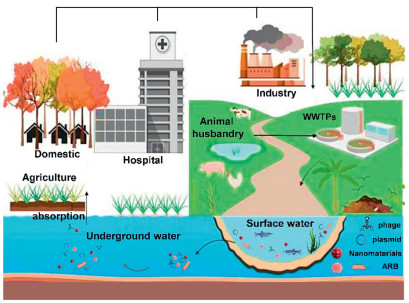
Figure 1.
ARGs in the aquatic environment.

Since the advent of antibiotics, their use has significantly improved the treatment and prevention of illnesses in both humans and animals [1]. However, the excessive and inappropriate use of antibiotics in veterinary medicine, animal husbandry, human health, and industrial practices has led to the rapid emergence and dissemination of antibiotic-resistant bacteria (ARB) and antibiotic-resistant genes (ARGs) into the environment (Text S1 in Supporting information), posing dire risks to human and animal health, and complicating the clinical disease management processes [2]. Currently, global consumption of antibiotics continues to rise annually, with an increase from approximately 21.1 tons in 2000 to 34.8 billion tons in 2015 [3]. According to the World Health Organization's (WHO's) Global Monitoring System for Antimicrobial Drugs, the estimated annual death toll from antibiotic-resistant bacterial infections stands at around 700,000 [4]. As an emerging category of contaminants, the transfer and proliferation of ARGs across various environmental media pose greater environmental challenges and health risks than antibiotics themselves [5]. For example, the presence of ARGs has significant implications to ecosystems, potentially exacerbating biodiversity loss in natural habitats. Over the past few years, ARGs have been identified in a wide range of aquatic environments, including surface water [6], municipal wastewater [7], and groundwater (Tables S1-S3 in Supporting information) [8]. In the 21st century, WHO has identified ARGs as one of the most serious threats to humanity [9].
In addition to the widespread dissemination of ARGs that results from the use and misuse of antibiotics, other non-antibiotic contaminants, including sterilization by-products, preservatives, and engineered nanomaterials (NMs), have also been implicated in the dissemination of ARGs [10–12]. The rapid advancement of nanotechnology has led to an increasing influence of nanoproducts on human life. The unique physicochemical properties (morphological features, surface properties, dimensional attributes) of NMs are harnessed to develop a diverse array of products that serve the societal, economic, and industrial needs [13]. For instance, nano-silver (Ag) is used as an antimicrobial agent in children's clothing, toys, and washing machines due to its antiseptic properties [14,15], while various metal oxide nanocomposites such as titanium dioxide (TiO2) and iron oxide (Fe2O3) are incorporated in cosmetics and sunscreens [16,17]. Engineered NMs are extensively used for protecting the environment, thanks to their advanced and specialized properties that facilitate the removal of pollutants (such as through adsorption and catalytic degradation) and exhibit antimicrobial activity [18,19]. For example, combinations of NMs and antibiotics have been used to combat multidrug-resistant bacteria [20]. However, these engineered NMs may inadvertently or deliberately be released into aquatic environments during their manufacture, application, or disposal. Conventional engineered NMs, including TiO2, Ag-NMs, and carbon nanotubes (CNTs), have been detected in various environmental media, prompting increased research interest in the adverse effects of NMs on the ecosystem [21,22].
Given the utilization of NMs and the mechanisms driving the rapid evolution of ARGs, concerns regarding their role in antibiotic resistance, particularly in the horizontal transfer of ARGs, have escalated [23,24]. Numerous studies have shown that NMs can exert varying effects on the elimination and horizontal transmission of ARGs [25], with some suggesting a retarding or inhibitory role of NMs in the horizontal transfer of ARGs [26,27], while others indicating a facilitating role of NMs in the horizontal transfer of ARGs [28–30]. This highlights the complex and diverse impact of NMs on microorganisms and their genetic material. In summary, while the development of nanotechnology has introduced numerous innovative products and applications, it has also presented a range of challenges and concerns to the safety of ecosystem. Therefore, it is necessary to study the interaction mechanism between NMs and ARGs transfer to effectively control the spread of ARGs.
In the present study, a co-occurrence network analysis is conducted using VOS Viewer to visualize the relationship between NMs and resistance genes. The analysis reveals a strong connection between these elements, as evidenced by the high-frequency words identified (Fig. S1 and Text S2 in Supporting information). The study provides a comprehensive overview of the mechanisms of the transmission of antibiotics and the influence of NMs on the propagation of ARGs. The specific pathways through which the chemical and physical characteristics of NMs affect the spread of ARGs are discussed. Moreover, the incidence, behavior, and potential hazards associated with resistance genes are also summarized. Furthermore, the NM-mediated removal of ARGs and the mechanisms underlying the horizontal gene transfer (HGT) are critically evaluated. The balance between HGT and the removal of resistance genes by NMs is highlighted, elucidating the differences in the transmission of ARGs attributed to various compositions of NMs. The results of this study are crucial for enhancing the understanding of how NMs interact with microorganisms. The research on the mechanism of how NMs affect the propagation of ARGs often focuses on metal NMs. This article comprehensively analyzes the mechanism of how NMs affect the transfer of ARGs by combining metal, non-metal, and composite NMs. Overall, the review aims at providing guidance on future research directions, giving insights into the role of NMs in the field of transmission of ARGs, and offering theoretical support for the development of safer and more effective applications of NMs.
The entry of antibiotics and other pollutants, their metabolites, and various mixtures of ARB into aquatic ecosystems occurs through both treated and untreated sewage, discharges from animal husbandry, aquaculture, and surface runoffs [31]. The persistent presence of antibiotic residues in aquatic environments poses significant threat to the health of humans and animals alike, inhibits the growth of certain aquatic organisms, disrupts the balance of microbial communities, and diminishes the self-purification capacity of water bodies. With this backdrop, HGT can be facilitated by the selective pressure exerted by antibiotics, leading to the accumulation of ARGs in aquatic environment. Wastewater treatment plants (WWTPs) serve as primary reservoirs for ARB and ARGs, significantly influencing the proliferation of ARGs among bacteria through HGT and their subsequent dissemination throughout aquatic ecosystems (Fig. 1) [32,33]. The ARGs detected in the influent of WWTPs predominantly include macrolides (ermB, ermF), tetracyclines (tetA, tetC, and tetW), sulphonamides (sul1, sul2), and β-lactams (blaOXA, blaTEM) [34]. Table 1 presents an overview of the detection of ARGs and ARB across various aquatic environments [35–42].
 DownLoad:
CSV
DownLoad:
CSV
| Environment | ARGs | MGEs | Remarks | Ref. |
| WWTP effluent | tetA, tetC, tetG, tetX, tetM, tetO, tetW, sul1, sul2 | intI2, intI1 | – | [35] |
| South China Sea | macB, acrB | – | Pristine environment with minimal human activity | [36] |
| Yangtze River | rosA, rosB, macB, bacA, acrB, adeB, cpxR, amrB, ceoB, cmeB, mdtB, mdtC, mdtF, tetP, mexB, mexC, mexD, mexE, oprC, mexF, mexI, mexW, qepA, vanR | – | Freshwater ecosystems | [37] |
| Livestock breeding wastewater | ant(6′)-Ia, tetX, aac(6′)-Ie, tetM, aph(3′)-IIIa, tetW, tetQ, ant(3″)-Ia, tetO | – | Pig farming wastewater | [38] |
| Mariculture system | tetB, tetG, tetX, qnrS, sul1, sul2, ermF, ermT, qnrA, qnrB, | intI1 | – | [39] |
| Karst River | aadA-01, sul2, aadA-02, blaOXA-21, aadA1, tetG, emrA, aadA2–03, qacEdelta1–01, mdtK, ereA, qacEdelta1–02, | tnpA-04, tnpA-01, tnpA-02, tnpA-03, intI1, cintI1, Tp614, tnpA-05, tnpA-07, IS613 | Drinking water source | [40] |
| Municipal solid waste leachates | sul1, sul2, tetM, tetQ, ermB, mefA | – | – | [41] |
| North Yellow Sea of China | sul1, sul2, tetM, tetX, tetB, blaTEM, qnrS, qnrA, qnrB, qnrF, tetG, ermF, ermT | intI1 | Coastal areas with dense human activities | [42] |
Despite the implementation of multiple techniques aimed at mitigating the environmental spread of ARGs through adsorption and degradation, the efficiency for the removal of ARGs remains suboptimal and, in certain cases, may inadvertently promote the spread of ARGs [43]. Previous research indicates that the concentration of ARGs in wastewater and sludge can be 1.0–14.1 times greater than that found in the influent water, suggesting that ARGs proliferate during their treatment phase [44,45]. Wastewater discharges from WWTPs are recognized as a significant source of antibiotics and ARGs in river systems [46]. Previous research has indicated that, compared to the aquatic bodies that remain relatively undisturbed, ARGs are spreading at much higher rates in river bodies, which is due to the anthropogenic effects [47]. For instance, 27 ARGs have been identified in raw water from the Pearl River Delta, a socioeconomically developed region in China [48], while a study detected 258 ARGs, predominantly aminoglycosides and bla genes, in various Chinese urban rivers [49]. Moreover, ARB and ARGs entering the surface water through infiltration, rainfall, and other factors can have additional effects on soils and groundwater.
Drinking water primarily originates from river systems and groundwater [50]. Therefore, the majority of these ARGs are introduced into the human body through drinking water. This water is essential for human survival, and the presence of ARGs in drinking water constitutes a crucial pathway for the transfer of ARGs and pathogens from the environment to humans (Fig. S2 in Supporting information) [51,52]. Consequently, the cyclic transmission of ARGs in the environment suggests that these genes will ultimately accumulate in human bodies. In addition to the discharge of antibiotics and ARGs from WWTPs, aquaculture represents another major source of antibiotic and ARG contamination in water bodies [53]. Wang et al. identified 21 types of ARGs in samples from industrial mariculture systems, with tetracycline, chloramphenicol, streptomycin, sulfonamides, and fluoroquinolones exhibiting a high detection frequency of 90%−100% [54]. These residual ARBs and ARGs can subsequently spread and proliferate in the natural aquatic environment.
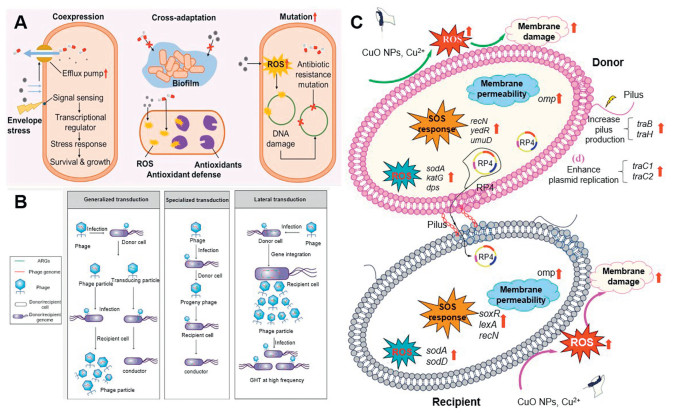
Antibiotic resistance is facilitated through the acquisition of resistance genes between bacterial species through vertical gene transfer (VGT) and HGT. Among them, HGT is the predominant mechanism for the transmission of ARGs [55]. Moreover, HGT among bacteria can be categorized into three main types: (1) transformation; a process in which competent recipient bacteria uptake extracellular DNA (eDNA), thereby acquiring resistance (Fig. 2A); (2) transduction; the transfer of ARGs from a donor to a recipient bacterium through phages (Fig. 2B); (3) conjugative; involves the transmission of various mobile genetic elements (MGEs) such as transposons, integrons, and plasmids through direct cell-to-cell contact (Fig. 2C). Conjugation facilitates the transfer of ARGs among various microorganisms through direct interactions, enabling MGEs to migrate among various microorganisms [56].
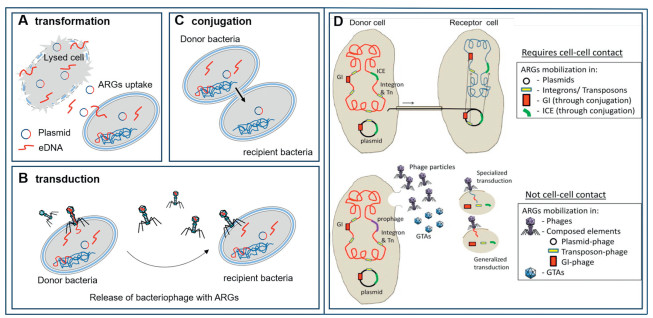
With regards to the three types of HGT, the conjugative transfer is regarded as the primary mechanism for the dissemination of ARGs, as it occurs in a wide range of transmissible resistance plasmids, offering superior resistance to environmental pressures and a more efficient means for penetrating host cells. Compared to phage transduction, this mechanism (conjugative transfer) typically facilitates a broader range of potential hosts [58]. In addition, MGEs mediate the transfer of ARGs between recipient bacteria and donors under conditions that promote cellular contact and fusion (Fig. 2D) [57]. Following the successful acquisition of a plasmid, these ARGs can be transmitted efficiently from donor to recipient bacteria through interactions involving cell surface hyphae or adherent intercellular contacts [26]. The process of MGE-driven HGT is closely linked to variations in the abundance of ARGs. Yang et al. demonstrated that MGEs significantly contribute to HGT and that there is a strong positive correlation between the absolute abundance of ARGs and MGEs [59].
Lucas et al. reported that ARGs present in French porcine Escherichia coli predominantly consisted of blaCTX-M-1 and IncI1/ST3 plasmids, which harbored numerous ARGs over the past decade and facilitated the horizontal transfer in the environment through the recombination of plasmids [60]. Numerous studies on the dissemination of antibiotic resistance plasmids in human infections indicated that once resistance genes were integrated into functional plasmids, they could rapidly spread across multiple strains, species, and even genera [61]. Globally, plasmid pCT has been identified as a vector for the transmission of blaCTX-M-14 between animals and humans across continents, with these genes being transferred to a variety of plasmids with a diverse range of hosts within the E. coli family and other human pathogens [62]. Three variants of transposons containing the same blaKPC-3 carbapenemase gene were successfully transmitted between different strains of E. coli and Klebsiella pneumoniae, as well as different plasmid incompatible groups [63]. This underscores the significant role of MGEs in facilitating gene transfer across a broad range of hosts.
Phage-mediated transduction of ARGs is also considered crucial in HGT, as bacteriophages play a key role in the transduction of ARGs. Present in various environments, phages can extract ARGs from infected bacteria, enabling the transfer of these genes to other bacteria through transduction, thus facilitating HGT of resistance genes to distinct bacteria [63,64]. Unlike transformation and coupling, transduction is strictly host-specific, necessitating recognition between the specific phage proteins and the host cell [65]. The movement and transfer of ARGs through phages have been documented across various bacterial species. For instance, Rands et al. demonstrated that the Acidaminococcus phage served as a vector for the β-lactam resistance gene aci1 [66]. Ji et al. reported that phages in the porcine cecum acted as the primary reservoir for MGEs and ARGs [67]. The process whereby bacteria directly absorb free extracellular ARGs is known as transformation. During the transformation process, bacteria can retract their hyphae to draw double-stranded DNA closer to their cell membranes, facilitating uptake through conserved membrane pores during natural transformation. However, transformation is generally less efficient than conjugation, which is due to the reason that extracellular DNA containing ARGs is prone to degradation in the surrounding environment [68].
Numerous studies have highlighted a significant association between the structure of bacterial communities and ARGs. For example, Han et al. found that the increase in ARGs' abundance resulting from the addition of fishmeal was primarily attributed to shifts in bacterial communities, with these community variations contributing more significantly to changes in antibiotic resistance patterns than other environmental factors or HGT mediated by MGEs [69]. Research investigating the link between microbial communities and ARGs in sludge obtained from municipal WWTPs has shown that the proliferation and distribution of specific ARGs are closely linked to certain microbial communities [70]. Chen et al. reported that removing microorganisms from manure significantly diminished the spread of ARGs and that a highly localized microbial community restricted the transport and dissemination of ARGs from manure to soil [71]. Moreover, previous studies have shown that, compared to simpler communities, microbial communities with considerable diversity are generally less susceptible to invasion by foreign microbes. Wu et al. discovered that bacterial communities predominantly drive changes in ARGs during aerobic composting, whereas the observed reduction in the abundance of ARGs can be attributed to the suppression of bacteria that potentially harbor ARGs during the composting process [72]. Overall, these findings suggest that environmental factors capable of influencing the structures of bacterial communities may also play a crucial role in the dissemination of ARGs.
Nanomaterials (NMs) impact the structures of bacterial communities. Additionally, both metallic NMs and their secreted metal ions can promote HGT (Text S3 in Supporting information). Qiu et al. discovered that NMs in aquatic environments significantly enhanced the HGT of multiple drug ARGs mediated by RP4, RK2, and pCF10 plasmids [73]. More specifically, nano-Al2O3 in water increased the frequency of RP4 plasmid transfer from E. coli to Salmonella by 200-folds, enhanced the Gram-negative to Gram-positive transfer by 50-folds, and increased the intraspecific transfer within the genus E. coli by 100-folds. In addition, Ding et al. found that nano-Al2O3, TiO2, and Fe3O4 significantly enhanced the plasmid-mediated transformation of ARGs into Gram-negative E. coli strains and Gram-positive Staphylococcus aureus, with nano-Al2O3 demonstrating the highest efficiency to this end [74]. Lu et al. showed for the first time that nano-Ag and Ag+ could facilitate the increased HGT of ARGs (Fig. S3 in Supporting information), as evidenced by the frequency of HGT for the transfer of PR4 plasmid from E. coli to Pseudomonas malodorata that was 1.8 times greater than that of controls in the presence of nano-Ag [75]. Markowicz et al. found that 10 mg/L of ZnO nanoparticles enhanced the transformation frequency of E. coli by nearly 1.8 times (Fig. 3A) [76]. Chen et al. demonstrated that the incorporation of nano-ZnO led to a significant increase in the abundance of MGEs (intI1), which showed a strong positive correlation with various ARGs, indicating that ARGs propagated through HGT following the addition of nano-ZnO [77]. Zhang et al. reported that environmentally relevant sub-inhibitory concentrations of nano-CuO and Cu2+ could stimulate the conjugative transfer of ARGs [78].
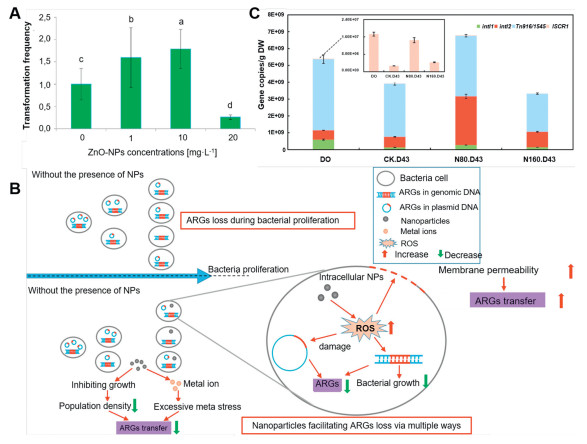
Furthermore, beyond enhancing the conjugative transfer of ARGs, certain NMs have been shown to contribute to the reduction of ARGs [79]. For instance, Su et al. reported that NMs synthesized from metals and metal oxides were effective in decreasing the diversity of ARGs, achieving reductions in the abundances that lied within the range of 1.4–3.2 orders of magnitude (Fig. 3B) [80]. Moreover, Fe2O3@MoS2 nanocomposites can inhibit the HGT of RP4–7 plasmid by suppressing the transcription of conjugative transfer genes [81]. Nano zero-valent iron (nZVI) has been shown to facilitate antibiotic biodegradation through adsorption and reduction while also impacting microbial communities [82]. Furthermore, Ma et al. indicated that the addition of 160 mg/L nZVI resulted in a reduction of integrons (intI1 and intI2), thereby lowering the risk of HGT of ARGs during the anaerobic digestion of cow dung (Fig. 3C) [83]. Research has shown that graphene nano zinc oxide (nano-GZnO) disrupts bacterial chemotaxis and minimizes ARG transfer by altering microbial community structure [84]. Nano-Ag control microbial populations and prevent ARG from spreading in anaerobic ammonia oxidation flocs by blocking the nitrogen cycle driven by anaerobic ammonia oxidation [85]. In addition, NMs can foster phage transduction while facilitating the conjugative transfer of ARGs (Text S4 in Supporting information). For instance, nano-TiO2 promotes the attachment of phages to the surface of E. coli TG1 cells, thereby promoting the transduction of ARGs through phage infections (Fig. 4A) [86]. In such a case, the transfer of ARGs that were mediated by the phage (gM13) in the presence of 20 nm TiO2 at a concentration of 0.5 mmol/L was found to be 4.5 times higher than that in the control group. This phenomenon was attributed to the presence of nano-TiO2 particles on the surface of cells, which provided a substantial specific surface area that facilitated the aggregation of phages around the cells and enhanced the process of phage transduction.
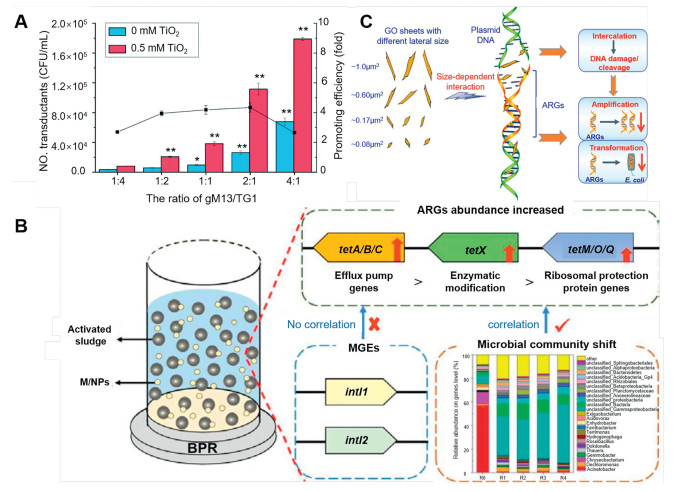
In addition to metal-based NMs, various synthetic non-metallic NMs such as graphene and microplastics can also facilitate the spread of ARGs [87]. For example, novel nanocomposites synthesized from Ag, TiO2, and graphene oxide significantly reduced the tetracycline-resistant bacteria (E. coli HB101) while simultaneously increasing the frequency of HGT within the genus E. coli by a factor of 1–2 [88]. Shi et al. showed that microplastics with the size range of 50-100 nm and NMs with the size range of 200-500 nm could promote conjugative transfer [89]. Zhou et al. reported that nanoplastics facilitated the transmission of MGEs (such as intl1 and intl2) along with multi-drug resistance genes (Fig. 4B) [90]. On the other hand, non-metallic NMs can also inhibit the spread of ARGs. For example, Xu et al. found that graphene oxide (GO) nanosheets, with modest lateral dimensions of 0.08–1.0 µm2, could effectively penetrate DNA molecules and cause significant cleavage of DNA [91], thereby obstructing the amplification and transformation of ARGs (Fig. 4C). However, elevated concentrations of GO can lead to the inactivation of ARB, thereby diminishing the frequency of conjugative transfer of ARGs [92].
In summary, numerous studies have indicated that NMs may promote HGT (Table 2), while reports on the inhibition of HGT by NMs are scarce in literature. The discrepancies in the impact of NMs on the propagation of ARGs may be linked to both the type and the concentration of NMs [12,26,75,78,81,86,88,93–96]. Typically, low concentrations of NMs can induce the production of reactive oxygen species (ROSs), thereby altering the permeability of cell membranes and facilitating the HGT of ARGs. In contrast, high concentrations of NMs may disrupt cellular activity, thereby inhibiting the HGT. Furthermore, the size of NMs is a critical factor influencing the propagation of ARGs. Bigger NMs do not significantly affect HGT, whereas smaller NMs induce oxidative stress and create nanopores on cell membranes, thus promoting the propagation of ARGs [91,97,98].
DownLoad:
CSV
| NMs | Physicochemical properties | HGT pathways | ARGs | Outcomes | Assessment endpoints | Ref. |
| Al2O3 | Metal oxide NMs | Conjugation | PBR322-based plasmid | Nano-Al2O3 improved the transmission of extracellular ARGs into bacteria through transformation. | Plasmid uptake, generation of ROSs, and permeability of cell membrane. | [94] |
| ZnO | Metal oxide NMs | Conjugation | Plasmid (pGEX4T-1) | The rate of transfer of RP4 plasmid increased in both the pure culture of E. coli and the mixed culture of native aquatic microbiota on exposure to zinc oxide nanoparticles. More precisely, compared to the non-ZnO nanoparticles-treated group, the transfer frequency of RP4 plasmid increased 24.3-folds in E. coli and 8.3-folds in aquatic microbiota on exposure to nano-ZnO at a concentration of 10 mg/L for 12 h. | Nano-ZnO facilitated the efficiency of transformation by naked plasmid pGEX4T-1 and improved the permeability of cell membrane. | [93] |
| TiO2 | Metal oxide NMs | Transduction | Bacteriophage (hM13) | Simultaneous exposure to humic acid (HA) and nanoscale titanium dioxide (nTiO2) synergistically promoted transduction under ultraviolet (UV)-A irradiation. The large amount of superoxide radicals generated by the photoexcitation of HA-nTiO2 combination increased the permeability of membrane and exacerbated the intracellular oxidative stress, thus accelerating the dispersion of ARGs through phage transduction. | Increased permeability of membrane, and generation of ROSs, induced intracellular oxidative stress. | [95] |
| CuO | Metal oxide NMs | Conjugation | Plasmid (RP4) | Under nano-CuO pressure (1–10 µmol/L), the conjugative transfer frequency of RP4 plasmid increased dramatically. When 5 µmol/L of nano-CuO was administered, the conjugative transfer frequency was more than twice that in the control groups. Conversely speaking, for the nano-CuO dosages of higher than 100 µmol/L, the rates of conjugative transfer of ARGs drastically dropped. | Overproduction of ROSs, permeability of membrane, RP4 gene expression, induced intracellular oxidative stress, ATP synthesis. | [78] |
| Ag | Metal NMs | Conjugation | Plasmid (RP4) | Conjugative transfer frequency of RP4 plasmid increased considerably on exposure to nano-Ag at the concentrations of 0.1 and 100 µg/L. Nevertheless, when the concentration of nano-Ag was increased to 1000 µg/L, the frequency of binding transfer of RP4 plasmids decreased significantly. | Overproduction of ROSs, permeability of membrane, RP4 gene expression, ATP synthesis. | [75] |
| Fe2O3@MoS2 | Nanocomposite materials | Conjugation | Plasmid (RP4–7) | Fe2O3@MoS2 nanocomposites significantly inhibited the conjugative transfer of RP4–7 by up to 100-folds. When the concentration of Fe2O3@MoS2 was lower than 100 mg/L, the inhibition of CT increased with the increase of the concentration of Fe2O3@MoS2 from 0 to 100 mg/L. | Suppression of plasmid horizontal transfer by inhibiting the expression of related genes (trbA, trbB, traF, and trfA). | [81] |
| Nanoplastics | Non-metallic NMs | Conjugation | Plasmid (RP4) | The 100 nm-sized Polystyrene (PS) has limited promotion of microorganisms due to its inhibitory effect on microbes. | PS particles altered the permeability of cell membrane and the activity of cell. | [86] |
| CeO2 | Metal oxide NMs | Conjugation | Plasmid (RP4) | In relatively high concentrations of nano-CeO2, the highly facilitated conjugation process contributed to the enhanced transfer of ARGs. Low concentrations of nano-CeO2 inhibited the conjugation process. At relatively high concentrations of nano-CeO2 (25 and 50 mg/L), the conjugative transfer of RP4 plasmids was enhanced by 118%–123%. In contrast, at low concentrations (1 and 5 mg/L), nano-CeO2 hindered the transfer by 22%–26%. | Production of ROSs, expression of plasmid-encoded genes. | [26] |
| TiO2/Ag/GO | Nanocomposite materials | Conjugation | ARB | 100 mg/L of TiO2/Ag/GO achieved 87.8%, 40.9%, and 56.4% inactivation on three bacteria (E. coli HB10663 and E. coli HB10667 and E. coli HB101) | The generation of ROSs induced intracellular oxidative stress. direct touch that oxidized the cell membrane and walls, and altered antioxidant enzymes. | [88] |
| MoS2 | Nanocomposite materials | Conjugation | Plasmid (RP4) | MoS2 significantly promoted the plasmid-mediated horizontal transfer of multiple ARGs. | Production of ROSs, altered ATP synthesis, and expression of related genes. | [96] |
| ZnS | Natural sphalerite | Conjugation | – | Natural sphalerite in lower concentrations (0.05–50 mg/L) facilitated the conjugation of ARGs, however higher concentrations (≥ 100 mg/L) prevented it. | Generation of ROSs, induced intracellular oxidative stress, membrane permeability, and conjugation-regulated gene expression. | [12] |
Given the variability in the types, morphologies, and sizes of NMs, different types of NMs exert varying degrees of regulatory influence on HGT of ARGs. Currently, the majority of studies have confirmed that NMs can facilitate the transmission of ARGs (Table S4 in Supporting information). Overall, the mechanisms by which NMs impact the transfer of ARGs encompass three primary components: (1) physical contact mediated by NMs; (2) metal ions released from metal-based NMs; and (3) ROSs generated by NMs. Section 5 will focus on (1) the physical properties of NMs and their effects on the physical state of bacteria, and (2) the regulation of the effect of ROSs produced by NMs on the physiological metabolism of bacteria.
Nanomaterials (NMs) exhibit a variety of physical characteristics such as their morphologies and sizes. Due to their unique adsorption capabilities, NMs are frequently employed as vectors for various contaminants, such as heavy metals and antibiotics, which can indirectly facilitate the HGT of ARGs [99]. Nanocomposites of TiO2/Ag/GO have been shown to enhance the frequency of gene transfer within E. coli spp. by 1–2 times [88]. Moreover, metallic NMs can accelerate the transfer of ARGs by releasing metal ions and inducing the production of ROSs. Previous research has indicated that specific physical properties of metallic NMs contribute to microbial resistance, thereby promoting bacterial resistance to other microbes (Fig. 5) [100]. In addition, the size of NMs plays a crucial role in the transfer of ARGs. For instance, nano-Al2O3 has been shown to facilitate conjugative linkages between Salmonella spp. and E. coli more effectively than bulk Al2O3 materials, thereby shortening the distance between the recipient and the donor cells and enhancing the exchange of genetic material [101]. Additionally, spherical NMs are more readily internalized through cell membrane, while sharp-edged NMs can penetrate the membrane by cutting through it.
NMs can facilitate the entry of nucleic acid fragments or drugs into the cells or tissues of plants and animals, as the cell membrane serves as a selective and partially permeable barrier that is vital in preventing the HGT of ARGs among various microorganisms. Moreover, NMs can adsorb onto the extracellular surface of bacteria or penetrate the cell membrane, thus increasing its permeability (Fig. S4A in Supporting information). This increase in membrane permeability enables the uptake of environmental DNA encoding ARGs by recipient bacteria, thus effectively accelerating the transformation and coupling of ARGs. Guo et al. demonstrated that GO could adsorb onto the surface of bacteria, thereby reducing the distance between donors and acceptors or altering the permeability of bacterial membrane, which increased the likelihood of transmembrane transfer of ARGs [102].
Furthermore, Kong et al. reported that 50 and 100 nm SiO2 increased the permeability of cell membrane by 12.5% and 9%, respectively, while 200 nm SiO2 showed no effect on the permeability (Fig. 6A) [103]. Nano-CeO2 enhances the permeability of E. coli cell membranes while decreasing the fluidity of membrane (Figs. 6B and C) [104]. Zhang et al. evaluated the permeability of cell membrane and discovered that both nano-CuO and Cu2+ promoted the conjugative transfer of ARGs [78]. Their findings indicated that exposure to nano-CuO or Cu2+ led to the upregulation of genes associated with the permeability of cell membrane in donor bacteria. Additionally, the proteins of OmpA, OmpC, and OmpF, which play crucial roles in regulating the permeability of membrane, exhibited increases of 2.1, 3.2, and 2.5 times in the level of gene expression, respectively. In addition, metal ions released by metallic NMs may interact with membrane proteins or enzymes containing amine, carboxyl, and sulfhydryl groups, subsequently leading to structural deformation of cell membrane [105,106]. For example, Ag+ emitted from Ag NMs can enhance the permeability of cell membrane and facilitate the transfer of extracellular ARGs to recipient cells [75].
NMs possess the capability to disrupt cell membranes and even cause nanopores within these membranes (Fig. S4B in Supporting information), thereby increasing the permeability of membranes and promoting the direct uptake of plasmids [74,107]. Compared to their bulk counterparts, the unique properties of NMs stem from their larger specific surface area, which is influenced by their physical characteristics including specific morphology and zeta potential [108]. Nano-ZnO with sharp edges has shown to induce the formation of nanopores on the surfaces of bacterial cells, resulting in a marked increase in the transformation efficiency of the naked plasmid of pGEX4T-1 [93].
Nno-Al2O3 (30 nm, 10 mg/L) significantly facilitated the transformation of ARGs from E. coli (E. coli ET12567) to Streptomyces coelicolor (M145), achieving a binding transfer rate of 182 cfu/108 cells, which was 60 times higher than that of the control [94]. The underlying mechanism was primarily attributed to the increased intracellular ROS levels that were induced by nano-Al2O3, which stimulated the expression of pore protein gene (ompC) and the binding-related gene (intA), thereby intensifying the diffusion of ARGs in the environment. This process allowed the transfer of ARGs, particularly the plasmid-based ones, from donor bacteria to recipient bacteria through the formed pores. Additional studies have indicated that nano-Al2O3 can also facilitate the transformation of ARGs into E. coli strain HB101 and Staphylococcus aureus IHEM 1.0301 by creating pores in the cell membrane [74]. Furthermore, nano-Ag increases the permeability of bacteria by promoting the aggregation of protein in the periplasmic space and the formation of nano-sized pores in the bacterial membrane [109].
Exposure to metal-based NMs leads to a rapid accumulation of mutations (Fig. S4C in Supporting information), which in turn fosters the emergence of new antimicrobial resistance mechanisms (Fig. 7A) [23]. For example, the exposure of nano-ZnO can induce alterations in the gyrB, mazG (involved in translation and transcription), and acrR (associated with the structure and translocation of membranes) genes, resulting in genetic modifications. Notably, changes in the gyrB gene have been shown to significantly contribute to the proliferation of ARGs [25]. Zhang et al. investigated the opportunistic pathogen E. coli K12's response to two types of NMs (nano-Al2O3 and nano-ZnO) and examined the hypothesis that NMs could induce antimicrobial resistance [25]. Nano-Al2O3 and nano-ZnO have been identified as significant enhancers of mutation frequency, thereby accelerating the development of antimicrobial resistance. Notably, 15 mutations across 7 genes were observed in 8 drug-resistant E. coli strains that were induced by Al2O3NMs. Among them, the expression of soxR protein was attributed to oxidative constitutive expression, which transiently triggered the transcription of soxS gene, subsequently facilitating the activation of certain regulatory ARGs by SoxS protein [110]. In the case of nano-ZnO, 16 mutations were detected across eight genes in eight drug-resistant E. coli strains, with alterations in the gyrB gene correlated with ARGs. Additionally, copper hydroxide NMs induce bacterial resistance to chloramphenicol and tetracycline through mutations in genes associated with E. coli K12 [111].
Phages act as carriers that mediate the HGT of ARGs, thus significantly influencing their dissemination (Fig. 7B) [64]. NMs possess the capability to adsorb eDNA, thereby potentially functioning in a phage-like capacity within cells (Fig. S4D in Supporting information). Certain studies have suggested that nano-Ag can serve as a vehicle for the delivery of genes [112]. Previous studies have shown that NMs such as pHEMA+ chitosan nanospheres (PCNS) and carbon nanomaterials can serve as carriers for gene delivery in vitro and in vivo [113,114]. Moreover, Pudlarz et al. demonstrated that various NMs including Fe2O3, carbon nanotubes, and gold can facilitate the transport of proteins, peptides, and other molecules into cells through both active and passive transport mechanisms [115]. Chang et al. found that mesoporous silica-centered NMs were effective carriers for the delivery of gene, particularly for RNA [116]. Zhao et al. showed that the modified magnetic nanomaterial of Fe3O4-PEI could transport exogenous DNA (including the plasmid of pEGFP-N1) across the cell membrane and into the cell [117].
Antibiotics are known to promote HGT by increasing the oxidative stress and SOS responses (Text S5 in Supporting information) [118]. When the levels of ROSs exceed the bacterial antioxidant capacity, the resultant damage to cell membrane occurs through interactions with various macromolecules such as lipids and proteins, thereby compromising the membrane barrier and inducing SOS responses that facilitate the transfer of ARGs [30,118,119]. Overall, an increase in the generation of ROSs correlates with a heightened efficiency of gene transfer.
Beyond their physical effects on bacteria, NMs can also induce cells to secrete ROSs, resulting in further chemical impacts on bacterial cells. Notably, significant accumulation of intracellular ROSs can lead to oxidative stress, promoting coupling effects [120]. The free hydroxyl radicals (•OH) produced during these interactions possess strong oxidizing potentials, posing risks to various micropollutants, bacterial cells, and potential damage to DNA [121]. For example, by testing and comparing the specific impacts of ROSs on MS-2 phage and E. coli, Cho et al. discovered that free •OH in solution could lead to the inactivation of MS-2 phage, while both free and surface-bound HO• could inactivate E. coli, which were additionally affected by other ROSs such as H2O2 and O2•- through secondary pathways [122]. Moreover, Sicwetsha et al. demonstrated that copper oxide NMs could catalyze the conversion of H2O2 into various ROSs (including HOO•-, •OH, and O2•-) [123]. Microplastics also can generate ROS. Microplastics' high specific surface area increased their capacity to come into touch with microbial cells. ROS are created when free radicals interact with molecules of dioxygen on their surface [124]. The oxidative stress of positively charged nanoplastics (50 nm modified by amine) is higher than that of negatively charged nanoplastics (55 nm unmodified) [125].
Beside the role of NMs in stimulating the secretion of ROSs from cells, numerous studies have underscored how the release of both NMs and ions can enhance the abundance of ARGs. It has been shown that metal ions such as Fe2+ and Cu2+ induce the production of •OH [126]. More specifically, Lu et al. found that nano-Ag and Ag+ could influence the activities of antioxidant enzymes such as catalase and alkyl hydroperoxide reductase, thereby promoting the conjugative transfer of ARGs [75]. Zhang et al. reported that CuO NMs enhanced the conjugative transfer of ARGs by increasing the formation of ROSs and initiating SOS responses [78]. Furthermore, exposure to nano-CuO and Cu2+ significantly upregulated the expression of oxidative stress-responsive genes (trxC, sodB, dps, sodA, katG, and katE) in donor E. coli strains, as well as oxidative stress-responsive genes (soxD, aphC, sodA, and aphF) in recipient Pseudomonas malodorans strains (Fig. 7C).
NMs represent a classic double-edged sword in the context of controlling the antibiotic resistance and the environmental transfer [127]. In addition, NMs can efficiently degrade antibiotics through adsorption and catalytic mechanisms (Fig. 8A) [128,129]. Some previous studies have shown that specific NMs can inhibit the HGT of ARGs by reducing the abundance of MGEs, suggesting their potential to limit the dissemination of ARGs. However, the prevailing view among researchers is that NMs facilitate the HGT of ARGs. Evidence suggests that while some NMs may suppress the gene expression of MGEs, they do not significantly alter the abundance of ARGs, thus allowing for their continued propagation in the environment.
In addition, the promotion of HGT of ARGs by NMs is influenced by the physical properties of NMs [130]. Variations in the size, material, and even shape of NMs distinctly affect the dynamics of HGT. Overall, NMs modulate the HGT of ARGs by altering the permeability of cell membrane, forming nanopores on the surface of membrane, promoting mutagenesis, acting as carriers, and facilitating the secretion of ROSs. These NMs can penetrate cells and induce the production of ROSs, which generally impacts the transfer of ARGs in a different way. Low levels of ROSs can enhance the permeability of cell membranes, thus promoting the HGT of ARGs. While excessive production of ROSs can further increase the permeability of membranes, damaging the ARGs encoded in both the genome and plasmid and ultimately inhibiting HGT [26,80]. Therefore, the combined effects of the generation of ROSs and the transfer of ARGs can be predicted to study the impact of NMs on ARGs. The results suggest that the critical factor determining whether NMs facilitate the propagation of ARGs or not is the synergistic effect.
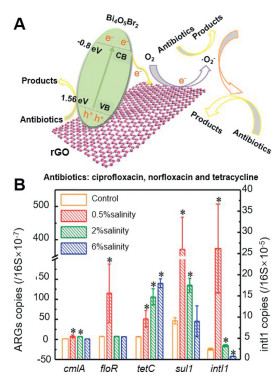
Furthermore, the dynamics for the transmission of ARGs are further complicated by the coexistence of various pollutants in the environment, which also affects the HGT of ARGs under different environmental conditions. For instance, compared to control conditions, the relative abundance of intI1 was found to be significantly higher at low salinities (0.5% and 2%), suggesting that lower salinity levels enhanced the HGT of ARGs, whereas higher salinity (6%) inhibited the spread of genes such as cmlA and floR (Fig. 8B) [131]. Guo et al. investigated the regional and seasonal distributions of ARGs in sediments from the Yangtze River Estuary [132], and reported that most ARGs exhibited significant seasonal fluctuations in average abundance, which was comparatively higher in summer and lower in winter. Therefore, further studies on the HGT of ARGs in practical settings are necessary.
As a typical emerging contaminant, the dissemination of ARGs poses concerns about the safety of environment and human health. Elucidating the transmission mechanism of ARGs is essential for controlling their spread in the environment. An increasing body of research has demonstrated that NMs enhance the HGT of ARGs by increasing the permeability of cell membrane, forming nanopores on the surfaces of membranes, and inducing oxidative stress responses. It is worth mentioning that NMs can simultaneously mediate the elimination and HGT of ARGs. Given that the concentration and size of NMs significantly impact the HGT of ARGs and considering the distinct physicochemical features of emerging NMs, it is crucial to investigate the varying potential effects of these novel NMs on the future removal and spread of ARGs. Therefore, we try to make some suggestions for future research on the impact of NMs on the transfer of ARGs.
(1) Recognizing that the widespread application of NMs leads to their inevitable release into the environment, prompting researchers to conduct comprehensive assessments of NMs;
(2) Understanding that the dissemination of ARGs is closely related to the concentration of NMs, highlighting the need to determine the specific concentrations of NMs in various environments before evaluating their impacts on ARGs;
(3) Acknowledging that different NMs possess unique physicochemical properties, necessitating consideration of the risks associated with different types of NMs including the effects of novel NMs on ARGs..
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Yun Bai: Formal analysis, Data curation. Shengnan Li: Software. Shih-Hsin Ho: Writing – review & editing, Supervision, Project administration.
This study was supported by the State Key Laboratory of Urban Water Resource and Environment (Harbin Institute of Technology) (No. 2022TS13), the key projects of National Natural Science Foundation of China (No. 2019YFC0408503), and the Key Research Program of Wuhan (No. 2022022202015015).
Supplementary material associated with this article can be found, in the online version, at doi:
S.F. Nadeem, U.F. Gohar, S.F. Tahir, et al., Crit. Rev. Microbiol. 46 (2020) 578–599. doi: 10.1080/1040841x.2020.1813687
R.S. Peixoto, C.R. Voolstra, M. Sweet, et al., Nat. Microbiol. 7 (2022) 1726–1735. doi: 10.1038/s41564-022-01173-1
E.Y. Klein, T.P. Van Boeckel, E.M. Martinez, et al., Proc. Natl. Acad. Sci. U. S. A. 115 (2018) 3463–3470.
H. Inoue, Glob. Hlth. Med. Jpn. 1 (2019) 61–64. doi: 10.35772/ghm.2019.01026
J.Q. Su, B. Wei, W.Y. Ou-Yang, et al., Environ. Sci. Technol. 49 (2015) 7356–7363. doi: 10.1021/acs.est.5b01012
H.Y. Zou, L.Y. He, F.Z. Gao, et al., Sci. Total Environ. 772 (2021) 145516. doi: 10.1016/j.scitotenv.2021.145516
L. Li, J. Nesme, M. Quintela-Baluja, et al., Environ. Sci. Technol. 55 (2021) 5939–5949. doi: 10.1021/acs.est.0c08548
S.M. Zainab, M. Junaid, N. Xu, et al., Water Res. 187 (2020) 116455. doi: 10.1016/j.watres.2020.116455
W. Zieliński, E. Korzeniewska, M. Harnisz, et al., Environ. Int. 156 (2021) 106641. doi: 10.1016/j.envint.2021.106641
T. Cen, X. Zhang, S. Xie, et al., Environ. Int. 138 (2020) 105544. doi: 10.1016/j.envint.2020.105544
Y. Zhang, A.Z. Gu, M. He, et al., Environ. Sci. Technol. 51 (2016) 570–580.
G. Li, X. Chen, H. Yin, et al., Environ. Int. 136 (2020) 105497. doi: 10.1016/j.envint.2020.105497
S.S. Ali, R. Al-Tohamy, E. Koutra, et al., Sci. Total Environ. 792 (2021) 148359. doi: 10.1016/j.scitotenv.2021.148359
M. Azizi-Lalabadi, F. Garavand, S.M. Jafari, Adv Colloid. Interfac. 293 (2021) 102440. doi: 10.1016/j.cis.2021.102440
T. Kuang, L. Deng, S. Shen, et al., Chin. Chem. Lett. 34(2023)108584. doi: 10.1016/j.cclet.2023.108584
D. Cardillo, V. Sencadas, T. Devers, et al., Chem. Eng. J. 405 (2020)126843.
M. Chaki Borrás, R. Sluyter, P.J. Barker, et al., J. Photoch. Photobio. B 207 (2020) 111883. doi: 10.1016/j.jphotobiol.2020.111883
R. Mukhopadhyay, B. Sarkar, E. Khan, et al., Crit. Rev. Env. Sci. Tec. 52 (2022) 2611–2660. doi: 10.1080/10643389.2021.1886891
H. Zhan, Y. Wang, X. Mi, et al., Chin. Chem. Lett. 31 (2020) 2843–2848. doi: 10.1016/j.cclet.2020.08.015
M. Aslam, A. Rani, B.N. Pant, et al., Recent development in the production and utilization of plant biomass-based nanomaterials„ in: S.K. Tiwari, M. Bystrzejewski, V. Kumar (Eds. ), Biomass-Based Functional Carbon Nanostructures for Supercapacitors, Springer Nature Singapore, Singapore, 2023, pp. 331–368.
H. Cui, A.L. Smith, Environ. Res. 204 (2021)112373.
A.A. Keller, A. Lazareva, Environ. Sci. Tech. Lett. 1 (2013) 65–70.
C. Zhang, R. Sun, T. Xia, Nano Today 34 (2020) 100909. doi: 10.1016/j.nantod.2020.100909
L.M. Stabryla, K.A. Johnston, N.A. Diemler, et al., Nat. Nanotechnol. 16 (2021) 996–1003. doi: 10.1038/s41565-021-00929-w
Y. Zhang, A.Z. Gu, S. Xie, et al., Environ. Int. 121 (2018) 1162–1171. doi: 10.1016/j.envint.2018.10.030
K. Yu, F. Chen, L. Yue, et al., Environ. Sci. Technol. 54 (2020) 10012–10021. doi: 10.1021/acs.est.0c01870
X. Lu, J. Hou, K. Yang, et al., Environ. Sci. Technol. 55 (2021) 9305–9316. doi: 10.1021/acs.est.1c02047
Q. Zhang, X. Liu, H. Zhou, et al., Chem. Eng. J. 450 (2022) 138301. doi: 10.1016/j.cej.2022.138301
Q. Zhang, H. Zhou, P. Jiang, et al., J. Hazard. Mater. 469 (2024) 133942. doi: 10.1016/j.jhazmat.2024.133942
S. Zhang, J. Lu, Y. Wang, et al., J. Hazard. Mater. 421 (2021) 126728.
E. Marti, E. Variatza, J. Luis Balcazar, Trends Microbiol. 22 (2014) 36–41. doi: 10.1016/j.tim.2013.11.001
A. Pruden, M. Arabi, H.N. Storteboom, Environ. Sci. Technol. 46 (2012) 11541–11549. doi: 10.1021/es302657r
N. Fouz, K.N.A. Pangesti, M. Yasir, et al., Trop Med. Infect. Dis. 5 (2020) 33. doi: 10.3390/tropicalmed5010033
Z. Wei, K. Feng, Z. Wang, et al., Environ. Sci. Technol. 55 (2021) 11824–11834. doi: 10.1021/acs.est.1c01250
W. Ben, J. Wang, R. Cao, et al., Chemosphere 172 (2017) 392–398. doi: 10.1016/j.chemosphere.2017.01.041
B. Chen, Y. Yang, X. Liang, et al., Environ. Sci. Technol. 47 (2013) 12753–12760. doi: 10.1021/es403818e
C. Jiang, Z. Zhao, D. Zhu, et al., Water Res. 249 (2023) 120911.
S. Jia, X. -X. Zhang, Y. Miao, et al., Water Res. 124 (2017) 259–268. doi: 10.1016/j.watres.2017.07.061
J. -H. Wang, J. Lu, J. Wu, et al., Environ. Pollut. 248 (2019) 462–470. doi: 10.1016/j.envpol.2019.02.062
B. Zhang, S. Qin, X. Guan, et al., Water Res. 203 (2021) 117507. doi: 10.1016/j.watres.2021.117507
D. Wu, Z. Huang, K. Yang, et al., Environ. Sci. Technol. 49 (2015) 4122–4128. doi: 10.1021/es506081z
J. Wang, J. Lu, J. Wu, et al., Chemosphere 301 (2022) 134718. doi: 10.1016/j.chemosphere.2022.134718
B. Shao, Z. Liu, L. Tang, et al., J. Hazard. Mater. 435 (2022) 129067. doi: 10.1016/j.jhazmat.2022.129067
R. Pallares-Vega, H. Blaak, R. van der Plaats, et al., Water Res. 161 (2019) 319–328. doi: 10.1016/j.watres.2019.05.100
I.D. Rafraf, I. Lekunberri, A. Sànchez-Melsió, et al., Environ. Pollut. 219 (2016) 353–358. doi: 10.1016/j.envpol.2016.10.062
N.A. Sabri, H. Schmitt, B. Van der Zaan, et al., J. Environ. Chem. Eng. 8 (2020) 102245. doi: 10.1016/j.jece.2018.03.004
S. Liu, P. Wang, C. Wang, et al., Water Res. 202 (2021) 117447. doi: 10.1016/j.watres.2021.117447
H.C. Su, Y.S. Liu, C.G. Pan, et al., Sci. Total Environ. 616 (2018) 453–461.
Z.C. Zhou, J. Zheng, Y.Y. Wei, et al., Environ. Sci. Pollut. R. 24 (2017) 1–10. doi: 10.1007/s11356-015-5582-4
H. Jia, H. Qian, L. Zheng, et al., Sci. Total Environ. 717 (2020) 137170. doi: 10.1016/j.scitotenv.2020.137170
E. O'Flaherty, C.M. Borrego, J.L. Balcázar, et al., Sci. Total Environ. 616 (2018) 1356–1364.
T. Zhang, K. Lv, Q. Lu, et al., J. Environ Sci. 104 (2021) 415–429. doi: 10.1016/j.jes.2020.12.023
H.Q. Anh, T.P.Q. Le, N. Da Le, et al., Sci. Total Environ. 764 (2021) 142865. doi: 10.1016/j.scitotenv.2020.142865
J.H. Wang, J. Lu, Y.X. Zhang, et al., Bioresource Technol. 253 (2018) 235–243. doi: 10.1016/j.biortech.2018.01.035
K. Qin, L. Wei, J. Li, et al., Chin. Chem. Lett. 31 (2020) 2603–2613. doi: 10.1016/j.cclet.2020.04.057
X. Shi, Y. Xia, W. Wei, et al., Water Res. 224 (2022) 119060. doi: 10.1016/j.watres.2022.119060
M. Brown-Jaque, W. Calero-Cáceres, M. Muniesa, Plasmid 79 (2015) 1–7. doi: 10.1016/j.plasmid.2015.01.001
A. Chevallereau, B.J. Pons, S. van Houte, et al., Nat. Rev. Microbiol. 20 (2022) 49–62. doi: 10.1038/s41579-021-00602-y
Y. Yang, S. Xing, Y. Chen, et al., J. Hazard. Mater. 404 (2021) 124149. doi: 10.1016/j.jhazmat.2020.124149
P. Lucas, E. Jouy, L. Le Devendec, et al., Vet. Microbiol. 224 (2018) 100–106. doi: 10.1016/j.vetmic.2018.08.005
Y. Che, Y. Yang, X. Xu, et al., Proc. Natl. Acad. Sci. U. S. A. 118 (2021) e2008731118. doi: 10.1073/pnas.2008731118
S. Sharma, A. Chauhan, A. Ranjan, et al., Front. Microbiol. 15 (2024) 1403168. doi: 10.3389/fmicb.2024.1403168
O'Donnell M., Mann J.F., Schutte A.E., et al., New Engl. J. Med. 375 (2016) 2404–2408. doi: 10.1056/NEJMc1612304
Y. Zhang, Y. Guo, T. Qiu, et al., Front. Microbiol. 13 (2022) 936267. doi: 10.3389/fmicb.2022.936267
M. Touchon, J.A. Moura de Sousa, E.P.C. Rocha, Curr. Opin. Microbiol. 38 (2017) 66–73. doi: 10.1016/j.mib.2017.04.010
C.M. Rands, E.V. Starikova, B. Harald, et al., Environ. Microbiol. 20 (2018) 2288–2300. doi: 10.1111/1462-2920.14276
Y. Ji, H. Xi, Z. Zhao, et al., Sci. Total Environ. 859 (2023) 160304. doi: 10.1016/j.scitotenv.2022.160304
Y. Zou, M. Wu, J. Liu, et al., J. Hazard. Mater. 429 (2022) 128275. doi: 10.1016/j.jhazmat.2022.128275
Y. Han, J. Wang, Z. Zhao, et al., Environ. Pollut. 242 (2018) 1711–1719. doi: 10.1016/j.envpol.2018.07.101
S. Raza, K.H. Kang, J. Shin, et al., Chemosphere 313 (2023) 137362. doi: 10.1016/j.chemosphere.2022.137362
Z. Chen, Q. Fu, Q. Wen, et al., J. Hazard. Mater. 423 (2021) 127149.
N. Wu, S. Xie, M. Zeng, et al., Sci. Total Environ. 744 (2020) 140920. doi: 10.1016/j.scitotenv.2020.140920
Z. Qiu, Y. Yu, Z. Chen, et al., Proc. Natl. Acad. Sci. 109 (2012) 4944–4949. doi: 10.1073/pnas.1107254109
C.Z. Ding, J. Pan, M. Jin, et al., Nanotoxicology 10 (2016) 1051–1060. doi: 10.3109/17435390.2016.1161856
J. Lu, Y. Wang, M. Jin, et al., Water Res. 169 (2020) 115229. doi: 10.1016/j.watres.2019.115229
A. Markowicz, S. Borymski, A. Adamek, et al., Environ. Res. 223 (2023) 115420. doi: 10.1016/j.envres.2023.115420
Y.R. Chen, X.P. Guo, J.N. Feng, et al., Environ. Sci.: Nano 6 (2019) 2405–2419. doi: 10.1039/c9en00338j
S. Zhang, Y. Wang, H. Song, et al., Environ. Int. 129 (2019) 478–487. doi: 10.1016/j.envint.2019.05.054
M.O. Ojemaye, M.A. Adefisoye, A.I. Okoh, J. Environ. Manage. 275 (2020) 111234. doi: 10.1016/j.jenvman.2020.111234
Y. Su, D. Wu, H. Xia, et al., Environ. Int. 128 (2019) 407–416. doi: 10.1016/j.envint.2019.05.007
H. Wang, H. Qi, M. Zhu, et al., Ecotox. Environ. Safe. 186 (2019) 109781. doi: 10.1016/j.ecoenv.2019.109781
W. Xue, X. Shi, J. Guo, et al., Water Res. 253 (2024) 121309. doi: 10.1016/j.watres.2024.121309
J. Ma, J. Gu, X. Wang, et al., Bioresource Technol. 289 (2019) 121688. doi: 10.1016/j.biortech.2019.121688
H. Zhang, W. Guan, J. Shu, et al., Sci. Total Environ. 881 (2023) 163520. doi: 10.1016/j.scitotenv.2023.163520
H. Sun, H. Chang, Y. Zhu, et al., J. Hazard. Mater. 470 (2024) 134128. doi: 10.1016/j.jhazmat.2024.134128
X. Han, P. Lv, L. -G. Wang, et al., Environ. Sci.: Nano 7 (2020) 1214–1224. doi: 10.1039/c9en01279f
G. Feng, H. Huang, Y. Chen, J. Hazard. Mater. 420 (2021) 126602. doi: 10.1016/j.jhazmat.2021.126602
M.T. Guo, X.B. Tian, J. Hazard. Mater. 380 (2019) 120877. doi: 10.1016/j.jhazmat.2019.120877
J. Shi, D. Wu, Y. Su, et al., Environ. Sci.: Nano 7 (2020) 3536–3546. doi: 10.1039/d0en00511h
C.S. Zhou, J.W. Wu, B.F. Liu, et al., J. Hazard. Mater. 431 (2022) 128547. doi: 10.1016/j.jhazmat.2022.128547
L. Xu, J. Zhao, Z. Liu, et al., Sci. Total Environ. 695 (2019) 133932. doi: 10.1016/j.scitotenv.2019.133932
Y. Liu, J. Gao, Y. Wang, et al., J. Hazard. Mater. 432 (2022) 128722. doi: 10.1016/j.jhazmat.2022.128722
X. Wang, F. Yang, J. Zhao, et al., NanoImpact. 10 (2018) 61–67. doi: 10.1117/12.2294518
X. Liu, J. Tang, B. Song, et al., Nanotoxicology 13 (2019) 1422–1436. doi: 10.1080/17435390.2019.1669731
Q. Zhang, H. Zhou, J. Qiao, et al., Chem. Eng. J. 486 (2024) 150276. doi: 10.1016/j.cej.2024.150276
C. Jin, J. Cao, K. Zhang, et al., Ecotox. Environ. Safe. 256 (2023) 114913. doi: 10.1016/j.ecoenv.2023.114913
X. Hu, X. Sheng, W. Zhang, et al., Environ. Sci. Tech. Lett. 7 (2020) 421–427. doi: 10.1021/acs.estlett.0c00311
W. Zou, X. Li, Z. Lai, et al., ACS Appl. Mater. Interfaces 8 (2016) 33165–33174. doi: 10.1021/acsami.6b09981
X. Guanghui, L. Yanjun, L. Xiaolong, et al., Sci. Total Environ. 912 (2024) 169293. doi: 10.1016/j.scitotenv.2023.169293
Q.R. Zhang, H.X. Zhou, P. Jiang, et al., J. Hazard. Mater. 455 (2023) 131658. doi: 10.1016/j.jhazmat.2023.131658
Q. Jiang, M. Feng, C. Ye, et al., Sci. Total Environ. 806 (2022) 150568. doi: 10.1016/j.scitotenv.2021.150568
M.T. Guo, G.S. Zhang, Chemosphere 183 (2017) 197–203. doi: 10.1016/j.chemosphere.2017.04.145
W. Kong, H. Li, J. Liu, et al., J. Mater. Chem. B 3 (2015) 1198–1203. doi: 10.1039/C4TB02096K
M. Zhuo, J. Ma, X. Quan, Chemosphere 281 (2021) 130865. doi: 10.1016/j.chemosphere.2021.130865
J.A. Lemire, J.J. Harrison, R.J. Turner, Nat. Rev. Microbiol. 11 (2013) 371–384. doi: 10.1038/nrmicro3028
N. Garg, P. Bhandari, Plant Biosys. 148 (2013) 609–621.
N.A. Smirnov, S.I. Kudryashov, A.A. Nastulyavichus, et al., Laser Phys. Lett. 8 (2018) 102245.
A.B. Djurišić, Y.H. Leung, A.M.C. Ng, et al., Small 11 (2014) 26–44.
A. Blessing, Molecules 22 (2017) 1370. doi: 10.3390/molecules22081370
H. Zhang, L. Xu, X. Hou, et al., Environ. Pollut. 360 (2024) 124676. doi: 10.1016/j.envpol.2024.124676
S. Yu, Y. Wang, F. Shen, et al., Sci. Total Environ. 815 (2022) 152885. doi: 10.1016/j.scitotenv.2021.152885
N. Rajkumari, S. Alex, K.B. Soni, et al., 3 Biotech. 11 (2021) 497.
E. Eroglu, P.M. Tiwari, A.B. Waffo, et al., Int. J. Nanomed. 8 (2013) 1403–1415.
Z.P. Xu, Q.H. Zeng, G.Q. Lu, et al., Chem. Eng. Sci. 61 (2006) 1027–1040. doi: 10.1016/j.ces.2005.06.019
A. Pudlarz, J. Szemraj, Open Life Sci. 13 (2018) 285–298. doi: 10.1515/biol-2018-0035
L. Chang, H. Yan, J. Chang, et al., Eur. Polym. J. 156 (2021) 110593. doi: 10.1016/j.eurpolymj.2021.110593
X. Zhao, H. Cui, W. Chen, et al., PloS One 9 (2014) e98919. doi: 10.1371/journal.pone.0098919
J.W. Beaber, B. Hochhut, M.K. Waldor, Nature 427 (2004) 72–74. doi: 10.1038/nature02241
D.J. Dwyer, M.A. Kohanski, J.J. Collins, Curr. Opin. Microbiol. 12 (2009) 482–489. doi: 10.1016/j.mib.2009.06.018
A.F. Seixas, A.P. Quendera, J.P. Sousa, et al., Front. Genet. 12 (2022) 821535. doi: 10.3389/fgene.2021.821535
A.S. Ezeuko, M.O. Ojemaye, O.O. Okoh, et al., J. Water Process Eng. 41 (2021) 102041. doi: 10.1016/j.jwpe.2021.102041
M. Cho, H. Chung, W. Choi, et al., Appl. Environ. Microb. 71 (2005) 270–275. doi: 10.1128/AEM.71.1.270-275.2005
Sicwetsha, Simbongile, Mvango, et al., J. Nanopart Res. 23 (2021) 1–18. doi: 10.1007/s11051-020-05135-8
Z. Zhang, Y. Chen, Chem. Eng. J. 382 (2020) 122955. doi: 10.1016/j.cej.2019.122955
X. Sun, B. Chen, Q. Li, et al., Sci. Total Environ. 642 (2018) 1378–1385. doi: 10.1016/j.scitotenv.2018.06.141
B. Halliwell, A. Adhikary, M. Dingfelder, et al., Chem. Soc. Rev. 50 (2021) 8355–8360. doi: 10.1039/d1cs00044f
Z. Xu, D. Zhao, J. Lu, et al., Chem. Eng. J. 455 (2023) 140927. doi: 10.1016/j.cej.2022.140927
X. Zheng, S. Xu, Y. Wang, et al., J. Colloid Interf. Sci. 527 (2018) 202–213. doi: 10.1016/j.jcis.2018.05.054
M. Xu, Y. Wang, E. Ha, et al., Chemosphere 265 (2020) 129013.
Q. Zhang, H. Zhou, P. Jiang, et al., J. Hazard. Mater. 455 (2023) 131658. doi: 10.1016/j.jhazmat.2023.131658
N. Guo, Y. Wang, T. Tong, et al., Water Res. 133 (2018) 79–86. doi: 10.1016/j.watres.2018.01.020
X.P. Guo, X. Liu, Z.S. Niu, et al., Environ. Pollut. 242 (2018) 576–584. doi: 10.1016/j.envpol.2018.06.099

Figure 2 Various mechanisms for the spread of ARGs: (A) transformation (B) transduction (C) conjugation. (D) Schematic of the distribution of antibiotic-resistant MGEs, requiring close cell-to-cell contact for successful transfer of ARGs. Herein, GI represents the genomic island, ICE represents the integrative conjugative elements, GTA represents the gene transfer agent, and Tn represents the transposon. Reproduced with permission [57]. Copyright 2015, Elsevier.

Figure 3 (A) Transformation frequency under different concentrations of nano-ZnO. Modified with permission [76]. Copyright 2023, Elsevier. (B) Schematic of possible mechanisms by which exposure to NMs affects the fate of ARGs during the proliferation of microbial populations. Reproduced with permission [80]. Copyright 2019, Elsevier. (C) Absolute abundances of MGEs on Days 0 and 43. Reproduced with permission [83]. Copyright 2019, Elsevier.

Figure 4 (A) Factors affecting the nano-TiO2-mediated phage transduction (ratio of gM13/TG1. Modified with permission [86]. Copyright 2020, Royal Society of Chemistry. (B) Risk of the propagation of ARGs as simulated by NMs. Reproduced with permission [90]. Copyright 2022, Elsevier. (C) Inhibition of extracellular ARGs by GO nanosheets. Modified with permission [91]. Copyright 2019, Elsevier.

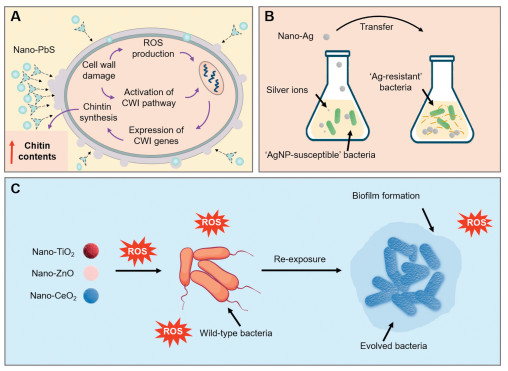
Figure 5 Development of resistance in relation to the physicochemical properties of NMs. (A) Nano-PbS particles can damage the cell wall. (CWI; cell wall integrity). (B) Resistance to nano-Ag in E. coli and P. aeruginosa results from the leakage of flagellin. (C) Sublethal exposure to NMs (TiO2, ZnO, CeO2) promotes the formation of bacterial biofilm. Modified with permission [100]. Copyright 2023, Elsevier.

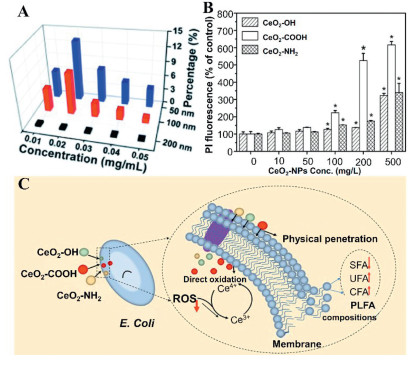
Figure 6 (A) Impacts of various sizes of nano-SiO2 on the permeability of membrane under various concentrations. Modified with permission [103]. Copyright 2015, Royal Society of Chemistry. (B) Membrane permeability of E. coli after treatment by various types of CeO2—NPs in NaCl solution for 6 h. Modified with permission [104]. Copyright 2021, Elsevier. (C) Schematic of the cytotoxicity mechanisms of E. coli induced by CeO2—NPs (UFA: unsaturated fatty acid, SFA: Saturated fatty acids, phospholipid fatty acid (PLFA), Conjugated fatty acids). Reproduced with permission [104]. Copyright 2021, Elsevier.

Figure 7 (A) Schematic of the nano-resistance for antibiotic resistance. Modified with permission [23]. Copyright 2020, Elsevier. (B) Horizontal transfer of phage-mediated ARGs. Modified with permission [64]. Copyright 2022, Elsevier. (C) Mechanisms of CuO NPs developed conjugative transfer of RP4 plasmid [78]. Copyright 2019, Elsevier.

Table 1. Examples of various ARGs detected in various aquatic environments.
| Environment | ARGs | MGEs | Remarks | Ref. |
| WWTP effluent | tetA, tetC, tetG, tetX, tetM, tetO, tetW, sul1, sul2 | intI2, intI1 | – | [35] |
| South China Sea | macB, acrB | – | Pristine environment with minimal human activity | [36] |
| Yangtze River | rosA, rosB, macB, bacA, acrB, adeB, cpxR, amrB, ceoB, cmeB, mdtB, mdtC, mdtF, tetP, mexB, mexC, mexD, mexE, oprC, mexF, mexI, mexW, qepA, vanR | – | Freshwater ecosystems | [37] |
| Livestock breeding wastewater | ant(6′)-Ia, tetX, aac(6′)-Ie, tetM, aph(3′)-IIIa, tetW, tetQ, ant(3″)-Ia, tetO | – | Pig farming wastewater | [38] |
| Mariculture system | tetB, tetG, tetX, qnrS, sul1, sul2, ermF, ermT, qnrA, qnrB, | intI1 | – | [39] |
| Karst River | aadA-01, sul2, aadA-02, blaOXA-21, aadA1, tetG, emrA, aadA2–03, qacEdelta1–01, mdtK, ereA, qacEdelta1–02, | tnpA-04, tnpA-01, tnpA-02, tnpA-03, intI1, cintI1, Tp614, tnpA-05, tnpA-07, IS613 | Drinking water source | [40] |
| Municipal solid waste leachates | sul1, sul2, tetM, tetQ, ermB, mefA | – | – | [41] |
| North Yellow Sea of China | sul1, sul2, tetM, tetX, tetB, blaTEM, qnrS, qnrA, qnrB, qnrF, tetG, ermF, ermT | intI1 | Coastal areas with dense human activities | [42] |
 下载: 导出CSV
下载: 导出CSV
Table 2. Typical NMs mediating the HGT of ARGs.
| NMs | Physicochemical properties | HGT pathways | ARGs | Outcomes | Assessment endpoints | Ref. |
| Al2O3 | Metal oxide NMs | Conjugation | PBR322-based plasmid | Nano-Al2O3 improved the transmission of extracellular ARGs into bacteria through transformation. | Plasmid uptake, generation of ROSs, and permeability of cell membrane. | [94] |
| ZnO | Metal oxide NMs | Conjugation | Plasmid (pGEX4T-1) | The rate of transfer of RP4 plasmid increased in both the pure culture of E. coli and the mixed culture of native aquatic microbiota on exposure to zinc oxide nanoparticles. More precisely, compared to the non-ZnO nanoparticles-treated group, the transfer frequency of RP4 plasmid increased 24.3-folds in E. coli and 8.3-folds in aquatic microbiota on exposure to nano-ZnO at a concentration of 10 mg/L for 12 h. | Nano-ZnO facilitated the efficiency of transformation by naked plasmid pGEX4T-1 and improved the permeability of cell membrane. | [93] |
| TiO2 | Metal oxide NMs | Transduction | Bacteriophage (hM13) | Simultaneous exposure to humic acid (HA) and nanoscale titanium dioxide (nTiO2) synergistically promoted transduction under ultraviolet (UV)-A irradiation. The large amount of superoxide radicals generated by the photoexcitation of HA-nTiO2 combination increased the permeability of membrane and exacerbated the intracellular oxidative stress, thus accelerating the dispersion of ARGs through phage transduction. | Increased permeability of membrane, and generation of ROSs, induced intracellular oxidative stress. | [95] |
| CuO | Metal oxide NMs | Conjugation | Plasmid (RP4) | Under nano-CuO pressure (1–10 µmol/L), the conjugative transfer frequency of RP4 plasmid increased dramatically. When 5 µmol/L of nano-CuO was administered, the conjugative transfer frequency was more than twice that in the control groups. Conversely speaking, for the nano-CuO dosages of higher than 100 µmol/L, the rates of conjugative transfer of ARGs drastically dropped. | Overproduction of ROSs, permeability of membrane, RP4 gene expression, induced intracellular oxidative stress, ATP synthesis. | [78] |
| Ag | Metal NMs | Conjugation | Plasmid (RP4) | Conjugative transfer frequency of RP4 plasmid increased considerably on exposure to nano-Ag at the concentrations of 0.1 and 100 µg/L. Nevertheless, when the concentration of nano-Ag was increased to 1000 µg/L, the frequency of binding transfer of RP4 plasmids decreased significantly. | Overproduction of ROSs, permeability of membrane, RP4 gene expression, ATP synthesis. | [75] |
| Fe2O3@MoS2 | Nanocomposite materials | Conjugation | Plasmid (RP4–7) | Fe2O3@MoS2 nanocomposites significantly inhibited the conjugative transfer of RP4–7 by up to 100-folds. When the concentration of Fe2O3@MoS2 was lower than 100 mg/L, the inhibition of CT increased with the increase of the concentration of Fe2O3@MoS2 from 0 to 100 mg/L. | Suppression of plasmid horizontal transfer by inhibiting the expression of related genes (trbA, trbB, traF, and trfA). | [81] |
| Nanoplastics | Non-metallic NMs | Conjugation | Plasmid (RP4) | The 100 nm-sized Polystyrene (PS) has limited promotion of microorganisms due to its inhibitory effect on microbes. | PS particles altered the permeability of cell membrane and the activity of cell. | [86] |
| CeO2 | Metal oxide NMs | Conjugation | Plasmid (RP4) | In relatively high concentrations of nano-CeO2, the highly facilitated conjugation process contributed to the enhanced transfer of ARGs. Low concentrations of nano-CeO2 inhibited the conjugation process. At relatively high concentrations of nano-CeO2 (25 and 50 mg/L), the conjugative transfer of RP4 plasmids was enhanced by 118%–123%. In contrast, at low concentrations (1 and 5 mg/L), nano-CeO2 hindered the transfer by 22%–26%. | Production of ROSs, expression of plasmid-encoded genes. | [26] |
| TiO2/Ag/GO | Nanocomposite materials | Conjugation | ARB | 100 mg/L of TiO2/Ag/GO achieved 87.8%, 40.9%, and 56.4% inactivation on three bacteria (E. coli HB10663 and E. coli HB10667 and E. coli HB101) | The generation of ROSs induced intracellular oxidative stress. direct touch that oxidized the cell membrane and walls, and altered antioxidant enzymes. | [88] |
| MoS2 | Nanocomposite materials | Conjugation | Plasmid (RP4) | MoS2 significantly promoted the plasmid-mediated horizontal transfer of multiple ARGs. | Production of ROSs, altered ATP synthesis, and expression of related genes. | [96] |
| ZnS | Natural sphalerite | Conjugation | – | Natural sphalerite in lower concentrations (0.05–50 mg/L) facilitated the conjugation of ARGs, however higher concentrations (≥ 100 mg/L) prevented it. | Generation of ROSs, induced intracellular oxidative stress, membrane permeability, and conjugation-regulated gene expression. | [12] |
下载: 导出CSV

 扫一扫看文章
扫一扫看文章

扫一扫关注我们

 下载:
下载: