm: Clavibacter michiganense subsp. michiganse. | |||||
In the design of isopropanolamine antibacterial agents, the introduction of small groups on the hydroxyl group of isopropanol will not affect the antibacterial activity to a certain extent, such as using a tetrahydrofuran ring for the hydroxyl group, introducing a halogenated alkane at the hydroxyl group or replacing the hydroxyl group with an amino group. The introduction of thioether or amine group into one end of isopropyl alcohol has good antibacterial activity, among which the amino site is generally a nitrogen-containing heterocyclic ring with better biological activity, while the amino alkyl chain is less active. In addition, the other end of the isopropanolamine can be introduced into the nitrogen-containing heterocyclic ring, and ester groups, oxygen atoms or alkyl chains can also be introduced. The carbon chain of the isopropanolamine can also be introduced with alkyl chains or halogenated alkyl groups. Compounds containing isopropanolamine have S and R isomers, and there is insufficient evidence to show whether there is a difference in activity between the S and R isomers. In addition, when designing isopropanolamine antibacterial agents, we can draw on the isopropanolamine structures with antibacterial properties in medicine or the isopropanolamine structures with antifungal properties in pesticides, which will help us to discover compounds with higher activity more easily.
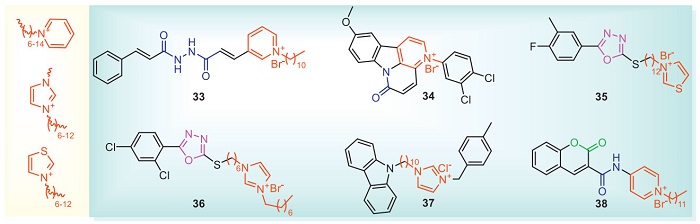
Onium salts are composed of a hydrophilic part and a hydrophobic part. This structure enables the cations to be electrostatically attracted to the negatively charged cell membranes of microorganisms, and then incorporate their hydrophobic chains into the phospholipid molecular layer to interact with the negatively charged phospholipids, thereby achieving the purpose of antibacterial action [72]. The hydrophilicity and lipophilicity of onium salts are one of the templates for the production of biologically active compounds especially for their excellent activity in bacteria and fungi, which has attracted the attention of researchers [73,74]. The structures of onium salts with antibacterial activities are shown in Fig. 5 and Table 4 [75-80]. Compound 33 reduces the guanosine triphosphatase activity of recombinant FtsZ (the filamentous temperature-sensitive protein Z). The results of Western blot assays showed that compound 33 decreased the expression of FtsZ, preventing binary fission. The KA between compound 33 and FtsZ was calculated to be 103.46 L/mol through fluorescence titration. Molecular docking shows that compound 33 had strong interaction with ASN34, GLN193 and GLN197 residues in the α-helix region of FtsZ [75]. At 15.62 µg/mL, the cell viability of compound 34 on BHK-21 cells (Baby hamster Syrian kidney cells) also exceeded 96%, indicating that it has low toxicity to mammalian cells. What is more, at 3.91 µg/mL, compound 34 has no inhibitory effect on the seed germination rate of turnip and wheat, indicating that compound 34 does not inhibit plant growth [76]. At 200 µg/mL, compound 35 has good activity in vivo against rice bacterial leaf blight, and its curative and protective activities are 48.5% and 51.5%, respectively. It was further discovered that the down-regulation of Xoo ribosomal differential proteins triggered by compound 35 disrupts the pathway of ribosome assembly and subsequently synthesizes a large number of functional proteins essential for normal life, eventually leading to the death of bacteria [77]. Compound 36 destroys the complete morphology of cells, and it may be that the protons of the imidazole cation promoted the interaction with the bacterial membrane. It is further speculated that the compound will be protonated and deposited through electrostatic interactions between cell components, and the hydrophobic fragments will penetrate the bacterial membrane, resulting in the chaos of cell membranes and material leakage [78]. Compound 38 increased the activity of aldehyde dehydrogenase ALDH2b, malate dehydrogenase, pyruvate kinase, acetoacetyl-CoA thiolase and lactoylglutathione lyase in the pyruvate metabolic pathway, and enhanced the resistance of rice to bacterial infection [80].
 DownLoad:
CSV
DownLoad:
CSV
| Compd | Bacterial | In vitro antibacterial activities (µg/mL) | Protective activities at 200 µg/mL (%) | Mechanism of action | Ref. |
| 33 | Xoo | EC50 = 1.0 | 63.2 | FtsZ | [75] |
| 34 | Rs | MIC = 3.9 | / | / | [76] |
| Ps | MIC = 3.9 | / | |||
| 35 | Xoo | EC50 = 0.1 | 51.5 | Disrupt the ribosome assembly pathway | [77] |
| Rs | EC50 = 9.6 | / | |||
| Xac | EC50 = 14.6 | / | |||
| 36 | Xoo | EC50 = 0.3 | / | Destroy cell morphology | [78] |
| Rs | EC50 = 0.6 | / | |||
| 37 | Xoo | EC50 = 0.6 | 42.6 | / | [79] |
| 38 | Xoo | EC50 = 0.2 | 55.3 | The activities of POD and phenylalanine ammonia-lyase (PAL) defense enzymes and proteins related to pyruvate metabolic pathway were increased | [80] |
| Xoc | EC50 = 1.1 | / | |||
| Xac | EC50 = 0.8 | / |
When designing an onium salt structure with an antibacterial compound, it is necessary to consider the type of onium salt, the length of the alkyl group, bridging atom, the position of the substituent, and the type of halogen. For the type of onium salt, the introduction of substituted pyridine and imidazole, or the introduction of quaternary amine and phosphonium structures can be considered. The length of the alkyl chain of the onium salt has a great influence on antibacterial activity. When the alkyl chain length is 6–12, the antibacterial activity is usually higher than the alkyl chain length of 0–6.
The quinazoline skeleton consists of two fused six-membered aromatic rings (benzene rings and pyrimidine rings). Due to its excellent biological activity, it has been rapidly developed, and then commercial drugs have been successfully developed, such as the fungicides tebufloquin and quinoxyfen [81-83]. In recent years, the quinazoline skeleton has also been introduced into the field of antibacterial agents. The structures and antibacterial activities of some quinazolines with antibacterial activities are shown in Fig. 6 and Table 5 [84-92]. The ABPP experiment was used to label the specific proteins of compound 39. After identification, it was determined that its target in Xoo is β-ketoacyl-ACP synthase Ⅱ (FabF). Molecular docking showed that 39 covalently bound to the Cys151 residue of FabF. Furthermore, through physiological and biochemical experiments, it was demonstrated that 39 inhibited the biosynthesis of fatty acids and disrupted the integrity of cell membranes [84]. Proteomics showed that compound 41 may affect the metabolic pathways of D-glucose and biotin. It is further speculated that compound 41 can regulate and destroy the D-glucose and biotin synthesis pathways, affect the energy metabolism in Xoo, and cause the death of bacteria. Furthermore, compound 41 also can destroy the integrity of bacteria and increase membrane permeability [86]. Compound 44 disrupts bacterial morphology and induces ROS accumulation in Xac. Further proteomic results show that 44 acts on the T2SS and T4SS secretion system of Xac [89]. Compound 47 can reduce the swimming motility of Xoc to exert its antibacterial effect, and further proteomic also shows that 47 can down-regulate the protein related to the biosynthesis of bacterial flagella and reduce the motility of flagella [92]. In summary, quinazoline compounds may exert antibacterial effects through multiple pathways, including inhibiting fatty acid synthesis, affecting the synthesis and transport of ribosomal proteins, influencing proteins associated with bacterial flagellar biosynthesis, interfering with energy metabolism pathways, disrupting the T2SS and T4SS, suppressing the formation of EPS, altering membrane permeability, and destroying bacterial morphology.
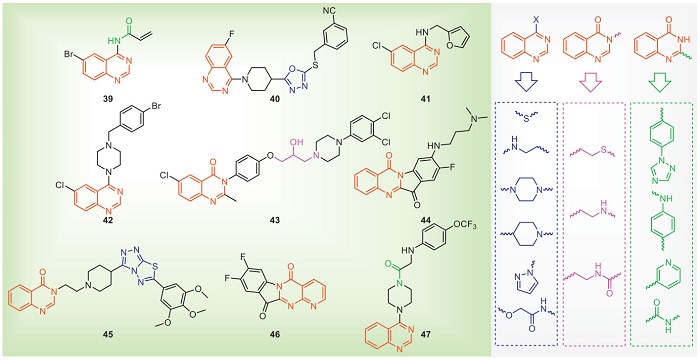 DownLoad:
CSV
DownLoad:
CSV
| Compd | Bacterial | In vitro antibacterial activities/EC50 (µg/mL) | Protective activities at 200 µg/mL (% | Mechanism of action | Ref. |
| 39 | Xoo | 13.2 µmol/L | 49.6 | FabF | [84] |
| 40 | Xoo | 26.0 | / | / | [85] |
| 41 | Xoo | 7.1 | / | Affect the metabolic pathways of D-glucose and biotin | [86] |
| Xac | 18.6 | / | |||
| 42 | Rs | 2.7 | / | Destructive bacterial morphology | [87] |
| 43 | Xoo | 1.5 | 46.7 | Affects ribosomal protein synthesis and transport, energy metabolism pathway and glutathione metabolism disorders | [88] |
| 44 | Xac | 0.8 | / | Interferes with the T2SS and T4SS secretory systems of Xac | [89] |
| Xoo | 1.3 | / | |||
| Psa | 15.5 | / | |||
| 45 | Xoc | 19.5 | / | Inhibit the formation of EPS, change the permeability of membrane and destroy the morphology of bacteria | [90] |
| Xoo | 4.1 | 47.6 | |||
| 46 | Xac | 0.3 | / | Interferes with the T4SS secretory systems of Xac | [91] |
| Xoo | 1.9 | / | |||
| Psa | 18.0 | / | |||
| 47 | Xoc | 7.1 | 50.5 | Down-regulating proteins associated with bacterial flagellar biosynthesis | [92] |
In the structural design of quinazoline with antibacterial compounds, the structural modification of quinazoline is usually at the substituents of the benzene ring and the 2-, 3- and 4-positions. The introduction of a benzene ring, aniline, pyridine or amide structure at the 2-position of quinazoline has no apparent effect on the antibacterial activity. The 2-position of quinazoline is not considered to be necessary to affect the activity. Similarly, introducing a benzene ring at the 3-position of quinazoline into a sulfur-containing, nitrogen-containing or amide-containing alkyl chain has no obvious effect on the antibacterial activity. The antibacterial activities increase after introducing a nitrogen-containing alkyl chain or piperazine at the 4-position of quinazoline. The substituents on the benzene ring of quinazoline are also more obvious to the antibacterial activity, and the introduction of some small groups may increase the activity.
Flavonoid compounds exist in the leaves, stems, roots, flowers and seeds of almost all plants. They have a wide range of biological activities. For bacteria, they can achieve antibacterial effects by means of inhibiting energy metabolism, destroying the cytoplasmic membrane and inhibiting nucleic acid synthesis, etc. [93,94]. With regard to plant bacterial diseases, the structures and antibacterial activities of some representative flavonoids are shown in Fig. 7 and Table 6 [95-100]. Current flavonoid research primarily focuses on compounds isolated from natural products. There is relatively little literature on the structural modification of flavonoids, and the research on their modes of action is extremely scarce. Compound 51 enhances the activities of catalase (CAT) in rice, thereby increasing the anti-Xoo ability. Compounds 48–53 cause the bacteria to deform and even rupture. Whether flavonoids can inhibit the energy metabolism and nucleic acid synthesis of plant bacteria to achieve antibacterial effects, as well as the search for their targets, remains to be further investigated by researchers.
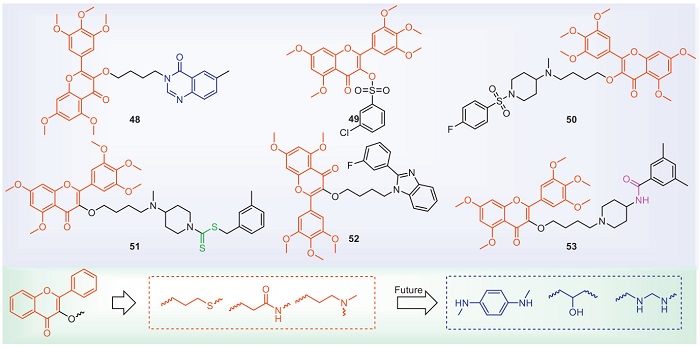 DownLoad:
CSV
DownLoad:
CSV
| Compd | Bacterial | In vitro antibacterial activities/ EC50 (µg/mL) | Protective activities at 200 µg/mL (%) | Mechanism of | Ref. |
| 48 | Psa | 28.1 | / | Destroy cell morphology | [95] |
| Xac | 16.9 | / | |||
| Xoo | 20.4 | / | |||
| 49 | Xoo | 11.9 | 47.0 | [96] | |
| Rs | 17.1 | / | |||
| 50 | Xoo | 4.7 | 63.3 | [97] | |
| 51 | Xoo | 1.6 | 60.1 | [98] | |
| 52 | Xoo | 8.2 | 48.6 | [99] | |
| 53 | Xoo | 2.7 | 64.2 | [100] | |
| Xac | 7.1 | / |
At present, the structural modification of flavonoids is mainly on the hydroxyl group at the 3-position of flavonoids. Introducing phosphate or sulfonate to improve antibacterial activity. The reason may be that the structure of flavonoids is relatively large and can be modified with alkyl chains. Alkyl chains containing amino groups have more advantages in antibacterial activity, which may be due to increased solubility. We suggest introducing isopropanolamine or ethylenediamine as bridge chains. In addition, we need to consider that the solubility of flavonoids is poor, and the bioavailability is not high. Retaining the hydroxyl groups on the flavonoids may improve antibacterial activity.
Amide derivatives have achieved great success in pesticides [101,102]. For example, among bisamide insecticides, flubendiamide and chlorantraniliprole act as ryanodine receptor modulators, while cytochrome bc1 complex inhibitors include pyraclostrobin and kresoxim-methyl. In recent years, many amide derivatives have been designed for the study of antibacterial activities. For example, the commercial drug niclosamide hinders the formation of biofilms by disrupting membrane permeability, and significantly affects xanthoside, EPS synthesis and quorum sensing signals in Xoo [103]. Fig. 8 and Table 7 show antibacterial activities of some representative amide compounds [104-118]. At 10 µg/mL, the inhibitory effect of compound 54 on hpa1 promoter activity is 98.05%. What is more, compound 54 reduces hypersensitive response (HR) of Xoo without affecting the growth or survival of bacteria [104]. At 75 g a.i./ha, the control effect of compound 55 against Ps (Pseudomonas syringae) was 39.2%. Furthermore, the control effect of compound 55 (35 g a.i./ha), prothioconazole (50 g a.i./ha) and tebuconazole (50 g a.i./ha) can reach 59% after being mixed [105]. The MIC of compound 57 designed and synthesized with FabH as the target on Xoo KACC10331 is 0.49 µg/mL, while compound 57 showed no inhibitory effect on FabH-deficient mutants, indicating that FabH can be used as a target for compound design [107]. Proteomic results show that compound 59 affects glyoxylate and dicarboxylate metabolism and galactose metabolism in rice to achieve the purpose of disease resistance [109]. Based on the excellent antibacterial activity of compound 64, 35 specific target proteins were screened by ABPP. Western blot assay and enzyme activity test identified the target as pyruvate kinase. Transcriptomic and pathogenicity test showed that the deletion of pyruvate kinase gene caused the downregulation of genes related to the flagellar assembly and two-component regulatory system pathway of Xoo, and weakened the flagellar formation ability, swimming ability and infection ability of rice. The target proteins interacting with pyruvate kinase protein were screened by Co-IP method. Finally, the interacting protein Clp (CRP-like protein Clp, Pxo_04006) and RegR (Two-component system regulatory protein, Pxo_00797) of pyruvate kinase were screened by bacterial two-hybrid and GST pull-down methods [114]. By ABPP and mass spectrometry, it was found that the target protein of compound 65 for Xoo was Clp. The microscale thermophoresis (MST) test showed that compound 65 had a high affinity with Clp protein, and the Kd value was 0.52 µmol/L. The binding value constant of compound 65 and Clp protein was positively correlated with the antibacterial effect. In addition, the results of qRT-PCR showed that compound 65 could inhibit the expression of Clp gene in Xoo. The results of pathogenicity tests indicated that compound 65 treatment and Clp gene deletion equally reduced the pathogenicity of Xoo. The results showed that compound 65 inhibited the expression of Clp gene in Xoo by binding to Clp protein [115]. The target of compound 67 on Xoo was identified by drug affinity responsive target stability (DARTS) method as 50S ribosomal protein L2. In addition, the excellent in vitro interaction between compound 67 and 50S ribosomal protein L2 was demonstrated by MST and fluorescence quenching spectra [117].
 DownLoad:
CSV
DownLoad:
CSV
| Compd | Bacterial | In vitro antibacterial activities | In vivo antibacterial activities | Mechanism of action | Ref. |
| 54 | Xoo | 98.05% inhibitors of Xoo T3SS at 10 µg/mL | / | Inhibit T3SS | [104] |
| 55 | Ps | / | 39.2% at 75 g a.i./ha | / | [105] |
| 56 | Xoo | EC50 = 11.6 µg/mL | 47.0% at 200 µg/mL | / | [106] |
| 57 | Xoo | MIC = 0.5 µg/mL | / | FabH | [107] |
| 58 | Erwinia carotorora | EC50 = 2.7 µg/mL | / | / | [108] |
| 59 | Xoo | EC50 = 4.2 µg/mL | 53.7% at 200 µg/mL | Affects glyoxylate and dicarboxylate metabolism and galactose metabolism in rice | [109] |
| 60 | Xoo | EC50 = 28.6 µg/mL | 51.3% at 200 µg/mL | Destroy cell morphology | [110] |
| Rs | EC50 = 55.3 µg/mL | / | |||
| 61 | Cmm | EC50 = 4.1 µg/mL | 71.8% at 100 µg/mL | Causes peroxidation damage and leakage of electrolytes and proteins from the bacterial cells | [111] |
| 62 | Xoo | EC50 = 5.7 µg/mL | 55.7% at 200 µg/mL | Disrupt the balance of the redox system | [112] |
| Xac | EC50 = 6.6 µg/mL | / | |||
| Psa | EC50 = 9.1 µg/mL | ||||
| 63 | Xcc | 77.5% inhibitors of Xcc T3SS at 20 µg/mL | / | Inhibit T3SS | [113] |
| 64 | Xoo | EC50 = 0.4 µg/mL | 50.0% at 200 µg/mL | Pyruvate kinase | [114] |
| Xoc | EC50 = 3.1 µg/mL | / | |||
| Xac | EC50 = 2.9 µg/mL | ||||
| 65 | Xoo | EC50 = 0.6 µg/mL | 45.3% at 200 µg/mL | Clp | [115] |
| Xoc | EC50 = 0.8 µg/mL | 51.7% at 200 µg/mL | |||
| 66 | Xoo | EC50 = 1.1 µg/mL | / | Up-regulated expression of genes OsBIPP2C1, OsPYL/RCAR5 and OsABF1 activated ABA signaling pathway | [116] |
| Xoc | EC50 = 0.9 µg/mL | 70.3% at 200 µg/mL | |||
| Xac | EC50 = 1.8 µg/mL | 64.4% at 200 µg/mL | |||
| 67 | Xoo | EC50 = 5.1 µg/mL | / | 50S ribosomal protein L2 | [117] |
| Xac | EC50 = 5.2 µg/mL | ||||
| 68 | Rs | / | / | Fucose-binding protein (PDB ID: 4CSD) | [118] |
In the design of the amide structure with antibacterial compounds, in addition to the aromatic ring, one end of the carbonyl group in the amide can also be connected to an alkyl chain or heteroatoms, such as introducing Schiff bases and thioethers. Using amide as a bridge, introducing alkenyl, cyclopropane, or changing the amide to thioamide may improve the antibacterial activity. When we design antibacterial compounds, the introduction of amide backbone is often beneficial to improve antibacterial activity in vivo.
The structure containing esters is an important skeleton in the development of pesticides. At present, pyrethroid insecticides, carbamates insecticides, methoxyacrylates, and carbamate fungicides have been successfully developed. The phenolic acid structure has excellent antibacterial activity in nature, and the further derivatization of the leading structure based on phenolic acid into ester has attracted the attention of researchers. In recent years, many ester derivatives have been designed to study their antibacterial activities. Fig. 9 and Table 8 show the antibacterial structures of some representative ester compounds [119-126]. Compounds 69–71 and 73 can reduce the mRNA level of hrpG and hrpX. Moreover, it was found that compounds 69–71 and 73 can affect the mRNA level of hrcT by a fluorescence-activated cell sorting system. It indicates that these compounds may play a role in the regulatory pathway to affect the expression of the T3SS gene, possibly through the HrpX-HrpG pathway to reduce the expression of hrp/hrc genes [119-121,123]. At 200 µg/mL, compound 72 has excellent in vivo antibacterial activities against rice bacterial leaf blight, and its curative and protective activities are 40.9% and 48.9%, respectively. What's more, compound 72 can regulate the oxidative phosphorylation pathway of rice, upregulate cytochrome c oxidase subunit 6b, Os08g0556600 protein, ETC complex I subunit conserved region family protein differential protein, and enhance rice disease resistance [122].
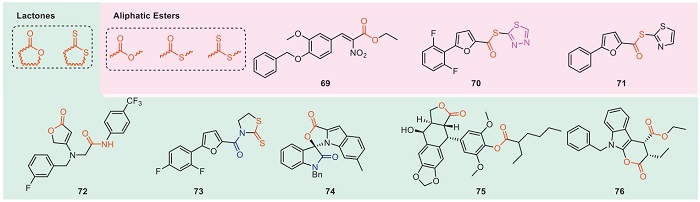 DownLoad:
CSV
DownLoad:
CSV
| Compd | Bacterial | Concentration | Antibacterial activities | Mechanism of action | Ref. |
| 69 | Xoo | 100 mmol/L | 99.0% inhibitors of Xoo T3SS | Inhibit T3SS | [119] |
| 70 | Xoo | 200 µg/mL | 58.1% | [120] | |
| 71 | Xoo | 200 µg/mL | 63.8% | [121] | |
| 72 | Xoo | / | EC50 = 35.8 µg/mL | Regulate the oxidative phosphorylation pathway of rice | [122] |
| 73 | Xoo | 10 µg/mL | 65.2% inhibitors of Xoo T3SS | Inhibit T3SS | [123] |
| 74 | Rs | 50 µg/mL | 88.2% | [124] | |
| 75 | Xoo | 100 µmol/L | 48.6% (the FtsZ GTPase activity) | FtsZ | [125] |
| 76 | Xoo | / | EC50 = 5.8 µg/mL | DHPS (dihydropteroate synthase) | [126] |
According to the differences in physical and chemical properties, esters are classified into lactones and aliphatic esters. When designing lactone structures with antibacterial activity, natural product structures such as coumarins and terpene-containing lactones can be considered for introduction. Introducing conjugated groups such as alkenyl or aromatic rings to the carbonyl moiety of the ester group may help enhance antibacterial activity.
When designing aliphatic ester structures with antibacterial activity, phenolic acids can be introduced into the acid moiety, as they play a key role in initiating the symbiotic relationship between plants and beneficial microorganisms. Additionally, phenolic compounds exhibit broad-spectrum antibacterial activity due to their ability to disrupt cell membrane integrity and decouple respiratory proton gradients [127].
The successful development of commercial agents such as thiazole copper, oxine-copper, copper abietate, zinc thiazole encouraged countless researchers to study the antibacterial activity of metal complexes [128-130]. The antibacterial mechanism of metal is usually achieved by interfering with the expression of various bacterial virulence factors, affecting the signal transduction involving the metal sensing system and the metal absorption system to achieve antibacterial. In addition, metals can also protect plants from infection [16]. Metals can be used alone or in the form of inorganic salts for antibacterial activity studies, or as complexes. At present, the research of metal complexes with antibacterial activity mainly focuses on metal ions such as copper, zinc, silver, manganese, nickel, tin, cobalt, selenium and magnesium [131-138]. Kouris et al. complex cobalt with 2,2′-bipyridine, 1, 10-phenanthroline, and 2,2′-bipyridylamine. The SAR shows that the cobalt complex of 1,10-phenanthroline shows excellent antibacterial activity against X. campestris, and its MIC is 0.125 µg/mL [131]. Naz et al. complexed 2-phenylbutyric acid derivatives with organotin, and obtained products with excellent antibacterial activity. The MIC for Xanthomonas vesicatoria and Erwinia carotovora can reach 0.0001 µg/mL [132]. Chen et al. combined selenium and lenized polylactic acid as Se@PLA, which showed excellent antibacterial activity against Xoo with an EC50 value of 13.4 µg/mL. The results of acute toxicity showed that Se@PLA did not cause earthworm death at 3400 µg/g, 14 days after treatment. In addition, at 24 h, the LC50 value for zebrafish was 3836.5 µg/mL. The mechanism of action showed that Se@PLA destroyed the complete morphology of Xoo cells and induced the overproduction of ROS [138]. When designing metal complexes, binding metal ions in the form of phenolic acids or alkaloids can be more considered.
Some structures with good antibacterial activity cannot be classified according to the active framework because of the few articles. Fig. 10 and Table 9 show some representative structures [139-153]. Chen et al. modified acetophenones and synthesized α-haloketone compound 77. The EC50 values of anti-Xoo and Xac were 0.09 and 0.16 µg/mL, respectively. The type of halogen atom at the α-position has a significant effect on the activity. At 50 µg/mL, compound 77 has excellent in vivo antibacterial activity against rice bacterial leaf blight, and its curative and protective activities are 48.9% and 52.1%, respectively [139]. Liu et al. modified the insecticide triflumezopyrim structure and found that the mesoionic compound 78 also has good antibacterial activity. At 200 µg/mL, the curative and protective activities of compound 78 against bacterial leaf blight were 72.04% and 75.12%, respectively [140]. Based on the structure of mesoionic compound 78, Liu et al. further modified the structure to obtain mesoionic compound 79. At 200 µg/mL, the curative and protective activities of compound 79 against rice bacterial leaf blight and bacterial leaf streak are 38.2%, 49.2% and 35.1%, 39.7%, respectively. Further, compound 79 is found to enhance SOD, PAL, CAT and oxidative phosphorylation pathways in rice, thereby improving disease resistance [141]. It shows that the mesoionic compound is a potential antibacterial agent structure. Yu et al. used antidesmone and maltol as leading structures to synthesize compound 80 with antibacterial activity. At 200 µg/mL, the curative and protective activities of compound 80 against bacterial leaf blight were 52.42% and 52.70%, respectively. It shows that pyridine-4(1H)-one may be a skeleton with antibacterial activity [142]. Sun et al. modified the structure of dehydrozingerone to obtain excellent antibacterial compounds 81 and 82. At 200 µg/mL, the protective activities of compounds 81 and 82 against rice bacterial blight can reach 51.7% and 57.4%. It shows that introducing the trifluoromethyl group at the α position of the carbonyl group can improve the antibacterial activity [143,144]. Li et al. modified the structure of the broad-spectrum antibiotic yanglingmycin and introduced a sulfur atom into the oxazole ring to obtain compound 83. The MICs of Rs and Pseudomonas syringae pv. actinidiae (Psa) were 3.91 and 7.81 µg/mL, respectively. Based on the relationship between structure and activity, oxazole rings containing hydroxyl groups are presumed to be the backbone of antibacterial activity [145]. Wu et al. found that compound 84 increased the SOD and CAT defense enzyme activities in rice, in addition, proteomics explains that compound 84 is resistant to Xoo invasion by upregating the photosynthetic pathway ATP synthase subunit d and the NADH dehydrogenase [ubiquinone] iron-sulfur 6 [146]. At a concentration of 10 µg/mL, compound 89 can inhibit the hpa1 of the T3SS at an inhibition rate of up to 86.4%, and it does not affect the growth of bacteria. In addition, the compound 89 can also effectively reduce the HR caused by Xoo in tobacco and decrease the pathogenicity of Xoo in rice [151]. The multifunctional supramolecular material containing compound 91 can effectively reduce the transcription of hrpG, hrpX, hrpE and hrpF in the T3SS, thereby resulting in the inhibition of the pathogenic ability of Xoo [153].
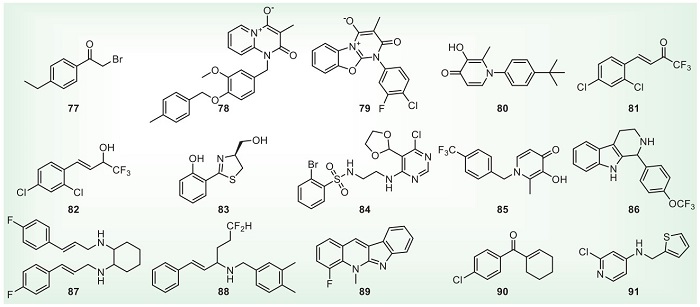 DownLoad:
CSV
DownLoad:
CSV
| Compounds | Bacterial | In vitro antibacterial activities (µg/mL) | Protective activities at 200 µg/mL (%) | Mechanism of action | Ref. |
| 77 | Xoo | EC50 = 0.1 | 52.1 at 50 µg/mL | Inhibit the formation of extracellular polysaccharide and biofilm of Xoo, and change the permeability of cell membrane | [139] |
| Xac | EC50 = 0.2 | ||||
| 78 | Xoo | EC50 = 1.1 | 75.1 | / | [140] |
| Xac | EC50 = 12.4 | ||||
| 79 | Xoo | EC50 = 36.8 | 49.2 | Enhance SOD, PAL, CAT and oxidative phosphorylation pathways in rice | [141] |
| Xoc | EC50 = 47.6 | 39.7 | |||
| 80 | Xoo | EC50 = 26.4 | / | / | [142] |
| 81 | Xoo | EC50 = 11.4 | 51.7 | / | [143] |
| 82 | Xoo | EC50 = 8.3 | 57.4 | / | [144] |
| Xoc | EC50 = 25.9 | / | |||
| 83 | Rs | MIC = 3.9 | / | / | [145] |
| Psa | MIC = 7.8 | ||||
| 84 | Xoo | EC50 = 26.7 | 45.1 | Increased POD and CAT in rice, upregulated ATP synthase subunit d and NADH dehydrogenase iron-sulfur 6 | [146] |
| Xoc | EC50 = 30.8 | 43.7 | |||
| Xac | EC50 = 65.4 | ||||
| 85 | Xoo | EC50 = 10.0 | 49.2 | Inhibition of extracellular enzymes, EPS, biofilm, T3SS effector virulence factors | [147] |
| Xac | EC50 = 13.3 | ||||
| 86 | Xoo | EC50 = 7.3 | 52.7 | Destroy the redox system of Xoo, causing ROS accumulation | [148] |
| Xac | EC50 = 4.9 | ||||
| 87 | Xoo | MIC = 3.1 | 55.9 | Destroy cell morphology and inhibit the production of EPS | [149] |
| Xac | MIC = 3.1 | ||||
| 88 | Xoo | EC50 = 6.6 | 37.5 | Destroy cell morphology | [150] |
| 89 | Xoo | 83.4% inhibitors of hpa1 at 10 µg/mL | / | Inhibit the transcription of hpa1, hrpG, hrpX and hrcT in the T3SS | [151] |
| 90 | Xoo | 84.8% at 100 µg/mL | / | / | [152] |
| 91 | Xoo | EC50 = 9.4 µg/mL | 48.1 | Inhibit the transcription of hrpG, hrpX, hrpE and hrpF in the T3SS | [153] |
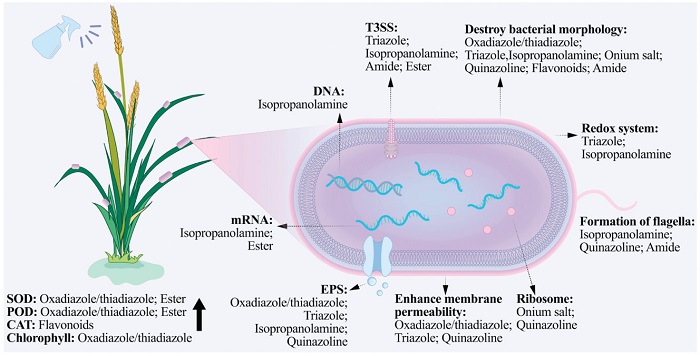
Plant bacterial diseases have caused substantial economic losses to agriculture worldwide and have seriously threatened the sustainable development of modern agriculture. Chemical control remains the most effective control method in the future. However, there are currently significant problems such as outdated varieties of agricultural antibacterial agents, low control efficacy, enhanced bacterial resistance, excessive pesticide residues, and a shortage of alternative varieties. To develop new, efficient and safe agricultural antibacterial agents, this article classifies antibacterial agrochemicals into oxadiazole/thiadiazole, triazole, isopropanolamine, onium salt, quinazoline, flavonoids, amide, ester, metal complex and others according to their active skeletons, and reviews the research on the SAR and action mechanisms of agricultural antibacterial agents in recent years. Summary the mechanisms of action of antibacterial agrochemicals was shown in Fig. 11. Oxadiazole/thiadiazole compounds can not only enhance the activities of SOD, POD and chlorophyll in plants but also enhance various regulatory pathways, thereby enhancing resistance to bacteria. In addition, oxadiazole/thiadiazole compounds destroy the morphology of bacteria, enhance the permeability of cell membranes, and achieve the purpose of antibacterial. Exciting results have also been achieved in the discovery of targets. Song et al. found through proteomics that compound 4 affects the motility and virulence of Xoc. Subsequently, the ABPP technique was further employed to screen out the target proteins CsrA and Xoc3530. Moreover, using the ABPP technique, the target protein DLST of Xoo and Xac was also identified for compound 1. Triazole compounds also disrupt the morphology of bacteria and enhance the permeability of cell membranes. What is more, triazoles may downregulate the bacterial type Ⅲ secretion system and transcription activator-like effector related proteins, and interfere with bacterial redox system. However, the understanding of the mechanism of action of triazole compounds is mainly based on the phenotypic analysis of pathogenic bacteria and the detection of physiological and biochemical indicators. Their action targets and regulatory mechanisms remain unclear. Isopropanolamine compounds can achieve antibacterial effects through various pathways, such as disrupting cell morphology, influencing bacterial DNA, the redox system, the biosynthesis of DSF, biofilm formation, EPS synthesis, QS pathway proteins, T3SS, extracellular enzymes, fatty acid synthesis, and reducing the aggregation of bacterial flagella. In terms of target discovery, phenotypic experiments have suggested that the possible potential targets of compounds 26 and 28 are the RpfF protein in the QS pathway. Onium salts are electrostatically attracted to the negatively-charged bacterial cell membranes. Then, their hydrophobic chains interact with the negatively-charged phospholipids, thereby disrupting the bacterial morphology. Additionally, onium salts may also affect bacterial FtsZ and ribosomal proteins. In terms of target discovery, based on the approach of similar structures having similar mechanisms and the determination of GTPase activity, it was found that the target of compound 33 is FtsZ. The target was further verified through phenotypic observation, binding constant testing, circular dichroism, and Western blot. Quinazoline compounds may exert antibacterial effects through multiple pathways, including inhibiting fatty acid synthesis, affecting the synthesis and transport of ribosomal proteins, influencing proteins associated with bacterial flagellar biosynthesis, interfering with energy metabolism pathways, disrupting the T2SS and T4SS, suppressing the formation of EPS, altering membrane permeability, and destroying bacterial morphology. In terms of target discovery, the ABPP technique was used to identify that the target of compound 39 against Xoo is FabF. The results of molecular docking show that compound 39 binds covalently to the Cys151 site in FabF. Flavonoid compounds disrupt the morphology of bacteria and enhance the CAT activity of the host. However, research on their mechanism of action is limited. It remains unknown whether these compounds can inhibit the energy metabolism, nucleic acid synthesis, and other pathways of plant bacteria. Moreover, the targets of flavonoids against plant bacterial diseases have not been discovered yet. Amide compounds have different antibacterial mechanisms. For example, compound 54 can reduce the HR of Xoo without affecting bacterial growth, while compound 61 can disrupt the bacterial cell morphology, leading to the leakage of electrolytes and proteins, thus killing the bacteria. In terms of targets, Song et al. used the ABPP technique to screen out that the target of compound 64 against Xoo is pyruvate kinase, and verified the target through Western blot and enzyme activity assays. The Co-IP method, bacterial two-hybrid assay, and GST pull down assay were used to screen the interacting proteins of pyruvate kinase, which were identified as Clp and RegR. Similarly, Song et al. used the ABPP technique to screen out that the target of compound 65 against Xoo is Clp, and verified it through MST experiments, qRT-PCR, and experiments on constructing knockout mutants. Yang et al. used the DARTS method to screen out that the target of compound 67 acting on Xoo is the 50S ribosomal protein L2, which was confirmed by MST and fluorescence quenching spectroscopy. Moreover, compound 57, which was designed and synthesized based on FabH as the target, exhibits excellent antibacterial activity against Xoo. Ester compounds may affect the expression of bacterial T3SS genes, thereby reducing virulence. Compounds 75 and 76 with excellent antibacterial activity were virtually screened based on the targets FtsZ and DHPS. Metal complexes not only interfere with a variety of bacterial virulence factors, affect signal transduction, involve metal sensing systems and metal uptake systems, but also protect plants from infection.
Despite the rapid development of antibacterial agrochemicals at present, there are still some issues that need further research and exploration in the future:
(1) Currently, there is a lack of structural diversity in commercially available agricultural antibacterial agents, which restricts the further development of structural modification of agricultural antibacterial agents. To continue the search for promising antibacterial active skeletons, we can draw on the active skeletons of antibacterial drugs, veterinary drugs, or natural products with antibacterial activity as lead compounds for structural modification. Through high-throughput screening technology, lead compounds with novel structures and unique action mechanisms can be screened. In addition, Artificial Intelligence-assisted screening of new, efficient and low-toxicity agricultural antibacterial agents is also a future development trend.
(2) Developing the structures of agricultural antibacterial agents based on targets. Currently, potential targets that have been discovered include pyruvate kinase, DLST, CsrA, Xoc3530, RpfF, FabF, 50S ribosomal protein L2, fucose-binding protein, FtsZ, Clp and XooDHPS, etc. However, there is a lack of research on target verification and regulatory pathways, as well as the screening of agricultural antibacterial agents targeting these targets. In addition, virulence factors such as the type Ⅲ secretion system that do not affect bacterial growth are ideal targets. However, the targets and action mechanisms of most inhibitors are still unclear at present, and there is a lack of reports on field applications and registrations. This prompts us to further verify the targets, analyze the structures and functions of the targets, and with the aid of computer-aided design platforms, use methods such as molecular docking and virtual screening to screen antibacterial agent structures that can bind to the targets and possess activity from the compound database.
(3) Currently, research on the antibacterial mechanisms of compounds is mainly limited to observing the phenotypic changes of pathogenic bacteria, physiological and biochemical indicators, and the regulatory pathways related to the pathogenicity/resistance of bacteria or plants through proteomic or transcriptomic experiments; predicting the targets of combined effects by detecting changes in pathogenic factors and gene expression. However, there is a lack of more in-depth explanations for the target proteins of bacteria or plants and their regulatory functions. Fortunately, with the continuous progress of molecular biology techniques, we can use techniques such as ABPP, drug affinity responsive target stability (DART), cellular thermal shift assay (CETSA), combinatorial functional genomics, proteomics, structural biology, bioinformatics, and big data to further explore the target proteins of highly active molecules. Identifying the targets of antibacterial compounds can guide the design of new antibacterial compounds and lay the foundation for solving the problem of antibacterial agent resistance and the research and development of multi-targeting.
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Shaobo Wang: Writing – review & editing, Writing – original draft, Validation. Honggui Yang: Writing – original draft. Chunni Zhao: Writing – original draft. Deyu Hu: Writing – review & editing, Writing – original draft, Validation, Supervision. Baoan Song: Writing – review & editing, Writing – original draft, Supervision, Project administration, Methodology, Investigation, Funding acquisition, Formal analysis, Data curation.
The financial support from the National Natural Science Foundation of China (No. 31972290), and National Key Research and Development Program of China (No. 2022YFD1700300).
G.W. Sundin, L.F. Castiblanco, X.C. Yuan, et al., Mol. Plant Pathol. 17 (2016) 1506–1518. doi: 10.1111/mpp.12436
J. Mansfield, S. Genin, S. Magori, et al., Mol. Plant Pathol. 13 (2012) 614–629. doi: 10.1111/j.1364-3703.2012.00804.x
M. Srilatha, N. Patyal, M.S. Saddala, J. Integr. Agric. 19 (2020) 735–747. doi: 10.1016/S2095-3119(19)62813-3
D.H. Zheng, H.H. Wang, H. Zhong, et al., mSystems 6 (2021) e00789.
S. Timilsina, N. Potnis, E.A. Newberry, et al., Nat. Rev. Microbiol. 18 (2020) 415–427. doi: 10.1038/s41579-020-0361-8
W.D. Liu, J.L. Liu, L. Triplett, J.E. Leach, G.L. Wang, Annu. Rev. Phytopathol. 52 (2014) 213–241. doi: 10.1146/annurev-phyto-102313-045926
V. Verdier, C.V. Cruz, J.E. Leach, J. Biotechnol. 159 (2012) 320–328. doi: 10.1016/j.jbiotec.2011.09.020
D.O. Niño-Liu, P.C. Ronald, A.J. Bogdanove, Mol. Plant Pathol. 7 (2006) 303–324. doi: 10.1111/j.1364-3703.2006.00344.x
S. Genin, New Phytol. 187 (2010) 920–928. doi: 10.1111/j.1469-8137.2010.03397.x
S.M. Im, N.H. Yu, H.W. Joen, et al., Pestic. Biochem. Phys. 163 (2020) 130–137. doi: 10.1016/j.pestbp.2019.11.007
F. Mourgues, M.N. Brisset, E. Chevreau, Trends Biotechnol. 16 (1998) 203–210. doi: 10.1016/S0167-7799(98)01189-5
A.C. Udayashankar Chithrashree, S. Chandra-Nayaka, M.S. Reddy, C. Srinivas, Biol. Control 59 (2011) 114–122. doi: 10.1016/j.biocontrol.2011.06.010
D. Peng, K. Luo, H.D. Jiang, et al., Pest Manag. Sci. 73 (2017) 1253–1257. doi: 10.1002/ps.4453
B.J. Jacobsen, N.K. Zidack, B.J. Larson, Phytopathology 94 (2004) 1272–1275. doi: 10.1094/PHYTO.2004.94.11.1272
A. Cameron, V. Sarojini, Plant Pathol. 63 (2014) 1–11. doi: 10.1111/ppa.12066
H. Fones, G.M. Preston, FEMS Microbiol. Rev. 37 (2013) 495–519. doi: 10.1111/1574-6976.12004
K. Griffin, C. Gambley, P. Brown, Y.J. Li, Crop Prot. 96 (2017) 144–150. doi: 10.1016/j.cropro.2017.02.008
F. Behlau, J.C. Hong, J.B. Jones, J.H. Graham, Phytopathology 103 (2013) 409–418. doi: 10.1094/PHYTO-06-12-0134-R
R. Chai, W.M. Rooney, J.J. Milner, D. Walker, Pest Manag. Sci. 77 (2021) 3836–3840. doi: 10.1002/ps.6312
V.O. Stockwell, D. Duffy, Rev. Sci. Tech. Off. int. Epiz. 31 (2012) 199–210. doi: 10.20506/rst.31.1.2104
G.W. Sundin, N. Wang, Annu. Rev. Phytopathol. 56 (2018) 161–180. doi: 10.1146/annurev-phyto-080417-045946
G. Bantleon, C. Scheer, K. Hartung, R.T. Voegele, J. Plant Pathol. 103 (2021) 155–165. doi: 10.1007/s42161-020-00694-0
L.L. He, L.T. Xiong, X. Wang, et al., Chin. Chem. Lett. 36 (2025) 110044. doi: 10.1016/j.cclet.2024.110044
N.T. Vu, C.S. Oh, Plant Pathol. J. 36 (2020) 204–217. doi: 10.5423/ppj.rw.04.2020.0074
G.L. Vanti, M. Kurjogi, K.N. Basavesha, et al., J. Biotechnol. 309 (2020) 20–28. doi: 10.1016/j.jbiotec.2019.12.009
Y.D. Li, Y.L. Liu, D.S. Yang, et al., J. Hazard. Mater. 394 (2020) 122551. doi: 10.1016/j.jhazmat.2020.122551
I. Güell, S. Vilà, E. Badosa, et al., Peptide Sci. 108 (2017) e23012. doi: 10.1002/bip.23012
A. Cameron, G.H.D. Zoysa, V. Sarojini, Biopolymers 102 (2014) 88–96. doi: 10.1002/bip.22423
J.F. Niu, D.Y. Nie, D.Y. Yu, et al., Pestic. Biochem. Phys. 143 (2017) 8–13. doi: 10.1016/j.pestbp.2017.10.004
H.T. Nguyen, H.G. Kim, N.H. Yu, et al., J. Agric. Food Chem. 69 (2021) 5471–5480. doi: 10.1021/acs.jafc.1c01162
D.Q. Pham, D.T. Ba, N.T. Dao, et al., Ind. Crop. Prod. 108 (2017) 442–450. doi: 10.1016/j.indcrop.2017.06.067
X.L. Su, S. Xu, Y. Shan, et al., Fitoterapia 127 (2018) 186–192. doi: 10.1016/j.fitote.2018.02.003
X.Y. Liang, X.Y. Yu, X.Y. Pan, et al., Mol. Plant Pathol. 19 (2018) 116–128. doi: 10.1111/mpp.12503
X.Y. Liang, Y.B. Duan, X.Y. Yu, et al., Pest Manag. Sci. 72 (2016) 997–1003. doi: 10.1002/ps.4080
L. Shi, P. Li, W.L. Wang, et al., Molecules 20 (2015) 11660–11675. doi: 10.3390/molecules200711660
B. Chen, Q.S. Long, Y.L. Zhao, et al., J. Agric. Food Chem. 67 (2019) 6962–6969. doi: 10.1021/acs.jafc.9b02059
B. Chen, Q.S. Long, J. Meng, et al., J. Agric. Food Chem. 68 (2020) 2340–2346. doi: 10.1021/acs.jafc.9b07192
P. Li, J. Yin, W.M. Xu, et al., Chem. Bio. Drug. Des. 82 (2013) 546–556. doi: 10.1111/cbdd.12181
M.N. Gao, L. Yu, P. Li, et al., Pestic. Biochem. Phys. 138 (2017) 37–42. doi: 10.1016/j.pestbp.2017.02.004
P. Li, L. Shi, X. Yang, et al., Bioorg. Med. Chem. Lett. 24 (2014) 1677–1680. doi: 10.1016/j.bmcl.2014.02.060
X.P. Song, P. Li, M.W. Li, et al., Pestic. Biochem. Phys. 147 (2018) 11–19. doi: 10.1016/j.pestbp.2017.10.011
A.W. Zhang, H.F. He, R.H. Wang, et al., J. Agric. Food Chem. 72 (2024) 1444–1453. doi: 10.1021/acs.jafc.3c05816
Z.B. Wu, J. Shi, J.X. Chen, D.Y. Hu, B.A. Song, J. Agric. Food Chem. 69 (2021) 8660–8670. doi: 10.1021/acs.jafc.1c01626
J.Z. Li, Q. Liu, S. Li, et al., J. Agric. Food Chem. 72 (2024) 10195–10205. doi: 10.1021/acs.jafc.3c08392
S.B. Wang, J.X. Chen, J. Shi, et al., J. Agric. Food Chem. 69 (2021) 11804–11815. doi: 10.1021/acs.jafc.1c03087
J.X. Chen, C.F. Yi, S.B. Wang, et al., Bioorg. Med. Chem. Lett. 29 (2019) 1203–1210. doi: 10.1016/j.bmcl.2019.03.017
Q.Q. Tao, L.W. Liu, P.Y. Wang, et al., J. Agric. Food Chem. 67 (2019) 7626–7639. doi: 10.1021/acs.jafc.9b02734
J. Xiang, D.Y. Liu, J.X. Chen, D.Y. Hu, B.A. Song, Pestic. Biochem. Phys. 170 (2020) 104695. doi: 10.1016/j.pestbp.2020.104695
S.K. Wu, J. Shi, J.X. Chen, et al., J. Agric. Food Chem. 69 (2021) 4645–4654. doi: 10.1021/acs.jafc.1c01204
Y.Q. Zhang, X. Wang, H.B. Shi, et al., J. Agric. Food Chem. 72 (2024) 9611–9620. doi: 10.1021/acs.jafc.3c07681
F. Xie, T.J.H. Ni, J. Zhao, et al., Bioorg. Med. Chem. Lett. 27 (2017) 2171–2173. doi: 10.1016/j.bmcl.2017.03.062
L.S. Li, X.F. Sun, X.J. Zhao, et al., J. Agric. Food Chem. 69 (2021) 4960–4967. doi: 10.1021/acs.jafc.0c07947
M.H. Ding, S.R. Wan, N. Wu, et al., J. Agric. Food Chem. 69 (2021) 15084–15096. doi: 10.1021/acs.jafc.1c02144
W.B. Shao, P.Y. Wang, Z.M. Fang, et al., J. Agric. Food Chem. 69 (2021) 15108–15122. doi: 10.1021/acs.jafc.1c05202
Q. Fei, C.Y. Liu, Y.B. Luo, et al., Mol. Divers. 29 (2025) 255–267. doi: 10.1007/s11030-024-10848-2
J. Shi, M.H. Ding, N. Luo, et al., J. Agric. Food Chem. 68 (2020) 9613–9623. doi: 10.1021/acs.jafc.0c01365
E. Stingaci, M. Zveaghinteva, S. Pogrebnoi, et al., Bioorg. Med. Chem. Lett. 30 (2020) 127368. doi: 10.1016/j.bmcl.2020.127368
Z.J. Fan, J. Shi, N. Luo, M.H. Ding, X.P. Bao, J. Agric. Food Chem. 67 (2019) 11598–11606. doi: 10.1021/acs.jafc.9b04733
X. Huang, H.W. Liu, Z.Q. Long, et al., J. Agric. Food Chem. 69 (2021) 4615–4627. doi: 10.1021/acs.jafc.1c00707
Y.L. Zhao, X. Huang, L.W. Liu, et al., J. Agric. Food Chem. 67 (2019) 7512–7525. doi: 10.1021/acs.jafc.9b02036
J.R. Rao, L.W. Liu, D. Zeng, et al., RSC Adv. 10 (2020) 682–688. doi: 10.1039/c9ra09639f
H.W. Liu, Q.T. Ji, G.G. Ren, et al., J. Agric. Food Chem. 68 (2020) 12558–12568. doi: 10.1021/acs.jafc.0c02528
M. Xiang, X. Zhou, T.R. Luo, et al., J. Agric. Food Chem. 67 (2019) 13212–13220. doi: 10.1021/acs.jafc.9b06147
Y.M. Feng, P.Y. Qi, W.L. Xiao, et al., J. Agric. Food Chem. 70 (2022) 6037–6049. doi: 10.1021/acs.jafc.2c01141
P.L. Chu, Y.M. Feng, Z.Q. Long, et al., J. Agric. Food Chem. 71 (2023) 6525–6540. doi: 10.1021/acs.jafc.2c07810
W.B. Shao, R.S. Luo, J. Meng, et al., J. Agric. Food Chem. 71 (2023) 16950–16961.
N. Wang, W.J. Jian, H. Liang, et al., Eur. J. Med. Chem. 275 (2024) 116609. doi: 10.1016/j.ejmech.2024.116609
S.S. Liu, D. Zeng, T.H. Zhang, et al., Eur. J. Med. Chem. 250 (2023) 115215. doi: 10.1016/j.ejmech.2023.115215
H.C. Zhao, X.Y. He, J.H. Yang, et al., Adv. Funct. Mater. (2024) 2418415.
L.Z. Xiang, X.H. Wang, J.H. Yang, et al., Chem. Eng. J. 499 (2024) 156140. doi: 10.1016/j.cej.2024.156140
L. Zhou, Y.Z. Chen, J.H. Yang, et al., Chem. Eng. J. 500 (2024) 157282. doi: 10.1016/j.cej.2024.157282
E. Oblak, A. Piecuch, J. Rewak-Soroczynska, E. Paluch, Appl. Microbiol. Biotechnol. 103 (2019) 625–632. doi: 10.1007/s00253-018-9523-2
A.A. Kalinlin, A.D. Voloshina, N.V. Kulik, et al., Eur. J. Med. Chem. 66 (2013) 345–354. doi: 10.1016/j.ejmech.2013.05.038
H.H. Zhu, D. Zeng, M.W. Wang, et al., J. Saudi Chem. Soc. 24 (2020) 127–138. doi: 10.1016/j.jscs.2019.10.002
X. Zhou, Y.M. Feng, P.Y. Qi, et al., J. Agric. Food Chem. 68 (2020) 8132–8142. doi: 10.1021/acs.jafc.0c01565
N. Li, D. Liu, J.K. Dai, et al., Molecules 24 (2019) 1553. doi: 10.3390/molecules24081553
M.W. Wang, H.H. Zhu, P.Y. Wang, J. Agric. Food Chem. 67 (2019) 12696–12708. doi: 10.1021/acs.jafc.9b03952
P.Y. Wang, M.W. Wang, D. Zeng, et al., J. Agric. Food Chem. 67 (2019) 3535–3545. doi: 10.1021/acs.jafc.8b06242
Q.T. Ji, D.K. Hu, X.F. Mu, et al., Nano Lett. 12 (2022) 4839–4847. doi: 10.1021/acs.nanolett.2c01203
X. Wang, T. Liu, R.H. Wang, et al., Pestic. Biochem. Phys. 202 (2024) 105967. doi: 10.1016/j.pestbp.2024.105967
J.X. Chen, Y. Wang, X. Luo, Y.F. Chen, Pestic. Biochem. Phys. 184 (2022) 105122. doi: 10.1016/j.pestbp.2022.105122
Z.Y. Jiang, W.D. Hong, X.P. Cui, et al., RSC Adv. 7 (2017) 52227–52237. doi: 10.1039/C7RA10352B
K.S. Van-Horn, W.N. Burda, R. Fleeman, L.N. Shaw, R. Manetsch, J. Med. Chem. 57 (2014) 3075–3093. doi: 10.1021/jm500039e
J. Meng, L. Zhang, X.X. Tuo, et al., J. Adv. Res. 74 (2025) 57–72. doi: 10.1016/j.jare.2024.10.002
J. Shi, N. Luo, M.H. Ding, X.P. Bao, Chin. Chem. Lett. 31 (2020) 434–438. doi: 10.1016/j.cclet.2019.06.037
Q.S. Long, L.W. Liu, Y.L. Zhao, et al., J. Agric. Food Chem. 67 (2019) 11005–11017. doi: 10.1021/acs.jafc.9b03419
W.B. Shao, Y.T. Zheng, J.M. Liu, et al., Bioorg. Med. Chem. Lett. 30 (2020) 126912. doi: 10.1016/j.bmcl.2019.126912
L.H. Shao, S. Zhao, S. Yang, et al., J. Agric. Food Chem. 71 (2023) 3939–3949. doi: 10.1021/acs.jafc.2c07264
G.L. Zhang, C.P. Li, Y. Li, et al., J. Agric. Food Chem. 71 (2023) 14232–14242. doi: 10.1021/acs.jafc.3c03738
N. Wu, Y.H. Yang, G.M. Tian, et al., J. Agric. Food Chem. 71 (2023) 19277–19287. doi: 10.1021/acs.jafc.3c03670
G.L. Zhang, C.P. Li, Y. Li, et al., J. Agric. Food Chem. 71 (2023) 6288–6300. doi: 10.1021/acs.jafc.3c01120
L. An, L. Yang, T.S. Yan, et al., Pest Manag. Sci. 80 (2024) 5307–5321. doi: 10.1002/ps.8256
Z.Y. Tan, J. Deng, Q.X. Ye, Z.F. Zhang, Curr. Top. Med. Chem. 22 (2022) 1009–1019. doi: 10.2174/1568026622666220221110506
J.R. Liang, X. Huang, G. Ma, RSC Adv. 12 (2022) 29197–29213. doi: 10.1039/d2ra02389j
T.T. Liu, F. Peng, X. Cao, et al., ACS Omega 6 (2021) 30826–30833. doi: 10.1021/acsomega.1c05256
S.J. Su, Q. Zhou, X.M. Tang, et al., Monatsh. Chem. 152 (2021) 345–356. doi: 10.1007/s00706-021-02739-1
S.C. Jiang, X. Tang, M. Chen, et al., Pest Manag. Sci. 76 (2020) 853–860. doi: 10.1002/ps.5587
S.C. Jiang, S.J. Su, M. Chen, et al., J. Agric. Food Chem. 68 (2020) 5641–5647. doi: 10.1021/acs.jafc.0c01652
M. Chen, X.M. Tang, T.T. Liu, et al., Fitoterapia 149 (2021) 104804. doi: 10.1016/j.fitote.2020.104804
M.H. Zhang, S. Feng, S. Chen, et al., Pest Manag. Sci. 79 (2023) 4795–4808. doi: 10.1002/ps.7675
Q. Bian, R.Q. Zhao, X.J. Peng, et al., J. Agric. Food Chem. 69 (2021) 3848–3858. doi: 10.1021/acs.jafc.0c07581
Z.B. Wu, H.Y. Park, D.W. Xie, et al., J. Agric. Food Chem. 69 (2021) 1214–1223. doi: 10.1021/acs.jafc.0c05702
S.K. Sahu, P. Zheng, N. Yao, Front. Plant Sci. 9 (2018) 408. doi: 10.3389/fpls.2018.00408
X.W. Xiang, H. Tao, S. Jiang, L.H. Zhang, Z.N. Cui, Pestic. Biochem. Phys. 149 (2018) 89–97. doi: 10.1016/j.pestbp.2018.06.011
K.M. Friedrich, K. Uirich, M. Randy, et al., Patent, WO 2018/109062, 2018.
R.F. Zhang, S.X. Guo, P. Deng, et al., J. Agric. Food Chem. 69 (2021) 10082–10092. doi: 10.1021/acs.jafc.1c03892
Y.T. Wang, T.Q. Shi, J. Fu, H.L. Zhu, Eur. J. Med. Chem. 171 (2019) 209–220. doi: 10.1016/j.ejmech.2019.03.026
W. Liu, S. Zhang, L.F. Xiao, et al., Pest Manag. Sci. 78 (2022) 1438–1447. doi: 10.1002/ps.6761
Y.L. Chen, T.T. Li, Z.C. Jin, Y.G.R. Chi, et al., J. Agric. Food Chem. 70 (2022) 6050–6058. doi: 10.1021/acs.jafc.2c01407
S.X. Guo, F. He, W. Zhang, et al., Food Energy Secur 12 (2023) e449. doi: 10.1002/fes3.449
S.X. Guo, F. He, W. Zhang, et al., Food Energy Secur. 12 (2023) e449. doi: 10.1002/fes3.449
H.W. Liu, S.S. Su, S.Y. Ma, et al., J. Agric. Food Chem. 71 (2023) 11035–11047. doi: 10.1021/acs.jafc.3c02615
J.B. Li, L.T. Xiong, Y.R. Lu, et al., Pest Manag. Sci. 79 (2023) 3666–3675. doi: 10.1002/ps.7545
A.W. Zhang, H.Z. Zhang, R.H. Wang, et al., Sci. China Life Sci. 67 (2024) 391–402. doi: 10.1007/s11427-023-2449-1
A.W. Zhang, R.J. Song, R.H. Wang, et al., J. Agric. Food Chem. 70 (2022) 9356–9366. doi: 10.1021/acs.jafc.2c02605
X.G. Wang, S.Y. Li, D.Y. Liu, et al., Pestic. Biochem. Phys. 202 (2024) 105955. doi: 10.1016/j.pestbp.2024.105955
X.C. Chen, X. Niu, L.J. Li, et al., J. Agric. Food Chem. 72 (2024) 3436–3444. doi: 10.1021/acs.jafc.3c09267
Y.H.I. Mohammed, I.M. Shamkh, A.H. Shntail, et al., Sci. Rep. 14 (2024) 11118. doi: 10.1038/s41598-024-59609-z
S. Jiang, H. Li, W. Ahmed, et al., Front. Microbiol. 10 (2019) 1874. doi: 10.3389/fmicb.2019.01874
H. Tao, H. Tian, S. Jiang, et al., Pestic. Biochem. Phys. 160 (2019) 87–94. doi: 10.1016/j.pestbp.2019.07.005
S. Jiang, M. He, X.W. Xiang, M. Adnan, Z.N. Cui, J. Agric. Food Chem. 67 (2019) 11867–11876. doi: 10.1021/acs.jafc.9b04085
H.L. Mou, J. Shi, J.X. Chen, D.Y. Hu, Pestic. Biochem. Phys. 178 (2021) 104913. doi: 10.1016/j.pestbp.2021.104913
H. Tao, H. Tian, S. Jiang, et al., Bioorg. Med. Chem. 27 (2019) 3364–3371. doi: 10.1016/j.bmc.2019.06.019
Y.G. Liu, G.Y. Luo, X. Yang, et al., Angew. Chem. Int. Ed. 59 (2020) 442. doi: 10.1002/anie.201912160
Y.L. Song, S.S. Liu, J. Yang, et al., Int. J. Mol. Sci. 23 (2022) 9119. doi: 10.3390/ijms23169119
K.P. Teng, Q. Liu, M. Zhang, et al., J. Agric. Food Chem. 72 (2024) 4622–4629. doi: 10.1021/acs.jafc.3c07491
Y. Zhang, W.Q. Zhang, L.L. Han, et al., Mol. Plant Pathol. 20 (2019) 1477–1490. doi: 10.1111/mpp.12854
S. Mishra, X.D. Yang, S. Ray, L.F. Fraceto, H.B. Singh, World J. Microb. Biot. 36 (2020) 55. doi: 10.9734/arrb/2020/v35i930271
D.X. Xiao, J.L. Cheng, W.L. Liang, et al., J. Agric. Food Chem. 67 (2019) 11577–11583. doi: 10.1021/acs.jafc.9b03700
M.A. Tsiaggali, E.G. Andreadou, A.G. Hatzidimitriou, A.A. Pantazaki, P. Aslanidis, J. Inorg. Biochem. 121 (2013) 121–128. doi: 10.1016/j.jinorgbio.2013.01.001
E. Kouris, S. Kalogiannis, F. Perdih, I. Turel, G. Psomas, J. Inorg. Biochem. 163 (2016) 18–27. doi: 10.1016/j.jinorgbio.2016.07.022
S. Naz, M. Sirajuddin, I. Hussain, et al., J. Mol. Struct. 1203 (2020) 127378. doi: 10.1016/j.molstruc.2019.127378
L.B. Jiang, Y.R. Liu, X.Y. Xu, et al., Front. Microbiol. 11 (2020) 571691. doi: 10.3389/fmicb.2020.571691
S. Chandra, S. Bargujar, R. Nirwal, K. Qanungo, S.K. Sharma, Spectrochim. Acta. A 113 (2013) 164–170. doi: 10.1016/j.saa.2013.04.114
H.G. Cao, Y.F. Yang, X.J. Chen, et al., Chin. Chem. Lett. 31 (2020) 1887–1889. doi: 10.1016/j.cclet.2020.01.027
D. Anastasiadoua, G. Psomasa, S. Kalogiannisb, et al., J. Inorg. Biochem. 198 (2019) 110750. doi: 10.1016/j.jinorgbio.2019.110750
S. Ntanatsidis, S. Perontsis, S. Konstantopoulou, et al., J. Inorg. Biochem. 227 (2022) 111693. doi: 10.1016/j.jinorgbio.2021.111693
X.Y. Chen, S.H. Zhuang, W.Y. Yan, et al., Chin. Chem. Lett. 35 (2024) 109635. doi: 10.1016/j.cclet.2024.109635
C.F. Yi, J.X. Chen, C.Q. Wei, et al., Bioorg. Med. Chem. Lett. 30 (2020) 126814. doi: 10.1016/j.bmcl.2019.126814
D.Y. Liu, J. Zhang, L. Zhao, et al., J. Agric. Food Chem. 67 (2019) 11860–11866. doi: 10.1021/acs.jafc.9b03606
T. Liu, J. Shi, D.Y. Liu, et al., J. Agric. Food Chem. 70 (2022) 99–110. doi: 10.1021/acs.jafc.1c04715
X.Q. Yu, X.Y. Zhu, Y. Zhou, et al., J. Agric. Food Chem. 67 (2019) 13904–13913. doi: 10.1021/acs.jafc.9b06296
X.M. Song, X.Y. Zhu, T. Li, et al., J. Agric. Food Chem. 67 (2019) 11354–11363. doi: 10.1021/acs.jafc.9b04231
M. Zhang, M.L. Dou, Y.Y. Xia, et al., J. Agric. Food Chem. 69 (2021) 5435–5445. doi: 10.1021/acs.jafc.1c00272
L.B. Li, W.J. Dan, F.F. Tan, et al., Chem. Pharm. Bull. 63 (2015) 33–37. doi: 10.1248/cpb.c14-00578
R. Wu, T. Liu, S.K. Wu, et al., J. Agric. Food Chem. 70 (2022) 9305–9318. doi: 10.1021/acs.jafc.2c02099
W.L. Xiao, N. Wang, L.L. Yang, et al., J. Agric. Food Chem. 71 (2023) 6603–6616. doi: 10.1021/acs.jafc.3c00465
S.S. Su, H.W. Liu, J.R. Zhang, et al., J. Integr. Agric. 23 (2024) 1259–1273. doi: 10.1016/j.jia.2023.05.031
Y. Ma, H. Zhou, Y.R. Wang, et al., Pest Manag. Sci. 79 (2023) 5321–5332. doi: 10.1002/ps.7742
Y. Shi, Z.S. Zhang, J. Shao, et al., Chin. Chem. Lett. 35 (2024) 1087.
J. Shao, Z.J. Zhang, Y. Shi, et al., J. Agric. Food Chem. 72 (2024) 6988–6997. doi: 10.1021/acs.jafc.4c00214
K. Tang, F. Su, S.J. Pan, et al., Chin. Chem. Lett. 35 (2024) 109495. doi: 10.1016/j.cclet.2024.109495
H.J. Dai, J.H. Yang, L.J. Fan, et al., Adv. Funct. Mater. 34 (2024) 2403823. doi: 10.1002/adfm.202403823

Figure 11 Schematic diagram of the mechanism of action of agricultural antibacterial agents.
Table 1. Antibacterial activities of oxadiazole/thiadiazole compounds and mechanism of action.
| Compd | Bacterial | In vitro antibacterial activities/ EC50 (µg/mL) | Protective activities at 200 µg/mL (%) | Mechanism of action | Ref. |
| 1 | Xoo | 9.9 | / | DLST | [35,36] |
| Rs | 8.3 | / | [37,38] | ||
| 2 | Xoo | 1.1 | 43.5 | Metabolism of pyrimidine | [39,40] |
| Xoc | 7.1 | / | |||
| 3 | Xoo | 4.8 | / | Purine metabolism | [41] |
| 4 | Xoo | 5.3 | 44.3 | CsrA and Xoc3530 | [42] |
| Xoc | 7.6 | 47.5 | |||
| 5 | Xoo | 1.8 | 51.3 | Glycolysis/gluconeogenesis | [43] |
| Xoc | 2.1 | 45.9 | |||
| 6 | Xoo | 0.3 | 40.3 | Interferes with proteins in the bacterial glycolysis-gluconeogenesis pathway | [44] |
| Xoc | 0.6 | 42.0 | |||
| Xac | 10.4 | / | |||
| 7 | Xoo | 0.4 | / | Glutathione metabolism | [45] |
| Xoc | 0.2 | 35.4 | |||
| 8 | Xoo | 0.4 | 55.8 | / | [46] |
| Xac | 6.7 | / | |||
| Rs | 5.1 | / | |||
| 9 | Xoo | 13.4 | 56.0 | Histidine metabolism and starch and sucrose metabolism | [47] |
| Rs | 33.5 | / | |||
| Xac | 24.6 | / | |||
| 10 | Xoo | 0.8 | / | / | [48] |
| Xoc | 2.2 | / | |||
| 11 | Xoo | 0.6 | 49.4 | Photosynthetic | [49] |
| Xoc | 1.6 | 49.7 | |||
| 12 | Xac | / | / | Inhibited hpa1 transcription and hypersensitive response (HR) | [50] |
| Xoc: Xanthomonas oryzae pv. oryzicola, Xac: X. axonopodis pv. citri. | |||||
 下载: 导出CSV
下载: 导出CSV
Table 2. Antibacterial activities of triazole compounds and mechanism of action.
| Compd | Bacterial | In vitro antibacterial activities | Protective activities at 200 µg/mL (%) | Mechanism of action | Ref. |
| 13 | Xoo | EC50 = 18.8 µg/mL | 50.8 | Destroyed the cell membrane structure | [53] |
| 14 | Xoo | EC50 = 5.0 µg/mL | 54.2 | Downregulated the expression of related proteins in the type Ⅲ secretion system and transcriptional activator of Xoo | [54] |
| 15 | Xac | EC50 = 38.2 µg/mL | 61.1 | Enhance the permeability of bacterial membrane and change the morphology of bacterial cells | [55] |
| 16 | Xoo | EC50 = 18.8 µg/mL | 56.2 | / | [56] |
| 17 | Erwinia amylovora | MIC = 0.0004 mmol/L | / | / | [57] |
| Erwinia carotovora | MIC = 0.0004 mmol/L | ||||
| X. campestris | MIC = 0.0008 mmol/L | ||||
| 18 | Xac | EC50 = 10.0 µg/mL | / | / | [58] |
| Xoo | EC50 = 7.2 µg/mL | 49.5 | |||
| 19 | Psa | EC50 = 4.6 µg/mL | 79.4 | / | [59] |
| 20 | Xoo | EC50 = 3.4 µg/mL | 53.2 | Interfere with the balance of the pathogen's redox system | [59] |
| Xac | EC50 = 2.9 µg/mL | / | |||
| Psa: Pseudomonas syringae pv. actinidiae. | |||||
下载: 导出CSV
Table 3. Antibacterial activities of isopropanolamine compounds and mechanism of action.
| Compd | Bacterial | In vitro antibacterial activities/EC50 (µg/mL) | Protective activities at 200 µg/mL (%) | Mechanism of action | Ref. |
| 21 | Xoo | 1.3 | 48.7 | / | [60] |
| Xac | 2.0 | / | |||
| Psa | 3.8 | / | |||
| 22 | Xoo | 5.0 | 48.2 | / | [61] |
| Xac | 4.7 | / | |||
| Psa | 8.5 | / | |||
| 23 | Xoo | 1.7 | 51.9 | / | [62] |
| Xac | 4.1 | / | |||
| Psa | 2.4 | 65.5 | |||
| 24 | Xoo | 4.8 | 51.8 | / | [63] |
| Xac | 3.2 | / | |||
| 25 | Xoo | 6.8 | 43.6 | Inhibit DSF biosynthesis and related QS pathway gene expression | [64] |
| Xoc | 8.9 | / | |||
| Xac | 3.1 | / | |||
| 26 | Xoo | 1.5 | 48.7 | RpfF | [65] |
| Xac | 2.0 | / | |||
| Psa | 5.7 | / | |||
| 27 | Xoo | 0.8 | 52.0 | Destroy cell morphology, and interacts with DNA in a groove binding mode | [66] |
| Xac | 4.8 | / | |||
| Psa | 2.3 | 80.0 | |||
| 28 | Xoo | 9.9 | 42.6 | RpfF | [67] |
| Xac | 7.0 | ||||
| 29 | Xoo | 1.3 | 43.3 | Interferes with genes involved in fatty acid synthesis and damages the integrity of bacterial cell membranes | [68] |
| Xac | 1.4 | / | |||
| Psa | 8.5 | / | |||
| Cmm | 8.2 | / | |||
| 30 | Xoo | 1.8 | 48.9 | Inhibit the formation of biofilms, alter cell membrane permeability, reduce the production of EPS, and decrease bacterial motility | [69] |
| 31 | Xac | 1.9 | 81.4 | Destruction of cell membrane | [70] |
| 32 | Xoo | 2.0 | 50.8 | Destroy cell membrane and inhibit EPS production | [71] |
| Cmm: Clavibacter michiganense subsp. michiganse. | |||||
下载: 导出CSV
Table 4. Antibacterial activities of onium salts compounds and mechanism of action.
| Compd | Bacterial | In vitro antibacterial activities (µg/mL) | Protective activities at 200 µg/mL (%) | Mechanism of action | Ref. |
| 33 | Xoo | EC50 = 1.0 | 63.2 | FtsZ | [75] |
| 34 | Rs | MIC = 3.9 | / | / | [76] |
| Ps | MIC = 3.9 | / | |||
| 35 | Xoo | EC50 = 0.1 | 51.5 | Disrupt the ribosome assembly pathway | [77] |
| Rs | EC50 = 9.6 | / | |||
| Xac | EC50 = 14.6 | / | |||
| 36 | Xoo | EC50 = 0.3 | / | Destroy cell morphology | [78] |
| Rs | EC50 = 0.6 | / | |||
| 37 | Xoo | EC50 = 0.6 | 42.6 | / | [79] |
| 38 | Xoo | EC50 = 0.2 | 55.3 | The activities of POD and phenylalanine ammonia-lyase (PAL) defense enzymes and proteins related to pyruvate metabolic pathway were increased | [80] |
| Xoc | EC50 = 1.1 | / | |||
| Xac | EC50 = 0.8 | / |
下载: 导出CSV
Table 5. Antibacterial activities of quinazoline compounds and mechanism of action.
| Compd | Bacterial | In vitro antibacterial activities/EC50 (µg/mL) | Protective activities at 200 µg/mL (% | Mechanism of action | Ref. |
| 39 | Xoo | 13.2 µmol/L | 49.6 | FabF | [84] |
| 40 | Xoo | 26.0 | / | / | [85] |
| 41 | Xoo | 7.1 | / | Affect the metabolic pathways of D-glucose and biotin | [86] |
| Xac | 18.6 | / | |||
| 42 | Rs | 2.7 | / | Destructive bacterial morphology | [87] |
| 43 | Xoo | 1.5 | 46.7 | Affects ribosomal protein synthesis and transport, energy metabolism pathway and glutathione metabolism disorders | [88] |
| 44 | Xac | 0.8 | / | Interferes with the T2SS and T4SS secretory systems of Xac | [89] |
| Xoo | 1.3 | / | |||
| Psa | 15.5 | / | |||
| 45 | Xoc | 19.5 | / | Inhibit the formation of EPS, change the permeability of membrane and destroy the morphology of bacteria | [90] |
| Xoo | 4.1 | 47.6 | |||
| 46 | Xac | 0.3 | / | Interferes with the T4SS secretory systems of Xac | [91] |
| Xoo | 1.9 | / | |||
| Psa | 18.0 | / | |||
| 47 | Xoc | 7.1 | 50.5 | Down-regulating proteins associated with bacterial flagellar biosynthesis | [92] |
下载: 导出CSV
Table 6. Antibacterial activities of flavonoid compounds and mechanism of action.
| Compd | Bacterial | In vitro antibacterial activities/ EC50 (µg/mL) | Protective activities at 200 µg/mL (%) | Mechanism of | Ref. |
| 48 | Psa | 28.1 | / | Destroy cell morphology | [95] |
| Xac | 16.9 | / | |||
| Xoo | 20.4 | / | |||
| 49 | Xoo | 11.9 | 47.0 | [96] | |
| Rs | 17.1 | / | |||
| 50 | Xoo | 4.7 | 63.3 | [97] | |
| 51 | Xoo | 1.6 | 60.1 | [98] | |
| 52 | Xoo | 8.2 | 48.6 | [99] | |
| 53 | Xoo | 2.7 | 64.2 | [100] | |
| Xac | 7.1 | / |
下载: 导出CSV
Table 7. Antibacterial activities of amide compounds and mechanism of action.
| Compd | Bacterial | In vitro antibacterial activities | In vivo antibacterial activities | Mechanism of action | Ref. |
| 54 | Xoo | 98.05% inhibitors of Xoo T3SS at 10 µg/mL | / | Inhibit T3SS | [104] |
| 55 | Ps | / | 39.2% at 75 g a.i./ha | / | [105] |
| 56 | Xoo | EC50 = 11.6 µg/mL | 47.0% at 200 µg/mL | / | [106] |
| 57 | Xoo | MIC = 0.5 µg/mL | / | FabH | [107] |
| 58 | Erwinia carotorora | EC50 = 2.7 µg/mL | / | / | [108] |
| 59 | Xoo | EC50 = 4.2 µg/mL | 53.7% at 200 µg/mL | Affects glyoxylate and dicarboxylate metabolism and galactose metabolism in rice | [109] |
| 60 | Xoo | EC50 = 28.6 µg/mL | 51.3% at 200 µg/mL | Destroy cell morphology | [110] |
| Rs | EC50 = 55.3 µg/mL | / | |||
| 61 | Cmm | EC50 = 4.1 µg/mL | 71.8% at 100 µg/mL | Causes peroxidation damage and leakage of electrolytes and proteins from the bacterial cells | [111] |
| 62 | Xoo | EC50 = 5.7 µg/mL | 55.7% at 200 µg/mL | Disrupt the balance of the redox system | [112] |
| Xac | EC50 = 6.6 µg/mL | / | |||
| Psa | EC50 = 9.1 µg/mL | ||||
| 63 | Xcc | 77.5% inhibitors of Xcc T3SS at 20 µg/mL | / | Inhibit T3SS | [113] |
| 64 | Xoo | EC50 = 0.4 µg/mL | 50.0% at 200 µg/mL | Pyruvate kinase | [114] |
| Xoc | EC50 = 3.1 µg/mL | / | |||
| Xac | EC50 = 2.9 µg/mL | ||||
| 65 | Xoo | EC50 = 0.6 µg/mL | 45.3% at 200 µg/mL | Clp | [115] |
| Xoc | EC50 = 0.8 µg/mL | 51.7% at 200 µg/mL | |||
| 66 | Xoo | EC50 = 1.1 µg/mL | / | Up-regulated expression of genes OsBIPP2C1, OsPYL/RCAR5 and OsABF1 activated ABA signaling pathway | [116] |
| Xoc | EC50 = 0.9 µg/mL | 70.3% at 200 µg/mL | |||
| Xac | EC50 = 1.8 µg/mL | 64.4% at 200 µg/mL | |||
| 67 | Xoo | EC50 = 5.1 µg/mL | / | 50S ribosomal protein L2 | [117] |
| Xac | EC50 = 5.2 µg/mL | ||||
| 68 | Rs | / | / | Fucose-binding protein (PDB ID: 4CSD) | [118] |
下载: 导出CSV
Table 8. Antibacterial activities of esters compounds and mechanism of action.
| Compd | Bacterial | Concentration | Antibacterial activities | Mechanism of action | Ref. |
| 69 | Xoo | 100 mmol/L | 99.0% inhibitors of Xoo T3SS | Inhibit T3SS | [119] |
| 70 | Xoo | 200 µg/mL | 58.1% | [120] | |
| 71 | Xoo | 200 µg/mL | 63.8% | [121] | |
| 72 | Xoo | / | EC50 = 35.8 µg/mL | Regulate the oxidative phosphorylation pathway of rice | [122] |
| 73 | Xoo | 10 µg/mL | 65.2% inhibitors of Xoo T3SS | Inhibit T3SS | [123] |
| 74 | Rs | 50 µg/mL | 88.2% | [124] | |
| 75 | Xoo | 100 µmol/L | 48.6% (the FtsZ GTPase activity) | FtsZ | [125] |
| 76 | Xoo | / | EC50 = 5.8 µg/mL | DHPS (dihydropteroate synthase) | [126] |
下载: 导出CSV
Table 9. Antibacterial activity of other compounds and mechanism of action.
| Compounds | Bacterial | In vitro antibacterial activities (µg/mL) | Protective activities at 200 µg/mL (%) | Mechanism of action | Ref. |
| 77 | Xoo | EC50 = 0.1 | 52.1 at 50 µg/mL | Inhibit the formation of extracellular polysaccharide and biofilm of Xoo, and change the permeability of cell membrane | [139] |
| Xac | EC50 = 0.2 | ||||
| 78 | Xoo | EC50 = 1.1 | 75.1 | / | [140] |
| Xac | EC50 = 12.4 | ||||
| 79 | Xoo | EC50 = 36.8 | 49.2 | Enhance SOD, PAL, CAT and oxidative phosphorylation pathways in rice | [141] |
| Xoc | EC50 = 47.6 | 39.7 | |||
| 80 | Xoo | EC50 = 26.4 | / | / | [142] |
| 81 | Xoo | EC50 = 11.4 | 51.7 | / | [143] |
| 82 | Xoo | EC50 = 8.3 | 57.4 | / | [144] |
| Xoc | EC50 = 25.9 | / | |||
| 83 | Rs | MIC = 3.9 | / | / | [145] |
| Psa | MIC = 7.8 | ||||
| 84 | Xoo | EC50 = 26.7 | 45.1 | Increased POD and CAT in rice, upregulated ATP synthase subunit d and NADH dehydrogenase iron-sulfur 6 | [146] |
| Xoc | EC50 = 30.8 | 43.7 | |||
| Xac | EC50 = 65.4 | ||||
| 85 | Xoo | EC50 = 10.0 | 49.2 | Inhibition of extracellular enzymes, EPS, biofilm, T3SS effector virulence factors | [147] |
| Xac | EC50 = 13.3 | ||||
| 86 | Xoo | EC50 = 7.3 | 52.7 | Destroy the redox system of Xoo, causing ROS accumulation | [148] |
| Xac | EC50 = 4.9 | ||||
| 87 | Xoo | MIC = 3.1 | 55.9 | Destroy cell morphology and inhibit the production of EPS | [149] |
| Xac | MIC = 3.1 | ||||
| 88 | Xoo | EC50 = 6.6 | 37.5 | Destroy cell morphology | [150] |
| 89 | Xoo | 83.4% inhibitors of hpa1 at 10 µg/mL | / | Inhibit the transcription of hpa1, hrpG, hrpX and hrcT in the T3SS | [151] |
| 90 | Xoo | 84.8% at 100 µg/mL | / | / | [152] |
| 91 | Xoo | EC50 = 9.4 µg/mL | 48.1 | Inhibit the transcription of hrpG, hrpX, hrpE and hrpF in the T3SS | [153] |
下载: 导出CSV

 扫一扫看文章
扫一扫看文章

扫一扫关注我们










 下载:
下载: