Received Date:
16 February 2025 Accepted Date:
14 May 2025 Revised Date:
12 May 2025 Available Online:
15 May 2026
Abstract:
Tumor-associated carbohydrate antigen (TACA)-based cancer vaccines face clinical challenges due to heterogeneous TACA expression, which compromises antibody-mediated tumor recognition and leads to suboptimal therapeutic outcomes. To address this limitation, we report a combined strategy that integrates vaccination with TACA-based antibody-recruiting molecules. This approach simultaneously redirects anti-TACA antibodies to tumor cells expressing a secondary target, thereby enhancing the efficacy of TACA-based vaccines. Using sialyl-Tn (sTn) as a model TACA and epidermal growth factor receptor (EGFR) and human epidermal growth factor receptor 2 (HER2) as model protein targets, we designed two nanobody (Nb)-sTn conjugates as TACA-based antibody-recruiting molecules: EGFR-targeting 7D12-sTn and HER2-targeting C7b-sTn. These conjugates were synthesized via sortase A-mediated ligation and demonstrated strong binding profiles. Importantly, they effectively redirected anti-sTn antibodies, generated by the Theratope vaccine, to target cells in situ, significantly improving the recognition of tumor cells by anti-sTn antibodies. The synergistic potential of these conjugates in amplifying the therapeutic effect of the sTn-KLH vaccine was further validated through complement-dependent cytotoxicity assays. This innovative strategy represents a highly promising approach to overcome the clinical challenges posed by TACA heterogeneity in cancer vaccine development.
Tumor-associated carbohydrate antigen (TACA) is a sort of aberrant glycan that overexpressed on the surface of many types of human cancer cells [1]. These carbohydrates play a crucial role in facilitating tumor formation, progression, and metastasis [2]. Due to their unique structure and distribution on cancer cells, TACAs have emerged as promising immune targets for vaccine development [3]. However, two primary challenges hinder the development of TACA-based vaccines: poor immunogenicity and immunotolerance [4,5]. Recent advancements in vaccine design have introduced several strategies to address these issues. These include glycoconjugate vaccines [6,7], polyvalent TACA vaccines [8,9], self-adjuvant vaccines [10-12], adjuvant formulations [13,14], non-natural TACA-based vaccines [15,16], glycoengineering-based vaccine [17,18], and among others [19,20]. These TACA-based vaccines have been shown to elicit successful T-cell dependent immune responses and generate anti-TACA antibodies capable of destroying cancer cells. Despite this promise, most of these strategies remain confined to preclinical research, with few achieving significant clinical efficacy [21].
Sialyl-Tn (sTn, Neu5Acα2–6GalNAcα1-O-Ser/Thr) is a typical TACA associated with MUC1, which is frequently overexpressed in various carcinomas, including those of the pancreas, lung, breast, prostate, bladder, and ovary [22]. The overexpression of sTn antigen can alter tumor malignant properties, leading to more aggressive cancer cell behavior, including decreased cell-cell aggregation, increased tumor growth, and metastasis. Moreover, sTn impairs the maturation and function of dendritic cells, limiting their ability to elicit protective anti-tumor T cell responses. Importantly, the absence of sTn in healthy tissue makes it to be an ideal candidate for cancer vaccine development [23,24]. Theratope, a glycoconjugate vaccine that combines the sTn disaccharide with the carrier protein keyhole limpet hemocyanin (KLH), is the most extensively studied TACA vaccine in clinical trials. Theratope has shown a favorable safety profile and elicited T-cell-dependent immune responses [25]. Notably, high titers of anti-sTn IgG/IgM antibodies can trigger antibody-dependent cellular cytotoxicity, effectively killing cancer cells and delaying tumor growth in mice, while also increasing progression-free survival in patients. However, a Phase Ⅲ trial involving over 1000 patients with metastatic breast cancer found that Theratope failed to meet its endpoints for delaying disease progression and improving overall survival [26].
The reasons for the disappointing clinical outcomes of the Theratope vaccine are multifaceted. One significant factor is the heterogeneity of sTn expression within tumors, which correlates with various clinical characteristics and outcomes [22,27-29]. In the enrolled patients, sTn expression level varied widely, ranging from 16% to 91% [23]. This substantial variability in sTn levels complicates the ability of vaccine-induced antibodies to accurately recognize and bind to tumor cells, which is believed to be a key reason for the lack of success in therapeutic cancer vaccines targeting sTn. A similar issue arose with adagloxad simolenin (AdaSim), a synthetic Globo H conjugate vaccine, where the heterogeneity of Globo H expression influenced the vaccine's anticancer efficacy in clinical investigation [30]. Although existing TACA-based vaccines have demonstrated good safety and can produce high-titer of T-dell dependent anti-TACA antibodies, the significant heterogeneity in TACA expression on tumor cells prevents antibodies from efficiently recognizing and effectively targeting tumor cells, leading to the failure of these vaccines. Therefore, there is an urgent need to develop new strategies to overcome the challenge posed by TACA expression heterogeneity, with the goal of enhancing the therapeutic efficacy of TACA-based vaccines.
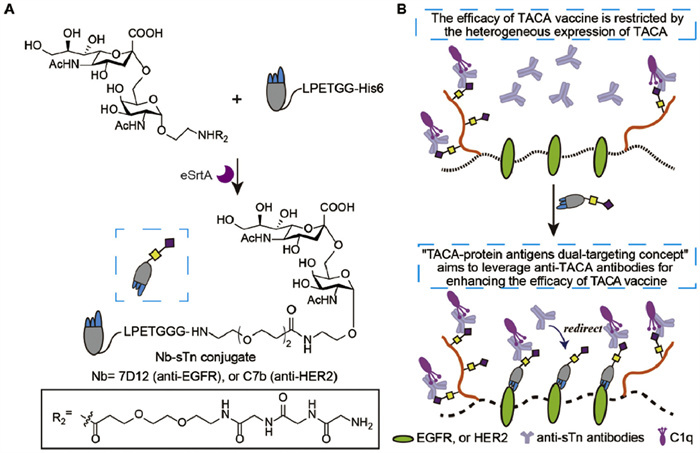
Redirecting naturally occurring endogenous antibodies, such as anti-DNP, anti-Rhamnose, and anti-α-Gal, to target cell surfaces—facilitated by rationally designed antibody-recruiting molecules—can activate the host immune system and trigger downstream immune responses to effectively eliminate target cells [31-35]. This strategy has emerged as a novel immunotherapeutic alternative to the administration of exogenous therapeutic monoclonal antibodies. For instance, the zanamivir-DNP-rhamnose conjugate can recruit the immune system and rapidly eradicate influenza virus infections more swiftly than any current therapy [36]. This therapeutic strategy has progressed into clinical development. Building on this concept and the sTn-based vaccine research, we propose a combined strategy that integrates vaccination with TACA-based antibody-recruiting molecules. This approach simultaneously redirects anti-TACA antibodies to tumor cells expressing a secondary target, thereby enhancing the efficacy of TACA-based vaccines (Fig. 1). Specifically, the sTn antigen was site-specifically conjugated with a tumor-targeting nanobody (Nb) using sortase-mediated ligation (SML, Fig. 1A). This sTn-Nb conjugate can target cell surface protein antigens and in situ recruit additional anti-TACA antibodies generated by the sTn-KLH vaccine. This dual-targeting approach aims to: (1) Increase the recognition of tumor cells by the immune system through the simultaneous targeting of TACA antigens (by anti-TACA antibodies) and cell surface protein antigens (by the Nb-sTn conjugate); (2) Enhance the elimination of heterogeneous tumor cells by recruiting more anti-TACA antibodies to the tumor site via the Nb-sTn conjugate, thereby addressing the challenge posed by the heterogeneity of TACA expression on tumor cells (Fig. 1B).
Figure 1
Figure 1.
Illustration of the "TACA-protein antigens dual-targeting concept" to enhance the effect of TACA-based cancer vaccine. (A) Design and construction of Nb-sTn conjugates as a synergistic agent. (B) When compared to sTn-KLH alone, synergetic treatment with Nb-sTn can achieve better efficacy by redirecting more anti-sTn antibodies onto targets.
By leveraging both TACA antigen recognition and the recruitment of additional anti-TACA antibodies, this proposed strategy has the potential to improve the overall efficacy of TACA-based cancer vaccines.
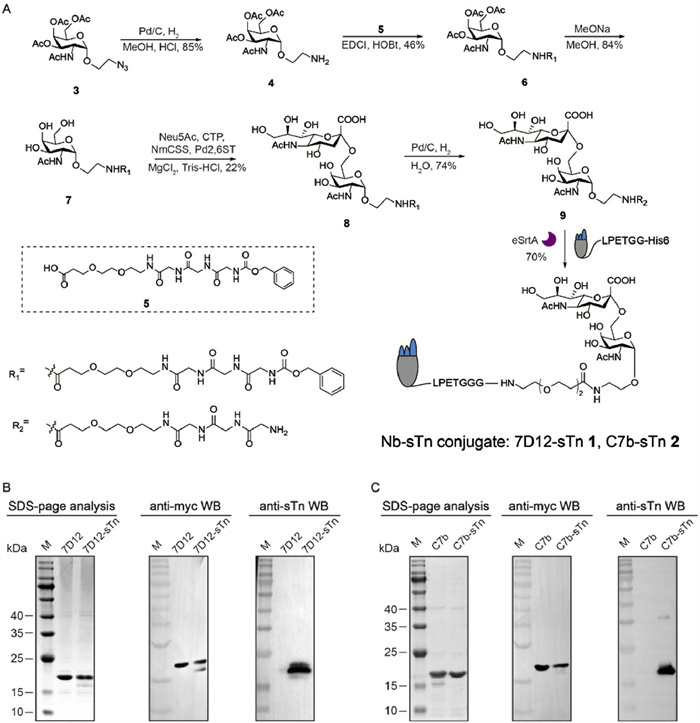
To validate the effectiveness of this dual-targeting concept, we first intended to prepare Nb-sTn conjugates via SML. Two known clinical validated targets, epidermal growth factor receptor (EGFR) and human epidermal growth factor receptor 2 (HER2), were chosen as models. Accordingly, an anti-EGFR Nb 7D12, and an anti-HER2 Nb C7b, were utilized. These Nbs were engineered with a C-terminal myc tag for immunoassay, a sortase A (SrtA) recognition tag for site-specific modification, and a 6× His tag for affinity chromatography. Both 7D12 and C7b were expressed in Escherichia coli (E. coli) with high yields. Meanwhile, to enable site-specific conjugation of the sTn antigen to Nbs, an oligoglycine-modified sTn antigen 9 was required (Fig. 2A). Based on our previous structure-activity relationship studies of nanobody-antigen linker configurations [32], we designed sTn antigen 9 by introducing a PEG3-equivalent distance spacer between the sTn and triglycine motif. Briefly, peracetylated Tn analog 4 was coupled with linker 5 to afford compound 6 in moderate yields. Next, the acetyl groups were removed under Zemplén transesterification conditions to obtain 7. Subsequently, a one-pot enzymatic method was employed, utilizing sialic acid synthetase (NmCSS) and α2,6-sialyltransferase (Pd2,6ST) to transfer sialic acid to the 6-position of acetylgalactosamine to generate 8. After removal of the Cbz protecting group, the triglycine-modified sTn antigen 9 was obtained in a 74% yield, with fully characterization by 1H nuclear magnetic resonance (NMR), 13C NMR, and mass spectrometry (refer to Supporting information).
Figure 2
Figure 2.
Chemoenzymatic synthesis and characterization of Nb-sTn conjugates. (A) Chemoenzymatic synthesis of oligoglycine-modified sTn antigen and Nbs-sTn conjugates. SDS-PAGE analysis, anti-Myc WB and anti-sTn WB of purified 7D12 and 7D12-sTn conjugate (B), and purified C7b and C7b-sTn conjugate (C). Lane M: marker.
The enzymatic synthesis of Nb-conjugates was performed by the incubation of Nb (20 µmol/L), sTn antigen 9 (500 µmol/L), and enhanced SrtA, (5 µmol/L) in a reaction buffer (50 mmol/L Tris–HCl, 150 mmol/L NaCl, 5 mmol/L CaCl2, pH 7.5) at 16 ℃ for 4 h with 200 rpm shaking. After completion, Ni-IDA magnetic beads were added to remove SrtA and unreacted Nbs. The crude products were purified via ultrafiltration, yielding 7D12-sTn and C7b-sTn conjugates with 70% efficiency. As shown in Figs. 2B and C, all the Nbs and conjugates appeared as a single band around 17 kDa on sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and Western blot (WB) using the anti-myc tag antibodies. These samples were also characterized by WB using anti-sTn mouse serum (MS), clearly, only Nb-sTn conjugates were stained by HRP-conjugated secondary antibodies, confirming the success of sTn modification. The Nbs (Fig. S1 in Supporting information) and the conjugates (Fig. S2 in Supporting information) were further confirmed by mass spectrometry, the results of which were consistent with the calculated molecular weight.
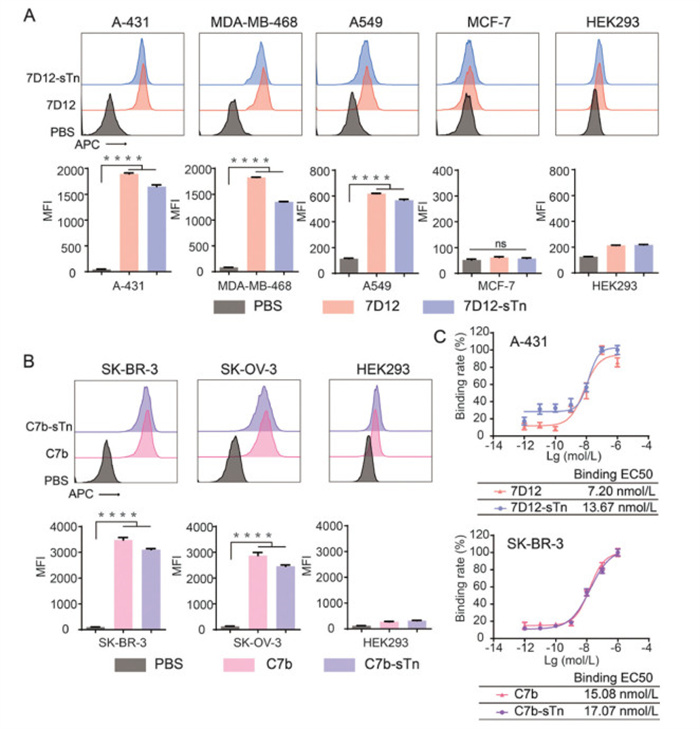
With Nb-sTn conjugates 7D12-sTn 1 and C7b-sTn 2 in hand, we first evaluated their binding profiles to corresponding protein antigens on tumor cells. EGFR-positive cell lines (human squamous carcinoma cell A-431, human triple-negative breast cancer cell MDA-MB-468, and human non-small cell lung cancer A549), HER2-positive cell lines (human breast cancer cell SK-BR-3 and human ovarian cancer cell SK-OV-3), and control cell lines (human breast cancer cell MCF-7, human embryonic kidney cell HEK293), were utilized. The expression of EGFR and HER2 on these cancer cells were confirmed in terms of their RNA expression from the Human Protein Atlas (HPA) database (Fig. S3 in Supporting information), and further verified by flow cytometry using anti-EGFR monoclonal antibody (mAb) Cetuximab, and anti-HER2 mAb Trastuzumab (Fig. S4 in Supporting information). Binding specificity assays were performed by treating cells with phosphate buffered saline (PBS), unmodified Nbs (7D12 or C7b) or Nb-sTn conjugates (7D12-sTn or C7b-sTn), followed by anti-myc tag mouse antibodies and Alexa Fluor 647-conjugated goat anti-mouse IgG antibodies. As shown in Fig. 3A, both 7D12 and 7D12-sTn conjugate could specifically bind to EGFR-positive A-431, MDA-MB-468 and A549 cells, but not EGFR-negative MCF-7 and HEK293 cells. Similarly, C7b and its conjugate only recognize HER2-positive cells (Fig. 3B). To further evaluate the binding affinities of Nb-sTn conjugates, cells were incubated with different concentrations of Nb samples. 7D12-sTn and C7b-sTn conjugates exhibited slightly decreased binding affinities relative to that of the unmodified Nbs, but remained at the nanomolar level at 13.67 and 17.07 nmol/L, respectively (Fig. 3C). These results demonstrated that Nb-sTn conjugates retained good binding abilities to target proteins.
Figure 3
Figure 3.
Evaluation of the binding profiles of Nb-sTn conjugates. Flow cytometry histograms and corresponding MFIs of EGFR-positive (A), and HER2-positive cells (B) treated with PBS, Nbs or Nbs-sTn conjugates, in the presence of anti-myc tag antibodies and Alexa Fluor 647-conjugated goat anti-mouse IgG antibodies. (C) Evaluation of the binding half maximal effective concentration (EC50) values of 7D12, 7D12-sTn conjugate to EGFR-positive A431 cells, and C7b, C7b-sTn conjugate to HER2-posiive SK-BR-3 cells. Data are reported as mean ± SD (n = 3). ****P < 0.0001. ns, no significance.
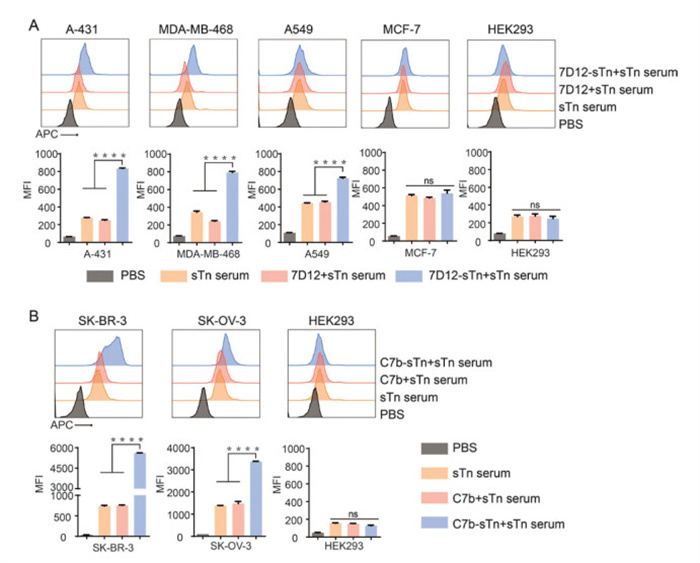
Next, we investigated whether Nb-sTn conjugates could enhance the ability of antibodies produced by the sTn-KLH vaccine to recognize tumor cells. To this end, anti-sTn serum was prepared by immunizing mice with synthetic sTn-KLH, the main component of Theratope vaccine (Fig. S5 in Supporting information). The animal protocol (JN. No. 20230530b0200815) for this study was approved by the Laboratory Animal Center (LAC, No. SYXK(Su)2016–0044) of Jiangnan University. Immune assay results revealed that the MS contained high titer of anti-sTn IgG antibodies (Fig. S6 in Supporting information), which could specifically recognize the sTn positive tumor cells (Fig. S7 in Supporting information). Thereafter, we treated cells with PBS, PBS + anti-sTn MS, unmodified Nbs + anti-sTn MS, or Nb-sTn conjugates + anti-sTn MS, followed by Alexa Fluor 647-conjugated goat anti-mouse IgG antibodies. Flow cytometry was then used to quantitatively assess the levels of anti-sTn antibodies bound to the cell surface. As shown in Fig. 4A, the mean fluorescence intensities (MFIs) of A-431 and MDA-MB-468 cells with high EGFR expression treated with Nb-sTn conjugates were 834 and 793, respectively, representing 2.0- and 1.3-fold higher compared to that of sTn serum groups. In A549 cells with moderate EGFR expression, this treatment also increased the number of anti-sTn antibodies on the cell surface by 60%. The synergistic effect of Nb-sTn conjugate was prominent in HER2-positive SK-BR-3 and SK-OV-3, with dramatically increased MFIs by 6.6- and 1.4-fold higher than sTn serum groups (Fig. 4B). Additionally, no significant enhancement was observed in cells treated with unmodified Nbs or in EGFR or HER2 dual-negative cells across all groups. These results indicated the introduction of Nb-sTn conjugates could significantly increase the number of anti-sTn antibodies on the surface of cells overexpressed with EGFR or HER2. These results were also confirmed by confocal fluorescence images experiments (Fig. S8 in Supporting information). Thus, redirecting more anti-sTn antibodies to tumor cells may hold promise to increase the ability of immune system to recognize and destruct target tumor cells.
Figure 4
Figure 4.
Evaluation of the feasibility of Nb-sTn conjugates to improve the recognition of tumor cells by the generated anti-sTn antibodies. Flow cytometry assays of EGFR-positive (A), and HER2-positive cells (B) treated with PBS + PBS (negative control), PBS + anti-sTn MS, unmodified Nbs + anti-sTn MS, or Nb-sTn conjugates + anti-sTn MS, followed by Alexa Fluor 647-conjugated goat anti-mouse IgG antibodies. Data are reported as mean ± SD (n = 3). ****P < 0.0001.
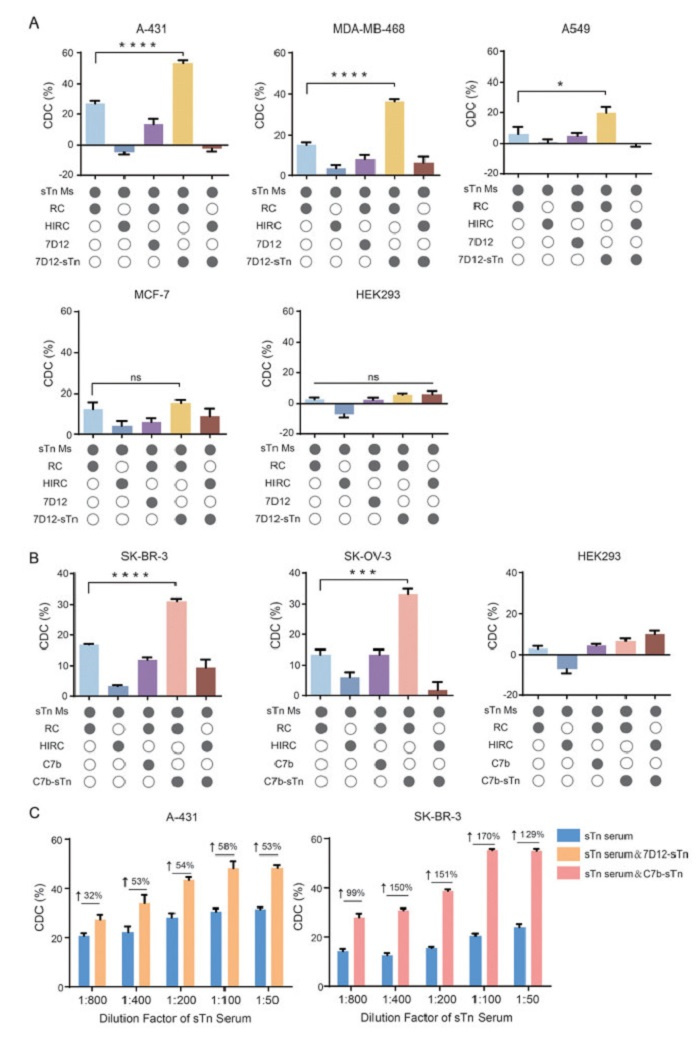
Complement-dependent cytotoxicity (CDC) is a vital mechanism for eradicating tumor cells in TACA-based cancer vaccines. The cytotoxicity of CDC is commonly employed to evaluate the efficacy of these tumor vaccines. To assess the synergistic effects of Nb-sTn conjugates on the antitumor efficacy of the sTn-KLH vaccine, experiments were conducted by incubating cells with 1% anti-sTn serum in the presence of PBS or Nbs or Nb-sTn conjugates, and rabbit complement (RC) at 37 ℃ for 2 h. The cell viability was finally measured using a CCK8 kit on a microplate reader. As shown in Fig. 5A, the addition of 7D12-sTn significantly elevated the cell lysis rates of A-431, MDA-MB-468, and A549 cells to 53.2%, 36.3%, and 19.8%, respectively, compared to the CDC activities of 27.0%, 15.2%, and 6.4% observed with anti-sTn serum alone. The minor cell death was observed in MCF-7 cells (sTn-positive but EGFR-negative), indicating that CDC activity in these cells was solely determined by intrinsic sTn expression. For EGFR-sTn dual-negative HEK293 cells, only < 5% cell lysis was observed, demonstrating the robust dual-target specificity of this strategy and its effectiveness in minimizing potential off-target toxicity. For HER2-targeting models, the synergistic effects of C7b-sTn were also observed, with CDC levels in SK-BR-3 and SK-OV-3 cells increasing from 16.8% to 31.1% and from 13.5% to 33.0%, respectively (Fig. 5B). Likewise, low cell lysis rates were detected in HER2-sTn double-negative HEK293 cells. Additionally, results showed that unmodified Nbs did not exhibit this CDC enhancement capability. No significant cell lysis was observed in all groups when using heat inactivated rabbit complement (HIRC), indicating that the detected cell death was exactly caused by CDC. To further evaluate the enhancement of CDC by Nb-sTn conjugates, A-431 cells and SK-BR-3 cells were selected as cell models. Following the addition of 2% rabbit complement and sTn serum at various dilutions (ranging from 1:800 to 1:50), we analyzed the differences in CDC activity levels in the presence or absence of Nb-sTn conjugates. As shown in Fig. 5C, the synergetic treatment with Nb-sTn conjugates significantly improved the CDC efficacy of the sTn-KLH vaccine. For instance, with the addition of 7D12-sTn, the cell lysis rates of A-431 cells treated with 1:800-, 1:400-, 1:200-, 1:100- and 1:50-diluted sTn serum reached 27.5%, 34.0%, 43.5%, 48.4%, and 48.5%, representing increases of 32%, 53%, 54%, 58% and 53%, respectively. This enhancement was even more pronounced in SK-BR-3 cells treated with the combined regimen of sTn serum and C7b-sTn, where the cell lysis rates were elevated by 99–170% across the tested serum concentrations. In a competitive inhibition assay with varying concentrations of free sTn, both of the CDC toxicities were gradually decreased in anti-sTn serum group and anti-sTn serum plus Nb-sTn groups (Fig. S9 in Supporting information), confirming the sTn-specificity of CDC mediated by anti-sTn serum and Nb-sTn conjugates.
Figure 5
Figure 5.In vitro CDC assays. (A) CDC mediated by 7D12 or 7D12-sTn against EGFR-positive and EGFR negative cells. (B) CDC mediated by C7b or C7b-sTn against HER2-positive and HER2 negative cells. Filled in circles indicate that those conditions were present in the assay arm. (C). Evaluation of the CDC values of serial diluted anti-sTn MS in the presence or absence of Nb-sTn conjugates. Data are reported as mean ± SD (n = 3). *P < 0.05, ***P < 0.001, ****P < 0.0001.
Antibody-dependent cellular cytotoxicity (ADCC) is also a crucial antitumor mechanism of sTn-KLH vaccine. Herein, ADCC activity was preliminarily evaluated by treating the two positive cancer cells (A-431 cells and SK-BR-3 cells as target cells) with Nb-sTn conjugates, unmodified Nbs or PBS in the presence of anti-sTn serum and freshly isolated healthy human peripheral blood mononuclear cells (PBMC, as a source of effector cells). Subsequently, tumor cell death was determined using a commercial lactate dehydrogenase (LDH) release kit. As presented in Fig. S10 (Supporting information), cell groups co-cultured with PBMC did not show apparent cell death. By contrast, significant cell killing activities were observed in groups treated with anti-sTn serum, confirming the ADCC effect of sTn-KLH vaccine. Moreover, significantly higher cancer cell lysis was observed in the groups treated with Nb-sTn conjugates, proving Nb-sTn conjugates could successfully bind to target cancer cells and recruit more anti-sTn antibodies to trigger a stronger ADCC. These results highlighted the advantages of Nb-sTn conjugates in directing anti-sTn antibodies to tumor cells and improving the recognition, thereby boosting the immunity killing activity of sTn-KLH vaccine.
In summary, we developed an integrated strategy that combining vaccination with TACA-based antibody-recruiting molecules that is able to simultaneously redirects anti-TACA antibodies to tumor cells expressing a secondary target, thereby enhancing the efficacy of TACA-based vaccines. Using sTn as a model TACA and EGFR/HER2 as model protein antigens, we designed two Nb-sTn conjugates: 7D12-sTn and C7b-sTn. These conjugates serve as a bridge between protein targets on tumor cell surfaces and circulating anti-sTn antibodies, effectively addressing the challenges posed by the heterogeneity of sTn expression. They significantly enhance CDC and ADCC effect of anti-sTn antibodies, thereby amplifying the therapeutic efficacy of the Theratope vaccine. Given that the heterogeneity of TACA expression on tumor cells is a common issue that adversely impacts the therapeutic outcomes of TACA vaccines, this strategy improves the efficacy of TACA-based vaccines and could facilitate their clinical translation. By combining tumor vaccines with antibody-recruiting molecules, it offers a promising approach to overcome tumor antigen heterogeneity.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
CRediT authorship contribution statement
Dan Li: Investigation, Formal analysis, Data curation. Haofei Hong: Writing – original draft, Formal analysis, Data curation. Han Lin: Methodology, Investigation, Formal analysis, Data curation. Teng Xu: Validation, Investigation, Formal analysis, Data curation. Zhifang Zhou: Validation, Resources, Formal analysis. Zhimeng Wu: Writing – review & editing, Writing – original draft, Supervision, Resources, Methodology, Investigation, Conceptualization.
Acknowledgments
This work was supported by the National Key R&D Program of China (No. 2023YFA0914300) and in partly funded by the Fundamental Research Funds for the Central Universties (No. JUSRP123037) and the 111 Project (No. 111–2–06).
Supplementary materials
Supplementary material associated with this article can be found, in the online version, at doi:10.1016/j.cclet.2025.111324.
U. Flucke, T.K. Zirbes, W. Schröder, et al., Anticancer Res. 21 (2001) 2189–2193.
[28]
A. López-Ferrer, C. Barranco, C. de Bolós, Am. J. Clin. Pathol. 118 (2002) 749–755. doi: 10.1309/LWP3-MFA8-8KX7-60YQ
[29]
S. Julien, P.A. Videira, P. Delannoy, Biomolecules 2 (2012) 435–466. doi: 10.3390/biom2040435
[30]
J. Hung, I. Chen, S. Ueng, et al., J. Immunother. Cancer 10 (2022) e004312. doi: 10.1136/jitc-2021-004312
[31]
A. Zhang, Annu. Rep. Med. Chem. 50 (2017) 481–518.
[32]
H. Hong, H. Lin, D. Li, et al., Angew. Chem. Int. Ed. 61 (2022) e202208773. doi: 10.1002/anie.202208773
[33]
H. Hong, Z. Zhou, K. Zhou, et al., Chem. Sci. 10 (2019) 9331–9338. doi: 10.1039/c9sc03840j
[34]
A. Uvyn, R. De Coen, M. Gruijs, et al., Angew. Chem. Int. Ed. 58 (2019) 12988–12993. doi: 10.1002/anie.201905093
[35]
J. Sianturi, Y. Manabe, H. Li, et al., Angew. Chem. Int. Ed. 58 (2019) 4526–4530. doi: 10.1002/anie.201812914
[36]
I. Shahriar, M. Kamra, A.K. Kanduluru, et al., Proc. Natl. Acad. Sci. U. S. A. 121 (2024) e2408469121.
Figure 1
Illustration of the "TACA-protein antigens dual-targeting concept" to enhance the effect of TACA-based cancer vaccine. (A) Design and construction of Nb-sTn conjugates as a synergistic agent. (B) When compared to sTn-KLH alone, synergetic treatment with Nb-sTn can achieve better efficacy by redirecting more anti-sTn antibodies onto targets.
Figure 2
Chemoenzymatic synthesis and characterization of Nb-sTn conjugates. (A) Chemoenzymatic synthesis of oligoglycine-modified sTn antigen and Nbs-sTn conjugates. SDS-PAGE analysis, anti-Myc WB and anti-sTn WB of purified 7D12 and 7D12-sTn conjugate (B), and purified C7b and C7b-sTn conjugate (C). Lane M: marker.
Figure 3
Evaluation of the binding profiles of Nb-sTn conjugates. Flow cytometry histograms and corresponding MFIs of EGFR-positive (A), and HER2-positive cells (B) treated with PBS, Nbs or Nbs-sTn conjugates, in the presence of anti-myc tag antibodies and Alexa Fluor 647-conjugated goat anti-mouse IgG antibodies. (C) Evaluation of the binding half maximal effective concentration (EC50) values of 7D12, 7D12-sTn conjugate to EGFR-positive A431 cells, and C7b, C7b-sTn conjugate to HER2-posiive SK-BR-3 cells. Data are reported as mean ± SD (n = 3). ****P < 0.0001. ns, no significance.
Figure 4
Evaluation of the feasibility of Nb-sTn conjugates to improve the recognition of tumor cells by the generated anti-sTn antibodies. Flow cytometry assays of EGFR-positive (A), and HER2-positive cells (B) treated with PBS + PBS (negative control), PBS + anti-sTn MS, unmodified Nbs + anti-sTn MS, or Nb-sTn conjugates + anti-sTn MS, followed by Alexa Fluor 647-conjugated goat anti-mouse IgG antibodies. Data are reported as mean ± SD (n = 3). ****P < 0.0001.
Figure 5In vitro CDC assays. (A) CDC mediated by 7D12 or 7D12-sTn against EGFR-positive and EGFR negative cells. (B) CDC mediated by C7b or C7b-sTn against HER2-positive and HER2 negative cells. Filled in circles indicate that those conditions were present in the assay arm. (C). Evaluation of the CDC values of serial diluted anti-sTn MS in the presence or absence of Nb-sTn conjugates. Data are reported as mean ± SD (n = 3). *P < 0.05, ***P < 0.001, ****P < 0.0001.

 DownLoad:
DownLoad:

 下载:
下载:






 下载:
下载:
