Citation:
Peizhang Zhao, Mengmeng Li, Jingwen Wang, Jun Li, Yunfeng Lin. Targeting the immune microenvironment: A novel strategy for treating infected bone defects with hydrogels[J]. Chinese Chemical Letters,
2026, 37(5): 111319.
doi:
10.1016/j.cclet.2025.111319
Targeting the immune microenvironment: A novel strategy for treating infected bone defects with hydrogels
English
Targeting the immune microenvironment: A novel strategy for treating infected bone defects with hydrogels
Orthopedic Research Institute, Department of Orthopedics, West China Hospital, Sichuan University, Chengdu 610041, China
b.
Trauma Center, West China Hospital, Sichuan University, Chengdu 610041, China
c.
State Key Laboratory of Oral Diseases, National Center for Stomatology, National Clinical Research Center for Oral Diseases, West China Hospital of Stomatology, Sichuan University, Chengdu 610041, China
d.
Sichuan Provincial Engineering Research Center of Oral Biomaterials, Chengdu 610041, China
Received Date:
14 September 2024 Accepted Date:
14 May 2025 Revised Date:
08 May 2025 Available Online:
15 May 2026
Abstract:
Infected bone defects (IBD) are intricate and formidable conditions characterized by elevated rates of infection recurrence and delayed healing, resulting from dysregulation of the bone immune microenvironment (IME) mediated by microbes. The conventional approaches including surgical intervention and antibiotic therapy encounter challenges such as antibiotic resistance and susceptibility to postoperative infections. Considering the diverse impacts of various immune cells (ICs) and cytokines, the investigations into the IME have been conducted to offer potential strategies for treating IBD by addressing the requirements of infection eradication and bone regeneration (BREG). However, there is still a lack of review discussing the impacts of IME on IBD in light of its diverse components. Hydrogels, as promising materials in the treatment of IBD, can mimic the extracellular matrix of natural tissues, providing an optimal environment for cell growth and tissue regeneration. Recent studies have focused on investigating immune modulation through hydrogel delivery for treating IBD. This review aims to discuss the effects of different types of ICs and cytokines on the IME in IBD while summarizing current progress and strategies targeting this microenvironment using hydrogels. The insights gained from this review will aid the development of future immunomodulatory approaches for IBD treatment.
IBD are complex and challenging conditions that significantly impact orthopedic treatment. As a major form of bone infection, osteomyelitis represents a substantial healthcare burden with an estimated incidence of 22 cases per 100,000 person-years in the United States. Concurrently, fracture-associated infections (FRI) demonstrate significant variability in occurrence rates, with closed fractures showing 2% incidence compared to open fractures reaching 50% infection rates [1]. These defects not only result in delayed healing but also have high rates of infection recurrence, posing a series of challenges that severely affect patients' quality of life [2,3]. One major issue with IBD is the delayed healing process. The presence of infection within the bone hinders the body's natural ability to repair itself, leading to prolonged recovery times for patients. This delay can be frustrating and debilitating, as it limits mobility and functionality, affecting daily activities such as walking or even simple tasks like picking up objects. Moreover, the high recurrence rates of infections in IBD further complicate treatment outcomes. Despite initial interventions and antibiotic therapies, infections may persist or reoccur due to various factors such as bacterial resistance or inadequate surgical debridement. This recurrent cycle not only prolongs the healing process but also increases the risk of complications and potential long-term damage to surrounding tissues [4,5]. In addition to physical challenges, IBD can have a significant psychological impact on patients. Living with chronic pain, limited mobility, and uncertainty about future outcomes can lead to feelings of frustration, anxiety, and depression. Patients may experience a loss of independence and reduced participation in social activities due to their condition's limitations [6]. Furthermore, managing IBD requires comprehensive multidisciplinary approaches involving orthopedic surgeons, infectious disease specialists, wound care experts, physical therapists, and other healthcare professionals. Coordinating these different specialties can be complex and time-consuming for both medical teams and patients alike.
The current clinical treatment standard focuses on controlling infection and promoting bone regeneration (BREG) [7]. Infection-induced inflammatory reactions generate a complex immune microenvironment (IME). While this microenvironment aids pathogen clearance, it paradoxically hinders the formation of an osteogenic microenvironment and BREG [8]. Therefore, it is common practice in clinical settings to combine antibiotics with bone grafting to address these intricate infections and facilitate BREG [9]. However, traditional multi-stage treatment strategies have led to increased bacterial resistance and prolonged treatment durations that burden patients [10]. Furthermore, conventional approaches fail to effectively balance the objectives of eradicating infection and suppressing inflammation while promoting BREG [11]. Consequently, there is an urgent and compelling demand for a comprehensive treatment strategy that exhibits potent antimicrobial properties and robust osteoinductive abilities throughout bone repair (BREP) of IBD therapy.
The relationship between the immune system and bone reconstruction has been extensively investigated in recent years. It is now widely acknowledged that a moderate immune response is crucial for proper bone healing, while an excessive or dysregulated response can impede regeneration. This concept of osteoimmunology has emerged as a novel field of research, focusing on the interactions between the skeletal and immune systems [12]. Numerous molecules participate in this crosstalk between the two systems, including cytokines, receptors, signaling pathways, and transcription factors [13]. These molecules can exhibit both pro-inflammatory and anti-inflammatory effects depending on their context and concentration levels. For instance, certain cytokines such as tumor necrosis factor (TNF)-α or interleukin (IL)-1β are recognized to promote inflammation but also stimulate bone resorption when present at elevated concentrations [14,15]. Apart from its role in bone healing, the immune system also plays a pivotal role in combating infections and eliminating pathogens from the body [16]. However, when this response becomes dysregulated or chronic (as observed in IBD), it can result in systemic inflammation affecting multiple organs including bones [17]. Therefore, there is a growing interest in developing strategies aimed at modulating the IME during bone healing processes [18]. This may involve utilizing immunomodulatory drugs or cell-based therapies targeting specific components of the immune system while promoting osteogenesis. By doing so, we may potentially achieve more natural and functional outcomes for BREP with fewer complications or side effects compared to current treatments allow for.
Hydrogels have emerged as promising biomaterials for the treatment of bone defects due to their unique properties. They are highly biocompatible and can mimic the extracellular matrix (ECM) of natural tissues, providing an ideal environment for cell growth and tissue regeneration [19]. Moreover, hydrogels can be engineered to release therapeutic agents in a controlled manner, allowing for localized drug delivery directly at the site of infection [8]. One key aspect that this paper focuses on is targeting the IME within IBD. The immune system plays a crucial role in combating infections and promoting tissue repair. However, in cases of chronic infections or compromised immune responses, it may not effectively eliminate pathogens or support proper healing processes [11]. To overcome these challenges, researchers have developed hydrogel-based systems that modulate the immune response within IBD. By incorporating immunomodulatory factors into hydrogel matrices, these strategies aim to enhance antimicrobial activity while promoting tissue regeneration simultaneously [20,21]. Furthermore, this paper will discuss different types of hydrogels used for treating infectious bone defects and highlight their advantages and limitations. It will also delve into specific techniques employed to optimize drug release kinetics from hydrogel systems and ensure sustained therapeutic effects over extended periods. Overall, this comprehensive review aims to provide insights into cutting-edge strategies utilizing hydrogels targeted at the IME for effective therapeutic interventions.
2.
Role of immune cells (ICs) in IBD
2.1
Neutrophils
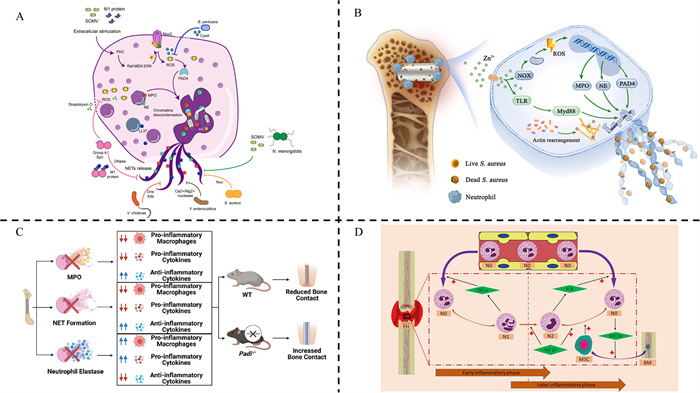
Neutrophils are derived from hematopoietic stem cells (HSCs) in the bone marrow and, upon differentiation and maturation, they are released into the bloodstream [22]. In the treatment of IBD, neutrophils play a pivotal role by simultaneously addressing infection control and bone defect repair. The latter encompasses an inflammatory response (IR) as well as processes of BREG and remodeling [7]. During the IR in IBD, neutrophils rapidly migrate to inflammatory sites upon chemotactic stimulation, playing a critical role in pathogen containment [23]. Activated neutrophils exert antimicrobial defense through the release of factors, direct phagocytosis, and secretion of neutrophil extracellular traps (NETs)-weblike DNA structures adorned with antimicrobial proteins. These NETs efficiently capture and eliminate bacteria after appropriate activation by lipopolysaccharide (LPS) and other factors (Fig. 1A) [24]. Furthermore, neutrophils can secrete various cytokines such as TNF-α, IL-1β, IL-6, and IL-10 to recruit macrophages along with other ICs. Apart from their ability to clear infections, neutrophils can have both beneficial and detrimental effects during the process of BREG. On one hand, they secrete vascular endothelial growth factors (VEGF) and facilitate the formation of fibronectin and fibrin, which support the recruitment and attachment of cells in bone fracture healing [25,26]. Additionally, the absence of neutrophils disrupts BREG [27]. On the other hand, neutrophils may also play a negative role. It has been reported that bacterial LPS-stimulated neutrophils express high levels of receptor activator of nuclear factor-κB ligand (RANKL), which can activate osteoclasts leading to rapid bone loss [28]. Recent studies have revealed that neutrophils can be divided into two phenotypes (N1 and N2) providing different perspectives to explain these controversial mechanisms. N1 neutrophils are known to promote IRs by secreting cytokines such as TNF-α, interferon (IFN)-γ, IL-6, monocyte chemoattractant protein (MCP)-1, and macrophage inflammatory protein (MIP)-2 to activate and recruit various ICs while regulating polarization, while N2 neutrophils were capable of secreting anti-inflammatory factors, inducing the polarization of M2 macrophages and promoting BREG [29,30]. Exposure to transforming growth factor (TGF)-β, granulocyte-colony stimulating factor (G-CSF), proper concentration of IL-8 will lead to a polarization towards N2 neutrophils; on the contrary inhibiting TGF-β signaling pathway or imposing IFN-γ causes N1-polarization in neutrophil phenotype [29,30]. During the early inflammatory phase in BREP, macrophages recruited by N1 neutrophils which could be typical proinflammatory LPS-activated are M1-polarized to clear the infection, tissue debris, preparing for the next stage: BREG in this proinflammatory microenvironment [29]. In contrast, N2 neutrophils possess abilities inducing anti‐inflammatory phenotypes in macrophages, CD4+ T cells as well as recruiting bone marrow-derived mesenchymal stem cells (BMSCs) during later BREG stages where stromal cell-derived factor (SDF)‐1/C-X-C chemokine receptor type 4 (CXCR4) axis along with angiotensin-converting enzyme (ACE)/nitric oxide synthase 1 (NOS1) could play roles [29,31].
Figure 1
Figure 1.
(A) The NETs effectively eliminate bacterial infections and bacteria employ virulence factors to evade NETs release during infection. Reproduced with permission [24]. Copyright 2022, Frontiers. (B) The presence of zinc ion (Zn2+) enhances the generation of ROS-dependent NETs and facilitates osseointegration in rat femur infection. Reproduced with permission [32]. Copyright 2023, KeAi Communication. (C) The formation of NETs is crucial for activating pro-inflammatory macrophages. In contrast, inhibiting NET formation reduces this pro-inflammatory response, which in turn enhances bone formation around implanted biomaterials in non-infectious environments. Reproduced with permission [33]. Copyright 2023, Elsevier. (D) The neutrophils differentiate into different phenotypes in the different phases of BREP. Reproduced with permission [34]. Copyright 2024, Elsevier.
The effects of NETs could also contribute to the explanation of controversial effects of neutrophils. The contradictory effects of NETs on healing of IBD can be attributed to their dual roles in infection and inflammation. The NETs are crucial for the clearance of infections, which is important for the early inflammatory acute infection phase and chronic bacterial infections in BREG of IBD (Fig. 1B) [32]. In the meantime, the NETs activated in incentive inflammatory environment may result in the sustained activation of the IR and inhibit osteogenesis by osteoblast; activate bone resorption by osteoclast through inducing RANKL expression in osteogenic cells and promoting osteoclast differentiation and function [35,36]. Reducing the presence of NETs not only alleviates inflammation but also mitigates the pro-inflammatory phenotype exhibited by macrophages in the BREG stage (Fig. 1C) [33]. The systematic relationships between N1/N2 neutrophils and different ICs in IBD remain unclear, while the effects of NETs with varying concentrations, structures, and modes of activation on the regulation of bone defects remain ambiguous [34,37]. However, an appropriate IR can promote osteogenesis; conversely, excessive inflammation caused by infection has detrimental effects on BREG [7]. The remarkable capability of NETs and N1/N2 neutrophils to inhibit infections and regulate inflammations offers a promising perspective for healing IBD from the standpoint of infection and inflammation (Fig. 1D) [34].
2.2
Macrophages
The macrophages are integral components of the mononuclear phagocytic cell system and originate from HSCs [38]. The predominant M1 phenotype during the early stage of inflammation and its transition to the M2 phenotype during the later stage of BREG play a pivotal role in regulating the inflammatory microenvironment and facilitating osteogenesis for effective healing IBD (Fig. S1A in Supporting information) [39,40]. In the early inflammatory phase of acute bacterial infections and initial process of BREG, myeloid ICs including macrophages are recruited by neutrophils (Fig. S1B in Supporting information) [41]. The M1 polarization of macrophages can be induced by direct stimulation from bacterial products and inflammatory molecules such as TNF-α, IFN-γ, and granulocyte-macrophage colony-stimulating factor (GM-CSF), thereby enhancing their bactericidal and phagocytic capabilities [11,42]. The formation of biofilms in methicillin-resistant Staphylococcus aureus (MRSA) can induce the transition of M1 macrophages to M2 macrophages, thereby attenuating the phagocytic and bactericidal functions of macrophages [43]. The M1 macrophages also secrete pro-inflammatory cytokines, including TNF-α, IL-1, IL-2, IL-6, IL-8, IL-12, and IL-23, to effectively impede pathogen growth and clear infections; however, excessive secretion of these pro-inflammatory cytokines can result in bone tissue damage [11,19,37,44]. Other than the ability to control infections, M1 macrophages also release high levels of VEGF to promote angiogenesis during the early inflammatory phase [45]. Meanwhile, in the process of BREG, the conversion of the M1 phenotype to the M2 phenotype is essential to regulate the inflammatory microenvironment and promote osteogenic differentiation for BREG [42,46]. Although promoting M1 macrophage polarization can eliminate persistent bacteria and clear chronic infections during BREG, sustained activation of M1 macrophages may induce an excessive inflammatory microenvironment, leading to bone damage and delayed BREG. Moreover, M1 macrophages possess the capability to inhibit osteoblast formation and function while stimulating osteoclast formation and activation through the RANKL/receptor activator of nuclear factor-κB (RANK)/osteoprotegerin (OPG) signaling pathway mediated by pro-inflammatory cytokines, particularly TNF-α and IL-6 [44]. On the contrary, M2 phenotypes, which can be activated by IL-4 and IL-13 or N2 neutrophils, have the ability to release anti-inflammatory cytokines such as IL-10 and IL-4 in order to counteract the side effects induced by pro-inflammatory cytokines [47]. The M2 macrophages also secrete a range of cytokines, such as TGF-β, insulin-like growth factor (IGF)-β, VEGF, bone morphogenetic protein (BMP)-2, and BMP-4, to facilitate the processes of vascularization and mineralization in BREG. This is accomplished by inducing the differentiation and proliferation of undifferentiated mesenchymal stem cells (MSCs) into osteoblasts (OBs), promoting osteogenic differentiation in BMSCs through the upregulation of osteogenic genes including recombinant runt related transcription factor 2 (RUNX-2), osteocalcin (OCN) and osteopontin (OPN), and inhibiting the differentiation of osteoclast precursors into osteoclasts [41,44,48,49].
Apart from secreting various cytokines and kinases. M1/M2 macrophages also assemble extracellular vesicles (EVs) to regulate the inflammatory microenvironment and BREG [50]. Extracellular vesicles, which have the ability for communication between cells and are classified into exosomes, micro-vesicles, microparticles by specific mechanisms, are membrane-enclosed vesicles derived from cells containing proteins, lipids, and RNAs from the parent cell and play a similar role as their parental cells [51]. The small-sized EVs (sEVs) derived from M1 macrophages were observed to augment the expression of RANKL and facilitate osteoclastogenesis [51]. In contrast, exosomes derived from M2 macrophages were found to modulate the polarization of M1 macrophages towards an M2 phenotype through the phosphatidylinositol 3-kinase (PI3K) pathways and promote the fracture healing by regulating the inflammatory and immune microenvironment [52]. Another experiment demonstrated that M2-derived exosomes enhanced osteogenesis and angiogenesis, as well as stimulated proliferation of human mesenchymal stem cells (HMSCs) and human umbilical vein endothelial cells (HUVECs) [53]. Furthermore, it was shown that M2-exosomes can promote osteogenic differentiation of BMSCs through various mechanisms, including upregulating BMP-2 expression, activating insulin receptor substrate-1 (IRS-1)/transcriptional coactivator with PDZ-binding motif (TAZ) signaling, promoting the expression of alkaline phosphatase (ALP), RUNX-2, OPN, and IL-10, and inhibiting salt-inducible kinase (SIK)-2 and SIK-3 via various cargo molecules [54]. However, further comprehensive analysis is required to investigate the effects of EVs on BREG at different stages. Meanwhile, the identification of biomarkers that can distinguish M1-exosomes from M2-exosomes remains unclear [53].
2.3
Dendritic cells (DCs)
DCs, which are professional antigen-presenting cells that initiate both innate and adaptive immune responses, play a crucial role in immune regulation. During the early and middle stages of IR, they often mature and aggregate at the site of damage [55]. In the process of anti-infection, DCs bridge the innate and adaptive immune systems through their regulation of T cells, which involves three signals [56]. Firstly, immature DCs are activated by pattern recognition receptors (PRRs) upon microbial stimulation. The antigens recognized by DCs are then presented to T cells along with major histocompatibility complex class Ⅱ (MHC-Ⅱ). Secondly, DCs upregulate the expression of cluster of differentiation (CD80)/CD86/CD40 to activate T cells. Thirdly, DCs secrete cytokines such as TNF-α, IL-1β, TGF-β, IL-10 to promote the differentiation of T cells into T helper (Th) 1, Th2, Th17, Th9 cells and regulatory T cells (Treg) subsets, in order to induce or inhibit IRs (Fig. S1C in Supporting information) [57]. In the process of BREG, exosomes derived from mature DCs have been discovered to possess the capability of enhancing osteogenic differentiation of BMSCs. This is achieved by suppressing large tumor suppressor kinase 1 (LATS1) expression and inhibiting Hippo signaling [58]. Further investigation has indicated that activated DCs can contribute to MSCs recruitment but do not significantly affect MSCs osteogenic differentiation [55]. Moreover, under the induction of macrophage colony-stimulating factor (M-CSF) and RANKL, GM-CSF or CD4+ T cells, DCs can be induced to differentiate into osteoclasts to directly regulate BREG [59,60]. Meanwhile, DCs indirectly impact BREG by regulating T cell subsets including Th1, Th17, and Treg.
2.4
Natural killer (NK) cells
NK cells are a crucial innate ICs type that possess the ability to induce an adaptive immune response [61]. NK cells primarily participate in the early inflammatory phase, where they play a pro-inflammatory role [62]. In addition to their capability for antibody-dependent cellular cytotoxicity (ADCC), NK cells also exhibit cytokine production capabilities, including IFN-γ and TNF-α (Fig. S1D in Supporting information) [63]. During the process of bone generation, it has been reported that NK cells can express M-CSF and RANKL to promote osteoclastogenesis in rheumatoid arthritis (RA). These osteoclasts are capable of bone resorption only in the presence of IL-15 [64]. Meanwhile, NK cells can also induce osteoclast apoptosis through high levels of expression of TNF-related apoptosis-inducing ligands [63].
2.5
B cells
B cells are derived from HSCs in the bone marrow and are well recognized as essential components of the adaptive immune system due to their ability to produce and secrete antibodies [59]. During early IRs, they differentiate into plasma cells that generate antibodies against pathogens. Additionally, activated B cells express high levels of TGF-β, which has been found to inhibit osteoblast activity during periodontal inflammation states [65]. Moreover, B cells with inflammatory phenotypes can exhibit elevated levels of RANKL, TNF, and G-CSF under an inflammatory microenvironment, thereby promoting proliferation of osteoclast progenitors [66,67]. B cells possess the ability to modulate the cytokine milieu towards OPG expression while concurrently suppressing RANKL production, thereby exerting inhibitory effects on osteoclasts and facilitating bone formation during the intermediate-to-late stages of BREG when B cells and plasma cells exhibit heightened levels of OPG expression (Fig. S2A in Supporting information) [66]. Conversely, it has been reported that both B cells and plasma cells derived from B cells can express RANKL and IL-7 to induce osteoclastogenesis and facilitate ovariectomy-induced bone loss [38,68]. Furthermore, B cells possess the capability to modulate osteoclast activity by inhibiting their function under Th1 cell activation, while promoting osteoclastogenesis under Th2 cell activation, thereby exerting regulatory effects on BREG [69].
Apart from traditional antibody-secreting B cells, regulatory B cells (Bregs) subsets known for their anti-inflammatory properties also contribute to the process of BREG by preventing excessive inflammation following infection [70]. Upon activation during bone formation, Bregs secrete anti-inflammatory cytokines including IL-10 and TGF-β to suppress differentiation and functions of osteoclasts while promoting differentiation of Treg cells and establishing an anti-inflammatory microenvironment [71]. The functions of B cells are closely associated with the inflammatory microenvironment, where their changes likely play a crucial and flexible role. The inflammatory microenvironment in IBD exhibits flexibility in order to clear infections and promote BREG [7]. The functions of B cells have significant and distinct impacts during different phases of IBD by directly regulating BREG through the RANKL/OPG axis (Fig. S2B in Supporting information) [70]. Additionally, interactions between T cells and B cells, as well as estrogen levels, may also influence the functions of B cells [38]. However, there is currently a lack of systematic analysis on the functions of B cells in different phases of IBD with varying inflammatory and immune microenvironments. Existing evidence suggests that a pro-inflammatory microenvironment may induce osteoclastogenesis by B cells while an anti-inflammatory microenvironment may lead to bone formation function for these same cells in IBD. Furthermore, the anti-inflammatory properties and ability to inhibit bone resorption exhibited by Bregs make them promising candidates for promoting BREG in IBD at the frontiers of osteoimmunology research [70].
2.6
T cells
T cells originate from the bone marrow and undergo development and maturation in the thymus, playing a crucial role in both innate and adaptive immune responses. The functional diversity of T cells is determined by their differentiation state, which can be categorized into two groups based on distinct T cell receptors (TCRs): γδ T cells and αβ T cells [59]. Unlike αβ T cells, γδ T cells do not require MHC-mediated antigen presentation; instead, they recognize and respond to infectious cells through multiple receptor-ligand interactions similar to innate immune responses. Within the αβ T cell population, helper T cells expressing CD4 receptors and cytotoxic T cells (Tc) expressing CD8 receptors are key players during the early IR phase as well as BREG phase [38,72]. Helper T cells differentiate into Th1, Th2, Th9, Th17, Th22 subtypes while cytotoxic T cells differentiate into subsets such as Tc1, Tc2, Tc9, Tc17, Tc22 [72]. The functions of different subsets of these specialized ICs populations are closely associated with controlling infections and promoting BREG of IBD treatment [59].
2.6.1
γδ T cells
γδ T cells are predominantly found in peripheral tissues, such as mucosal tissues, skin, and lungs [73]. Residing in musculoskeletal tissues, γδ T cells are rapidly activated by damage-associated molecules and stress-induced self-antigens released during infection and bone defects [74]. Upon activation, these cells produce abundant cytokines that play a crucial role in early tissue inflammation regulation [74,75]. During infection-induced early IR, γδ T cells can enhance neutrophil recruitment and facilitate bacterial clearance in S. aureus infections [76]. Activated by viral antigens, γδ T cells express pro-inflammatory cytokines including IFN-γ and TNF-α to induce inflammation and eliminate the pathogen [77]. Recent research has found that γδ T cells can directly interact with macrophages to skew them towards an M1 phenotype and delay the switch to anti-inflammatory M2 activation during skin healing through IFN-γ production [75]. In addition to their ability to control infections, IL-17-producing γδ T cells were also found to promote osteoblastogenesis of MSCs in fracture healing [74]. Further studies have indicated that the majority of IL-17-producing γδ T cells implicated in fracture repair originate locally and are involved in the early stages of BREG with an association with the intestinal microbiome [78]. Additionally, γδ T cells can inhibit osteoclastogenesis and osteoclast function by downregulating RANK expression to suppress the RANKL pathway and inhibit bone resorption [79]. However, there is still limited understanding regarding how γδ T cell functions relate to anti-infection mechanisms and promoting BREG specifically within IBD.
2.6.2
CD8+ T cells
In addition to the rapid responses mediated by γδ T cells, the adaptive arm of the immune system further relies on CD8+ T cells, which play a key role in eliminating infected cells through cytotoxic mechanisms. CD8+ T cells are widely known as cytotoxic T cells that kill cells infected with intracellular pathogens through Fas cell surface death receptor ligand (FasL), perforin and granzymes. During early acute IRs induced by infection, they also produce pro-inflammatory cytokines including IFN-γ and IL-2 [80,81]. In the process of BREG, it has been observed that CD8+ T cells are capable of producing IFN-γ and actively participate in inflammatory processes. Depletion in the number of CD8+ T cells has shown to enhance bone healing in the absence of infection [82]. Further study found that patients with impaired fracture healing have elevated levels of effector CD8+T cells in peripheral blood [83]. In contrast, eliminating CD8+ T cells with OX8 monoclonal antibody (CD8 antibodies) was found to hinder cancellous bone healing in terms of implant fixation which could be the impacts of experimental models [19,84].
2.6.3
Th1 and Th2 cells
Th1 and Th2 cells are two subsets of CD4+ T cells that undergo polarization upon antigen stimulations [59]. Th1 cells produce pro-inflammatory cytokines, including IFN-γ, TNF-α, and IL-12, to eliminate pathogens primarily by activating macrophages [38]. They also contribute to the induction of osteoclastogenesis and promotion of fibrogenesis [85]. Meanwhile, Th2 cells secrete IL-4, IL-10, and IL-13 and mainly interact with B lymphocytes, mast cells, and granulocytes to enhance humoral immunity against extracellular pathogens while preventing bone destruction and promoting BREG [80,85,86]. It has been observed that Th2 cells can directly or indirectly inhibit RANKL-induced osteoclastogenesis through the action of IL-4 and IL-13 (Fig. S2C in Supporting information) [80]. Additionally, IL-4 can induce naive T cells to polarize towards the Th2 phenotype which secretes elevated level of IL-4 while inhibiting the generation of Th1 cells and IFN-γ production which creates an anti-inflammatory microenvironment conducive for BREG [85,87].
2.6.4
Th17 and Treg cells
Th17 cells are a subset of CD4+ T cells characterized by their robust secretion of IL-17 [88]. Polarization towards Th17 cells can be induced by various factors, including IL-6, inflammation, and recognition signals from Toll-like receptors (TLRs) upon bacterial encounter [89]. Reacting to infections, Th17 cells possess the ability to recruit B cells via C-X-C motif chemokine ligand 13 (CXCL13) and stimulate antibody production for effective pathogen clearance [90]. Additionally, they secrete cytokines such as IL-21, IL-22, and TNF-α to further enhance inflammation and facilitate ICs activation and recruitment [80]. The production of IL-17 by Th17 cells is capable of inducing osteoclastogenesis through RANKL induction [38]. Moreover, Th17 cells have the capability to directly secrete RANKL or indirectly release pro-inflammatory cytokines such as TNF-α and IL-1, thereby initiating osteoclastogenesis and promoting bone destruction [89]. On the contrary, CD4+ Treg cells exert opposing effects on Th17 cells [38]. Treg cells can suppress Th1 and Th17 responses through the expression of T-bet and retinoid-associated orphan receptor γt (RORγt) [90]. The substantial secretion of TGF-β and IL-10 by Treg can concurrently suppress both adaptive and innate immunity while directly inhibiting osteoclastogenesis [80]. Furthermore, Treg cells can hinder inflammation and osteoclastogenesis by producing cytotoxic T-lymphocyte antigen (CTLA)-4 and interacting with CD80/86 on pre-osteoclast cells [12]. In addition, newly research found that Treg can be recruited by C–C motif chemokine ligand 3 (CCL3) and CCL17 secreted by macrophages and play a crucial role in BREG mediated by B-cell progenitors [91]. In summary, Treg primarily functions as an anti-inflammatory agent while promoting BREG during the process of IBD. The balance between Th17 and Treg cells plays a critical role in clearing infections while suppressing IRs to facilitate BREG (Fig. S2D in Supporting information) [92]. Transitioning to a state of Treg may help maintain immune tolerance and promote bone reconstruction in an inflammatory microenvironment [89,91,93,94].
3.
Role of immune cytokines in IBD
The pathogenesis of IBD encompasses intricate interactions between immune dysregulation and tissue remodeling, wherein cytokines serve dual roles in mediating inflammation and regulating bone metabolism. TNF-α promotes inflammation through TNF receptor (TNFR)-1/2 signaling pathways while exerting concentration-dependent effects on both osteogenesis and osteoclastogenesis. IL-1 facilitates the clearance of infections but concurrently exacerbates bone resorption via RANKL and prostaglandin E2 pathways. IFN-γ exhibits paradoxical effects by directly inhibiting osteoclastogenesis while amplifying inflammation-induced bone loss. In contrast, anti-inflammatory cytokines such as IL-4 and IL-10 mitigate inflammation by polarizing macrophages toward an M2 phenotype and suppressing osteoclast differentiation; however, these actions may potentially compromise host antimicrobial responses. Collectively, these cytokines modulate the immune-bone axis in IBD, with infection status, cytokine concentration, and microenvironmental cues collectively determining their net impact on disease progression and skeletal complications. The supplementary materials provide a comprehensive discussion of these mechanisms, highlighting their therapeutic implications for the management of IBD.
4.
Strategies for direct regulation of ICs via hydrogels
4.1
Regulating neutrophils N1-N2 polarization and NETs formation
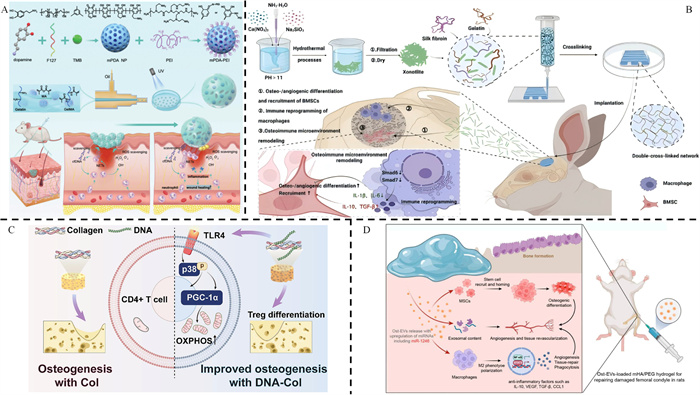
Neutrophils play a crucial role in the process of combating infections, regulating inflammation, and promoting bone reconstruction. An Ozone-delivering hydrogel has been shown to effectively combat localized S. aureus infections in acute osteomyelitis by promoting the generation of reactive oxygen species (ROS) and NETs, while reducing neutrophil infiltration. This suggests a potential therapeutic effect of enhancing NETs generation to upregulate neutrophil functions for anti-acute infection in IBD [95]. Another study developed a “micro cage” scavenger that carries functionalized mesoporous polydopamine (mPDA)-cationic polyethyleneimine (PEI) to capture NETs in chronic inflammatory cascades, leading to the clearance of NETs and suppression of neutrophil polarization towards N1 phenotype, thereby reducing excessive inflammation [96]. Moreover, it has been observed that the polarization of N2 neutrophils can induce M2 macrophage polarization, which helps suppress excessive inflammation and promote BREG [97].
The strategy of promoting NETs formation by neutrophils for early bacterial clearance and inducing their polarization towards an anti-inflammatory N2 phenotype during later stages of bone reconstruction shows significant promise as it can regulate the IME to both clear infections and induce BREG in typical cases. For patients with excessive inflammation, a novel bio-based hydrogel microspheres “micro-cage” has been developed, aiming to effectively clear the overexpression of NETs and facilitate wound healing (Fig. 2A) [96]. However, impaired capability of forming NETs may result in reduced ability to fight off infections [98]. Additionally, it has been reported that the DNA released in NETs can enhance Treg differentiation through the promotion of mitochondrial oxidative phosphorylation (OXPHOS) in CD4+ T cells [99]. Therefore, there is still an urgent need for a deeper understanding of the complex interactions between neutrophils, pathogens, the immune system, and skeletal system [34,98].
Figure 2
Figure 2.
(A) A ’micro-cage’ composed of mPDA-PEI@GelMA that clears the NETs in the damaged area to suppress inflammation. Reproduced with permission [96]. Copyright 2024, Wiley. (B) The 3D-printed silk fibroin/gelatin basal scaffold, loaded with xonotlite nanofiber (Ca6(Si6O17) (OH)2, CS), was observed to effectively modulate the polarization of macrophages towards the M2 phenotype. Reproduced with permission [39]. Copyright 2024, Springer Nature. (C) The DNA-Col scaffold, crosslinked with deoxyribonucleic acid, induced the recruitment of Tregs to enhance bone defect healing. Reproduced with permission [94]. Copyright 2023, Elsevier. (D) The composite system facilitates the transportation of EVs to enhance bone tissue regeneration. Reproduced with permission [100]. Copyright 2024, American Chemical Society.
4.2
Regulating macrophages M1-M2 polarization based on different states
Different strategies have been proposed to regulate macrophages in order to enhance the healing process of IBD. Emerging research indicates that promoting M2 polarization of macrophages during the early stage of acute infection can suppress excessive inflammation, accelerate tissue repair, while in chronic infection, M1 polarization can aid in clearing biofilms and intracellular bacteria to prevent persistent and recurrent bacterial infections [11]. In an acute infection model, it has been reported that a nanosilver/halloysite nanotubes/gelatin methacrylate (nAg/HNTs/GelMA) hybrid hydrogel can promote M2 polarization, inhibiting the inflammatory and cytotoxic microenvironment caused by infection while facilitating new bone formation [101]. A composite hydrogel-metal scaffold was designed for delivering IL-4 and BMP-2 to promote macrophage polarization towards the M2 phenotype, accelerating BREG. Additionally, this scaffold possesses inherent antibacterial capability through quaternary chitosan to clear infections [102]. Yang et al. developed a three-dimensional (3D)-printed scaffold composed of silk fibroin/gelatin (Gel) and xonotlite nanofibers, which has the potential to induce immune reprogramming in macrophages through the TGF-β signaling pathway (Fig. 2B) [39]. In a chronic infection microenvironment where bacteria form biofilms and intracellular bacteria protect themselves from antibiotic killing, promoting M1 polarization is beneficial for pathogen clearance [11]. However, enhancing the M1 phenotype may improve its antibacterial activity but impair its ability for bone induction while promoting the M2 phenotype may facilitate BREG but compromises immune function during chronic infection. Therefore, a potential solution could be promoting M2 polarization of macrophages while preserving their phagocytic function [103]. It has been discovered that gold nanoparticles with a diameter of 50 nm (Au50) can induce M2 polarization in macrophages while also enhancing clearance of intracellular bacteria through overexpression of triggering receptor expressed on myeloid cells 2 (TREM2) [103]. On the contrary, it has been discovered that inducing M1 polarization during the early stages of acute infection can enhance bacterial eradication and prevent recurrent infections, while promoting M2 polarization in later phases can facilitate angiogenesis and tissue repair, thereby significantly impacting the treatment of MRSA-infected acute osteomyelitis [104]. Researchers developed injectable microspheres composed of an interpenetrating network of ionic-cross-linked sodium alginate (SA) and genipin (Gp)-cross-linked Gel, incorporating tannic acid (TA) and copper ions (Cu2+). These microspheres effectively polarize macrophages towards a pro-inflammatory M1 phenotype during early infection, subsequently promoting an anti-inflammatory M2 phenotypic microenvironment after successful bacterial elimination to support tissue repairment [104].
The regulation of macrophage polarization between M1 and M2 types is closely related to the patient's condition. For instance, diabetic patients produce excessive ROS due to hyperglycemia. This drives a shift toward the M1 phenotype and disrupts the balance between M1 and M2 polarization [105]. Therefore, promoting M2 polarization of macrophages can aid in suppressing excessive inflammation, eliminating bacterial infections, and promoting osteogenesis for diabetic patients. The incorporation of wild type macrophages into collagen hydrogels enhanced BREG in diabetic wounds, potentially through modulation of M1/M2 ratios and an increase in the population of M2 macrophages [106]. Scientists have developed a DNA-based hydrogel that carries silver nanoclusters (AgNCs) and extracellular vesicles derived from M2 macrophages (M2EVs), which not only promotes BREP but also exhibits antibacterial and anti-inflammatory properties through inducing macrophage polarization towards the M2 phenotype in diabetic condition [107]. The thermosensitive composite hydrogel PF-127@DOX/ALG-MS@PA was discovered to effectively suppress inflammation and bone resorption by modulating macrophage polarization towards the M2 phenotype, through its staged release of proanthocyanidin (PA) [108].
4.3
Regulating B lymphocytes in IBD
B lymphocytes play a flexible role in IBD through its regulation on OPG/RANKL axis. Scientists designed hydrogel microbeads encapsulated with mangiferin (MAG) for antibacterial and cytosine-phosphate-guanosine oligodeoxynucleotides (CpG ODN) which is considered as a strong B cell activator that activates Breg cells. The research reveals that the hydrogel activating Breg cells can inhibit RANKL expression on day 4 and inhibit osteoclast differentiation while reducing expressions of pro-inflammatory cytokines including IL-1β to promote alveolar BREG [109]. The effects of B lymphocytes are closely related to its subsets as well as inflammation while types of infections also impact on the activation and functions of B lymphocytes [110]. However, the research on the effects of hydrogel regulating B lymphocytes in treating IBD is still relatively limited, and practical applications require further investigation.
4.4
Regulating T lymphocytes in IBD
An artesunate-loaded thermosensitive chitosan hydrogel was found to suppress inflammation and promote BREG by increasing the proportion of Th2 cells while reducing Th1 cells, thereby promoting the expression of OPG and ALP and downregulating the expression of RANK [111]. Additionally, scientists have developed a microneedle (MN) patch containing biodegradable GelMA MNs that release IL-4 and TGF-β in later stages to facilitate anti-inflammatory polarization towards M2 macrophages and Treg cells, accelerating tissue healing [112]. Moreover, since the osteoinductive and immunomodulatory properties of DNA were discovered [99], scientists have designed a DNA-modified collagen (DNA-Col) that enhances recruitment and differentiation of Treg cells through modulation of mitochondria-related metabolic processes, leading to improved BREG (Fig. 2C) [94]. Additionally, it has been discovered that inducing Tregs through immunomodulation on DCs towards a tolerogenic phenotype presents a promising approach for individuals with diabetes [113]. In conclusion, the modulation of Th2 and Treg via hydrogel to suppress inflammation and induce osteogenesis could potentially serve as a prospective approach in the treatment of IBD.
4.5
Regulating DCs in IBD
The involvement of DCs in immune responses and their ability to differentiate into osteoclasts or indirectly influence the process of BREG by regulating T cell subsets make the concept of modulating DCs for infection control and promotion of BREP highly promising. The compromised tolerogenic DCs and heightened IR have been observed to contribute to delayed healing of periodontitis, whereas tolerogenic DCs can stimulate the production of Tregs to suppress inflammation and mitigate alveolar bone loss [113]. Consequently, a chitosan-based thermosensitive injectable self-assembled hydrogel (TISH) was developed with the ability to inhibit differentiation towards mature DCs, promote polarization of DCs into a tolerogenic phenotype, and induce Treg formation for creating an anti-inflammatory microenvironment that facilitates improved BREP [113]. Scientists have discovered that the utilization of chitosan and hyaluronic acid-coated Mg2+ (CSHA-Mg) can effectively inhibit the activation of T cells by DCs. This inhibition is achieved through the induction of hypoxia inducible factor (HIF)-1α expression in DCs via transient receptor potential melastatin channel (TRPM)-7 channels and activation of the mitogen-activated protein kinase (MAPK) signaling pathways. Consequently, this approach leads to an elevation in Tregs levels, resulting in the creation of an immunosuppressive osteogenic microenvironment that facilitates BREG [114].
5.
Strategies for the delivery of EVs via hydrogels
EVs, derived from diverse cell sources, serve as potent mediators of intercellular communication by transferring bioactive molecules to regulate tissue repair and immune responses [115]. Recent advances highlight the potential of MSC-derived EVs in enhancing BREG, though their rapid clearance limits therapeutic efficacy [116]. To address this, hydrogels engineered for sustained EV delivery have emerged as innovative platforms to prolong EV retention and bioactivity. Studies demonstrate that hydrogel-encapsulated EVs can simultaneously combat infection, modulate inflammation, and promote osteogenesis (Fig. 2D) [100]. For instance, oxygen-releasing hydrogels loaded with MSC-EVs mitigate anaerobic infections while supporting BREP, and T-cell immunoglobulin mucin family member 3 (TIM3)-overexpressing EVs in GelMA hydrogels drive M2 macrophage polarization to resolve inflammation [117,118]. These strategies underscore the dual role of EV-hydrogel systems in balancing immune regulation and regenerative processes. The supplementary materials elaborate on these cutting-edge approaches, emphasizing their translational potential for addressing infection-impaired bone healing in IBD.
6.
Combination of hydrogels with advanced techniques
Emerging hydrogel-integrated technologies include injectable hydrogels, 3D-printed scaffolds, stimuli-responsive biomaterials, and programmable DNA hydrogels are revolutionizing therapeutic approaches for IME modulation in IBD. These innovations enable precise immunoregulation, spatiotemporal drug delivery, and personalized tissue engineering while addressing challenges such as irregular defect geometry and dynamic IRs. A comprehensive analysis of their translational potential, technical limitations, and clinical applicability in IBD management is systematically evaluated in the Supporting information.
7.
Conclusion and future perspectives
The intricate nature of the IME often renders biomaterials with direct bactericidal activity ineffective over time, thereby impeding the efficacy of traditional strategies [38]. Moreover, infection-induced dysfunction of the immune system can disrupt BREG and promote bone resorption, resulting in delayed BREP in cases of bone defects. Therefore, the immunomodulatory strategies that focus on targeting the IME of IBD hold promise for effectively controlling infections, suppressing excessive inflammation, and achieving more natural and functional outcomes for BREP with reduced complications or side effects. Compared to other drug delivery systems, hydrogels possess several key advantages including low invasiveness and injectability, controlled release of encapsulated substances, biodegradability with non-toxic degradation products, responsiveness to external stimuli, and programmed delivery ability [19]. This makes hydrogels widely used in regulating the IME for BREP [17,100,104]. This review discusses the effects of different ICs and cytokines in regulating anti-infection processes as well as inflammation regulation and induction or hindrance of BREP at different stages. Additionally, it explores strategies and current research progress related to directly regulating ICs and delivering EVs through hydrogels to regulate microenvironments and promote BREP for treating IBD. In summary, this review aims to provide a comprehensive overview along with new strategies for clearing infections and accelerating BREP in IBD through hydrogels. However, although the regulation of IME through hydrogels holds promise as a strategy for effectively clearing infections and establishing a pro-osteogenic IME for BREP, there are still several challenges that need to be addressed and overcome. Meanwhile, advancements in emerging technologies also stimulate novel perspectives.
The current study is primarily focused on investigating the role of macrophages in IBD, while there remains a dearth of research regarding the impact of other ICs, including neutrophils, NK cells, DCs, eosinophils and basophils, as well as their respective phenotypes. Furthermore, the effects exerted by these ICs during different phases of IBD remain largely unknown. There is a lack of knowledge regarding the systematic interaction between different ICs and cytokines in the IME, as well as the network effects composed of these ICs and cytokines with regards to BREP in IBD. This limitation results in most immunomodulatory hydrogels focusing only on single types of cells, neglecting their network impacts on the entire immune system and lacking a comprehensive theoretical foundation for their programmed delivering to exert diverse regulatory capacities at various stages of IBD. Consequently, this approach fails to optimize therapeutic efficacy or may encounter hindrances due to spontaneous regulation of the immune system in IBD. Most studies have primarily focused on investigating the effects of the IME on IBD caused by bacterial infections, while neglecting the impact of viral and fungal infections. This limitation hinders the further development and widespread utilization of immunomodulatory hydrogels. Meanwhile, the intricate interaction between the IME in IBD and the overall systemic microenvironment remains poorly elucidated [19]. The investigation into modulating the IME for individuals with specific health conditions, such as obesity, diabetes, vitamin deficiencies, and autoimmune diseases that may impact immune system functionality and inflammation levels still lacks clarity to some extent, which poses challenges in tailoring personalized hydrogel treatments [19].
The immunomodulatory strategies targeting the IME via hydrogels hold significant promise for clinical translation in IBD treatment. However, several challenges remain that must be addressed to facilitate their clinical application. First, safety and efficacy are paramount considerations for the clinical translation of hydrogels. Given that hydrogels consist of various components, the safety and effectiveness of these components require further verification through rigorous clinical trials. Additionally, accurate analysis of in vitro and in vivo degradation rates is essential to ensure that hydrogel formulations align with their intended applications. Ideally, hydrogels should initially provide mechanical support while gradually degrading alongside tissue regeneration, serving as a scaffold that promotes tissue regeneration without hindering cell migration, proliferation, or differentiation. Finally, the synthesis process of hydrogels should be simplified to reduce costs while maintaining safety and efficacy. One promising solution to these challenges is model-based modular hydrogel design, a framework that integrates theoretical modeling with modular synthesis strategies. This approach aims to optimize the multifunctional properties of hydrogels and accelerate their clinical translation in biomedical applications through a systematic methodology [119].
Hydrogels delivering molecules and bioactive substances, including BMP-2 which is currently recognized as the most effective osteoinductive drug [120], may result in unstable and unsustainable drug release efficiency through hydrogels. Therefore, engineered EVs, considered as an efficient medium for transmitting biological information, can be utilized to carry molecules such as proteins, micro-RNAs, and long non-coding RNA (lncRNAs) and assist in packaging drugs that cannot be directly loaded into hydrogels [118]. Furthermore, the current research primarily focuses on EVs derived from MSCs. Retaining the excellent bioactivity of parent cells and offering significant advantages beyond cell therapy [100], natural EVs derived from various cells exhibit distinct effects on the IME, making them a promising avenue for regulating the microenvironment in IBD when harnessed through hydrogels. Furthermore, 3D-printed scaffolds have been extensively utilized in bone defect research due to their personalized design capabilities, excellent biocompatibility, and ability to form porous structures that create optimal conditions for cellular implantation, thereby expediting the process of BREP [121,122]. However, it has been observed that 3D-printed scaffolds can serve as a microenvironment for bacterial colonization and induce an IR leading to dysregulation of inflammation [121,123]. Therefore, the integration of 3D printing technology and IME regulation strategies could potentially serve as a prospective approach for mitigating the drawbacks associated with 3D printing while harnessing its advantages in the treatment of IBD, as 3D-printed scaffolds can continuously release molecules to modulate the IME, and regulate the process of neovascularization and osteogenesis at different stages [124,125].
Four-dimensional (4D) printing technology is based on 3D printing, it integrates time as the fourth dimension, enabling printed constructs to dynamically transform their shape or functionality in response to external stimuli. 4D printing utilizes intelligent materials as printing ink to enable bioactive scaffolds to respond dynamically to various stimuli over time, including physical factors such as temperature, light, electricity and chemical factors such as pH, along with biological factors like enzymes [124,126]. In comparison to traditional 3D printed scaffolds, the unique ability of 4D-printed scaffolds to change their properties under different stimuli makes them more intelligent in terms of programmed delivery of hydrogels and 3D-printed hydrogels [124]. The key processes of 4D printing primarily involve the selection of materials and the conduct of external stimulations. The materials utilized in 4D printing must possess biocompatibility, encompassing not only biological inertness but also biostability and functional biological activity including the functional responsiveness under physiological conditions. Additionally, these materials should exhibit biodegradability, meaning they must be capable of breaking down under specific environmental conditions. Furthermore, printability is crucial as the fundamental structures of 4D printing materials are constructed using 3D printing technology. In addition to the aforementioned requirements, materials utilized in 4D printing should also possess properties such as shape memory, self-healing, and self-assembly. These characteristics are essential for achieving the complex deformation behaviors inherent to 4D printing and for forming ordered structures that can serve as fundamental components of shape memory materials [127]. In addition to material composition, external stimuli should also be considered in 4D printing. The materials used in 4D printing are capable of altering their structural configurations in response to a variety of physical and chemical stimuli. These stimuli include, but are not limited to, temperature variations, magnetic fields, electric fields, light exposure, acoustic waves, pH levels, and changes in ionic concentrations, achieved through the incorporation of responsive units [128]. Overall, the intelligent-response feature of 4D printings holds great promise for dynamically regulating IME suitable for infection clearance and creating osteoinductive microenvironments that adapt to changing stimuli over time. Five-dimensional (5D) printing introduces two additional rotational axes (total 5 axes) to the traditional 3D printing system, allowing curved-layer deposition and improved structural integrity [129]. The printers of 5D printing, unlike the x, y, and z axes of 3D printing, incorporate two additional axes to enhance structural integrity by enabling curved layers that are specifically designed for defect areas. This innovative approach enhances mechanical properties while optimizing material usage, making it particularly suitable for orthopedic applications because the human skeletal surface is not flat but curved. Meanwhile, the mechanical properties of implanted materials for load-bearing bones in IBD treatment have stringent requirements, which makes 5D printing a particularly promising technology that surpasses both 3D and 4D printing [124,130]. Six-dimensional (6D) printing represents an innovative concept that integrates the dynamic adaptability of 4D printing with the multi-axis capabilities of 5D printing, aiming to achieve real-time adaptation to intricate microenvironmental changes [131]. Beyond printing along multiple axes, 6D printing emphasizes leveraging established 4D and 5D printing techniques to develop biomaterials capable of intelligent responses to environmental stimuli. Utilizing five degrees of freedom, it produces smart objects that can alter their shape or properties in response to external stimuli, while retaining the design flexibility characteristic of 5D printing materials [129]. These attributes enable 6D printing materials to meet the complex shape requirements and stringent mechanical properties necessary for IBD treatment, intelligently responding to changes in the IBD environment. Consequently, this integration of 3D/4D/5D technologies yields superior treatment outcomes compared to individual printing methods. However, the widespread adoption of 6D printing has been impeded by manufacturing challenges, including the complexity of 6D printing equipment and its currently high cost, which future research should aim to address. Overall, as emerging technologies in printing, both 5D and 6D printing offer possibilities for enhancing the quality, programmability, and tunability of printed objects' multi-intelligent response to microenvironmental stimuli in IBD [124]. The integration of artificial intelligence (AI) with immunomodulatory strategies offers a promising tool for personalized patient care [132]. For instance, machine learning (ML) and deep learning (DL) techniques enable infection recognition, drug susceptibility analysis, and disease progression prediction in osteomyelitis [133]. Meanwhile, it can also be applied in personalized diagnosis and treatment. The utilization of ML and DL presents immense potential in exploring the intricate interactions among pathogenic microorganisms, the immune system, and the skeletal system.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
CRediT authorship contribution statement
Peizhang Zhao: Data curation, Conceptualization. Mengmeng Li: Data curation. Jingwen Wang: Data curation. Jun Li: Conceptualization. Yunfeng Lin: Writing – review & editing, Funding acquisition, Conceptualization.
Acknowledgments
This study was supported by the National Natural Science Fund of China (Nos. 82202726, 82370929); the National Clinical Research Center for Geriatrics, West China Hospital, Sichuan University (No. Z20192013); Key research and development project of Sichuan Science and Technology Department (No. 2023YFG0219); "ZerotoOne" Innovation Research Project of Sichuan University (No. 2022SCUH0014); Frontiers Medical Center, Tianfu Jincheng Laboratory Foundation (No. TFJC2023010001), Sichuan Science and Technology Program (No. 2022NSFSC0002), Sichuan Province Youth Science and Technology Innovation Team (No. 2022JDTD0021), Research and Develop Program, West China Hospital of Stomatology Sichuan University (Nos. RD03202302, RCDWJS2024–1), Health Commission of Sichuan Province Medical Science and Technology Program.
Supplementary materials
Supplementary material associated with this article can be found, in the online version, at doi:10.1016/j.cclet.2025.111319.
[1]
J. Wang, P. Zhao, M. Li, J. Li, Y. Lin, Chin. Chem. Lett. 36 (2025) 110686. doi: 10.1016/j.cclet.2024.110686
X. Han, Q. Saiding, X. Cai, et al., Nanomicro Lett. 15 (2023) 239.
[125]
Y. Yuan, Y. Xu, Y. Mao, et al., Adv. Sci. 11 (2024) e2306428.
[126]
L. Qin, S. Yang, C. Zhao, et al., Bone Res. 12 (2024) 28.
[127]
Z. Wang, D. Ma, J. Liu, et al., Bioact. Mater. 48 (2025) 370–399.
[128]
X. Chen, Z. Xu, Y. Gao, et al., Adv. Mater. 36 (2024) e2406118.
[129]
B. Makurat-Kasprolewicz, H. Ipakchi, P. Rajaee, et al., Chem. Eng. J. 494 (2024) 152703.
[130]
A. Sheikh, M.A.S. Abourehab, P. Kesharwani, Drug Discov. Today 28 (2023) 103391.
[131]
M. Li, P. Zhao, J. Wang, X. Zhang, J. Li, Mater. Horiz. 12 (2025) 20–36.
[132]
T.J. DuCote, K.J. Naughton, E.M. Skaggs, et al., Lab. Invest. 103 (2023) 100176.
[133]
Z. Shao, H. Gao, B. Wang, S. Zhang, Front. Cell. Infect. Microbiol. 14 (2024) 1380136.
Figure 1
(A) The NETs effectively eliminate bacterial infections and bacteria employ virulence factors to evade NETs release during infection. Reproduced with permission [24]. Copyright 2022, Frontiers. (B) The presence of zinc ion (Zn2+) enhances the generation of ROS-dependent NETs and facilitates osseointegration in rat femur infection. Reproduced with permission [32]. Copyright 2023, KeAi Communication. (C) The formation of NETs is crucial for activating pro-inflammatory macrophages. In contrast, inhibiting NET formation reduces this pro-inflammatory response, which in turn enhances bone formation around implanted biomaterials in non-infectious environments. Reproduced with permission [33]. Copyright 2023, Elsevier. (D) The neutrophils differentiate into different phenotypes in the different phases of BREP. Reproduced with permission [34]. Copyright 2024, Elsevier.
Figure 2
(A) A ’micro-cage’ composed of mPDA-PEI@GelMA that clears the NETs in the damaged area to suppress inflammation. Reproduced with permission [96]. Copyright 2024, Wiley. (B) The 3D-printed silk fibroin/gelatin basal scaffold, loaded with xonotlite nanofiber (Ca6(Si6O17) (OH)2, CS), was observed to effectively modulate the polarization of macrophages towards the M2 phenotype. Reproduced with permission [39]. Copyright 2024, Springer Nature. (C) The DNA-Col scaffold, crosslinked with deoxyribonucleic acid, induced the recruitment of Tregs to enhance bone defect healing. Reproduced with permission [94]. Copyright 2023, Elsevier. (D) The composite system facilitates the transportation of EVs to enhance bone tissue regeneration. Reproduced with permission [100]. Copyright 2024, American Chemical Society.

 DownLoad:
DownLoad:

 下载:
下载:



 下载:
下载:
