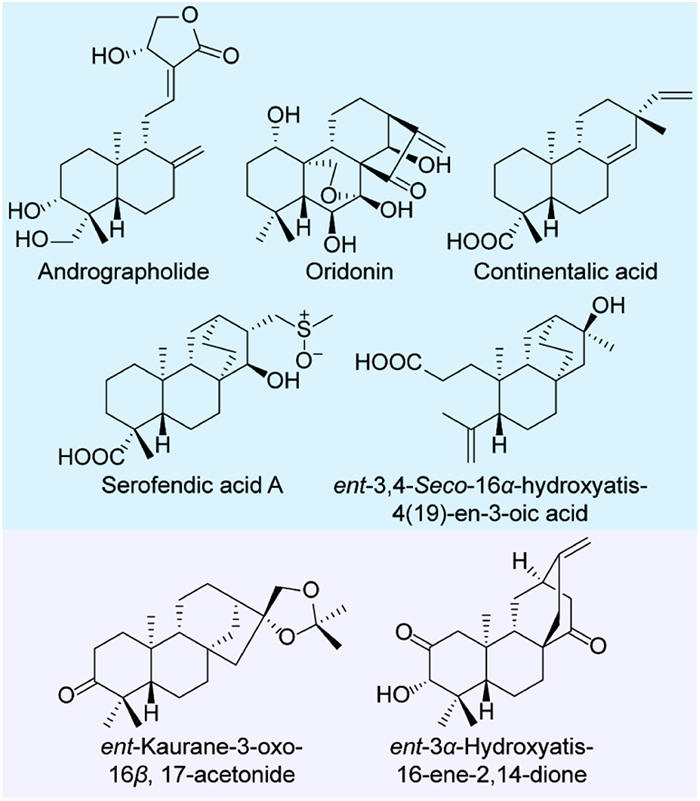
Figure 1.
Examples of active LRDs respectively from E. fischeriana (shadowed in pink) and other species (shadowed in blue).
Mining substrate-promiscuity cytochrome P450s from Euphorbia fischeriana for heterologous bioproduction of diverse labdane-related diterpenoids
Ke Gao , Lin Zhao , Lei Wang , Rui Zhang , Jianxun Zhu , Pengcheng Lin , Jiachen Zi

Labdane-related diterpenoids (LRDs), the largest group of diterpenoids, exhibit a vast structural diversity, including bicyclic structures (e.g., labdane), tricyclic structures (e.g., pimarane and abietane) and tetracyclic structures (e.g., kaurane, atisane, beyerene, and stemodane) [1]. Many LRDs have been used for drug development [2], such as andrographolide (ent–labdane type, an anti-inflammatory drug) [3,4], oridonin (ent–kaurane type, a promising anti-cancer lead) [5], continentalic acid (ent–pimarane type, with a dramatic inhibitory activity against methicillin-resistant Staphylococcus aureus) [6], serofendic acid A (ent–atisane type, a potent neuroprotective agent) [7,8], and ent–3,4-seco-16α-hydroxyatis-4(19)-en-3-oic acid (ent–atisane type, with a significant inhibitory activity against the biofilm adherence of Pseudomonas pseudoalcaligenes) (Fig. 1) [9]. For all these LRDs, oxidative functionalization is required for their bioactivities. In the biosynthesis of plant terpenoids, the olefin precursors are mainly oxidized by cytochrome P450s (CYPs) [10]. Although CYPs involved in the biosynthesis of plant LRDs have been widely characterized, the oxidative functionalization of many sites in the LRDs isolated to date remains unknown. Mining CYPs with novel catalytic properties (e.g., specificity or promiscuity in catalysis and substrate tolerance) can expand the toolkit for bioproduction of diverse LRDs.
Euphorbia fischeriana Steud. is widely distributed in northern China. Its dry roots, known as “langdu”, were used in traditional Chinese medicine for the treatment of dyspepsia, abdominal distension, abdominal pain, cough, and lymph node tuberculosis [11]. Over 180 LRDs have been isolated from its root extracts [12-14]. These LRDs with various skeletons including abietane, pimarane, kaurane, and atisane, play vital roles in the anticancer actions of this plant [15-17]. Because most of them are oxidized at multiple sites, we assume that E. fischeriana is a worthwhile deposit for mining CYPs capable of oxidizing various LRD skeletons. In the previous study, we characterized an ent–copalyl diphosphate (ent–CPP) synthase and four class Ⅰ LRD diterpene synthases (diTPSs) which can respectively produce ent–atiserene (1), ent–kaurene (2), ent–isopimara-7(8),15-diene (3) and ent–neoabietadiene (4) [18].
We performed a phytochemical investigation on E. fischeriana, which led to the isolation of 19 LRDs, including fourteen abietanes, three atisanes, one pimarane and one kaurane (Fig. S1 in Supporting information) [19-21]. The configuration of all the isolated LRDs is ent. This is consistent with the finding that EfTPS54 (an ent–CPP synthase) is the only class Ⅱ LRD diTPS in E. fischeriana [12,13]. The liquid chromatography-tandem mass spectrometry (LC-MS/MS) analysis [22,23] of the isolated LRDs was conducted. The characteristic fragmentation patterns of the LRDs with distinct skeletons were deduced, such as the transformation of α,β-unsaturated γ-lactone (ring D) into 2H-oxete via loss of carbon monoxide (CO) for abietanes, C ring cleavage for atisanes and kauranes, and B ring cleavage via retro Diels-Alder reaction for pimaranes (Fig. S2 in Supporting information). Based on these characteristic fragmentation patterns, a total of 74 LRDs were identified from the extracts of the roots, flowers, leaves, and stems of E. fischeriana (Figs. S1 and S3–S6 in Supporting information), including 46 abietanes, 9 atisanes, 8 pimaranes, 7 kauranes, and 4 abietane-phloroglucinol meroterpenoids. Among them, 66 LRDs (89.2%) were mainly found in the roots. The abundances of 62 LRDs in roots were over ten times higher than those in the other tissues. Whereas stems, leaves, and flowers respectively contain only one predominant LRD with an abundance over ten times higher than those in the other tissues. Therefore, LRDs of E. fischeriana predominantly exist in the root, the part used for medicinal purposes in traditional Chinese medicine. This finding further indicates that LRDs are the important pharmacological ingredients of this plant.
In the previous study, a transcriptome database across various tissues was established [18]. We thus analyzed the correlation between the relative contents of 74 LRDs and the transcript levels of CYP genes across all the tissue samples. Out of 424 CYP transcripts identified from the transcriptome data of E. fischeriana, the levels of 247 were closely correlated with the relative contents of 74 LRDs (Pearson's r = 0.9) (Fig. S7 in Supporting information). 199 CYP transcripts with a length over 700 bp were chosen for further analysis.
Next, we used the previously characterized LRD-related CYPs (Table S1 in Supporting information) as the queries to perform a BLAST search against the transcriptome data of E. fischeriana. The top 10 closest CYPs of each query sequence were selected. A total of 40 CYPs were identified after eliminating the redundant sequences. Among them, 25 were included in the 199 CYPs whose transcript levels were highly correlated with the tissue distribution of LRDs. Phylogenetic analysis showed that all the previously characterized LRD-related CYPs belong to CYP71 and CYP85 clans (Fig. S8 in Supporting information). Among 25 candidate CYPs, 20 locate within CYP71 clan and 5 locate within CYP85 clan (Fig. S8). In particular, 16 CYPs belong to the CYP71 subfamily, and 4 CYPs belong to CYP76 subfamilies (Fig. S8) in which many members are related to LRD biosynthesis [1]. We subsequently sought to characterize the 25 candidate CYPs.
We successfully cloned 22 out of 25 candidate CYP genes. Saccharomyces cerevisiae was firstly used to characterize these CYPs. To increase the production of the potential diterpenoid products in S. cerevisiae, we expressed tHMG1 (the truncated 3–hydroxy-3 methylglutaryl coenzyme A reductase) and UPC2–1 (the G888D mutant of the transcription factor UPC2) to reinforce the mevalonate (MVA) pathway [24,25], expressed two fusion enzymes BTS1 (GGPP synthase)-ERG20 (farnesyl diphosphate synthase) and ERG20-mERG20F96C (the F96C mutate of ERG20) to increase the conversion rate of farnesyl diphosphate (FPP) to geranylgeranyl diphosphate (GGPP) [26], and depressed ERG9 by replacing its native promotor with a glucose-induced promotor PHXT1 to reduce FPP consumption in triterpenoid/steroid biosynthesis [27], thus establishing the strain LRD04 (Table S2 in Supporting information). Then, we respectively expressed EfTPS43 (an E. fischeriana ent–atiserene synthase), EfTPS1 (an E. fischeriana ent–kaurene synthase), EfTPS41 (an E. fischeriana ent–isopimara-7(8),15-diene synthase) and EfTPS14 (an E. fischeriana ent–neoabietadiene synthase) [18] along with ZmAN2 (a Zea mays ent–CPP synthase) [28] in LRD04 to obtain LRD05 to LRD08 (Table S2). A tiny amount of 1 and 2 were detected in LRD05 and LRD06 (Fig. S9 in Supporting information), respectively, but neither 3 nor 4 was detected in LRD07 and LRD08. As diffusion of the intermediates produced by the upstream enzymes may lead to decrease in the concentration surrounding the downstream enzymes, fusion of the consecutive enzymes may bring their active sites into a closer proximity, thereby enhancing metabolic flux channeling to biosynthesis of the target products [26]. It was reported that the fusion of SmCPS (a labdadienyl/copalyl diphosphate synthase from Salvia miltiorrhiza) and SmKSL (a kaurene synthase-like from S. miltiorrhiza) resulted in a significant increase of miltiradiene titer [29]. Hence, we respectively fused EfTPS43, EfTPS1, EfTPS41 and EfTPS14 with ZmAN2, using GSG (glycine–serine-glycine) as the linker, and subsequently expressed the resulting fusion enzymes in LRD04 to increase the titers of the diterpene olefins. LRD09 expressing ZmAN2-GSG-EfTPS43 and LRD10 expressing ZmAN2-GSG-EfTPS1 respectively produced much more 1 and 2 compared to LRD05 and LRD06 (Table S2 and Fig. S9), but 3 and 4 were still not detected.
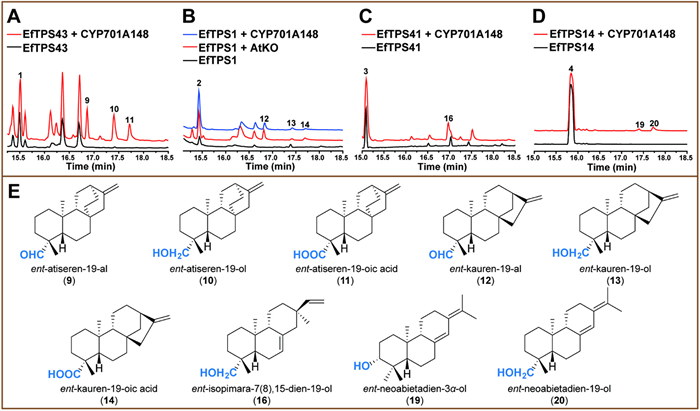
We thus used LRD09 and LRD10 to screen 22 candidate CYPs by expressing each of them associated with the different cytochrome P450 reductases including AaCPR (from Artemisia annua) [25], ATR1 (from Arabidopsis thaliana) [30], SaCPR (from Santalum album L.) [31] and CrCPR (from Catharanthus roseus) [32]. The results showed that CYP71D450 and CYP701A148 oxidized both 1 and 2 (Figs. 2 and 3), and exhibited the highest activities when accompanied by AaCPR.
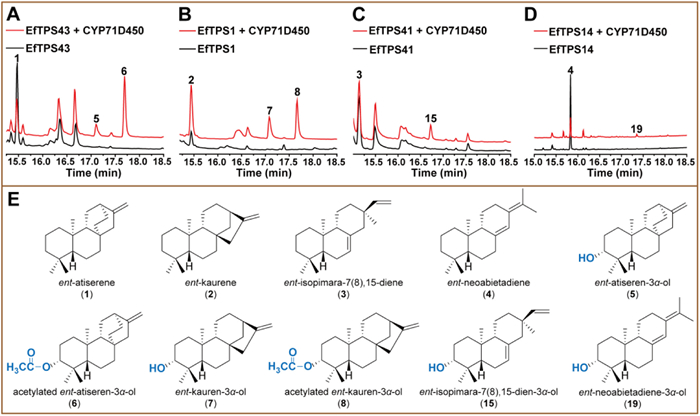
CYP71D450 oxidized 1 into two compounds 5 and 6 with the molecular weights of 288 and 330 Da, respectively (Fig. 2A and Fig. S10 in Supporting information). To further increase the production of 5 and 6 and prepare sufficient amounts of them for structural elucidation, CYP71D450 and AaCPR were integrated into the ROX1 (a yeast transcriptional repressor which can repress the expression of genes involved in terpene biosynthesis in yeast) locus of LRD09 to afford LRD13 (Table S2) [33,34]. After LRD13 was cultured by fed-batch fermentation, 6 was isolated as an orthorhombic crystal and its structure was determined as acetylated ent–atiseren-3α-ol by nuclear magnetic resonance (NMR) and X-ray crystallography. Because the molecular weight of 5 (288 Da) is 42 Da less than that of 6 (330 Da) (Fig. S10), 5 is ent–atiseren-3α-ol. We assumed that 6 was acetylated by a yeast endogenous acetyltransferase as CYPs are incapable of catalyzing acetylation reaction. To confirm this, we expressed CYP71D450 in S. cerevisiae WAT11 harboring ATR1, and prepared microsome to perform in vitro reactions (Fig. S11 in Supporting information). Only 5 was detected from the extract of the reaction mixture, which supported that acetylation of 5 to 6 was catalyzed by a yeast acetyltransferase. CYP71D450 also oxidized 2 to 7 and 8 in S. cerevisiae (Fig. 2B and Fig. S10). After fed-batch fermentation, sufficient 7 was isolated for acquisition of its NMR data which completely match those of ent–kauren-3α-ol [35]. The signal of H-3 in the 1H NMR spectrum of 7 is a doublet of doublets with a big coupling constant of 11.6 Hz and a small coupling constant of 4.9 Hz (Table S3 in Supporting information), suggesting an axial position of H-3, which is consistent with 3α-hydroxylated ent–kauranes [36]. 7 was therefore determined as ent–kauren-3α-ol. And like 5, 7 was also acetylated in S. cerevisiae to yield acetylated ent–kauren-3α-ol (8), with a molecular weight of 330 Da which is 42 Da bigger than that of 7 (Fig. 2B, Figs. S10 and S11).
Expression of CYP701A148 and AaCPR in the ent–atiserene-producing strain LRD09 led to the detection of three products 9, 10 and 11 with the molecular weights of 286, 288 and 302 Da, respectively (Fig. 3A and Fig. S12 in Supporting information). After fed-batch fermentation, sufficient 9 and 10 were purified for NMR analysis and their structures were respectively elucidated as ent–atiseren-19-al [37] and ent–atiseren-19-ol [38], thereby indicating that 11 is ent–atiseren-19-oic acid according to its molecular weight (Fig. S12). This result was also confirmed by in vitro biochemical reaction using the microsome as the catalyst (Fig. S13 in Supporting information).
When CYP701A148 and AaCPR were expressed in the ent–kaurene-producing strain LRD10, three products 12, 13, and 14, also respectively possessing the molecular weights of 286, 288 and 302 Da, were observed (Fig. 3B and Fig. S12). We expressed AtKO (an A. thaliana ent–kaurene oxidase) [39] in LRD10 and found that the retention time and the MS fragment patterns of 12, 13 and 14 are consistent with those of ent–kauren-19-al, ent–kauren-19-ol and ent–kauren-19-oic acid (Fig. 3B). Hence, their structures were determined. Additionally, 12, 13, and 14 were also detected in the reaction mixture of 2 and the microsome containing CYP701A148 (Fig. S13).
As we failed to construct the S. cerevisiae strains producing 3 and 4, we used Nicotiana benthamiana, Escherichia coli and in vitro biochemical reactions to identify CYPs capable of oxidizing 3 and 4. The results showed that both CYP71D450 and CYP701A148 can accept 3 and 4 as their substrates.
For 3, CYP71D450 and CYP701A148 oxidized it into 15 and 16, respectively, in E. coli and the microsome-containing reaction mixtures (Figs. 2C, 3C and Figs. S10–S13). Both 15 and 16 possess a molecular weight of 288 Da (Figs. S10E and S12G). According to the functions of CYP71D450 and CYP701A148 on 1 and 2, we assumed that 15 and 16 might be ent–isopimara-7(8),15–dien-3α-ol and ent–isopimara-7(8),15–dien-19-ol, respectively. Sufficient 15 and 16 were prepared through large-scale fermentation of the corresponding E. coli strains for NMR analysis. The NMR data of 15 and 16 were respectively consistent with those of nor-isopimara-7,15–dien-3β-ol [40] and nor-isopimara-7,15–dien-19-ol [41]. Given that 15 and 16 were derived from ent–isopimaradien-7(8),15-diene (3), their structures were therefore respectively determined as the enantiomers of nor-isopimara-7,15–dien-3β-ol and nor-isopimara-7,15–dien-19-ol, i.e., ent–isopimara-7,15–dien-3α-ol (Fig. 2E) and ent–isopimara-7,15–dien-19-ol (Fig. 3E). Like 7, H-3 of 15 possesses an axial orientation due to its peak of double of doublets (11.4 and 4.3 Hz), which is also consistent with those of 3α-hydroxylated ent–pimaranes [42]. This further confirms the configuration of C-3 of 15.
For 4, CYP701A148 oxidized it into two products, 19 and 20, in the E. coli strain ELRD04 (Fig. 3D and Table S2). Both 19 and 20 possess the same molecular weight of 288 Da (Fig. S12). We prepared sufficient amounts of 19 and 20 for NMR analysis. 20 is ent–neoabietadien-19-ol (Fig. 3E), which is consistent with the function of CYP701A148 on the other three diterpene olefins. Unexpectedly, 19 harbors a hydroxy group at C-3. Like both 7 and 15, the peak of H-3 of 19 is also a doublet of doublets with coupling constants of 11.7 and 3.2 Hz, which is consistent with those of the abietane-type diterpenoids with a 3α–hydroxyl group [43]. And the nuclear Overhauser effect (NOE) correlations between H-3 and H-5 and between H-3 and H3–18 were observed in the NOESY spectrum of 19. Therefore, the orientation of 3-OH of 19 was determined as α. We used N. benthamiana, E. coli and in vitro microsome reaction to characterize the function of CYP71D450 on 4. CYP71D450 only functions in N. benthamiana to hydroxylate 4 into 19 (Fig. 2D).
In addition, CYP88A176 shares a 67.9% sequence identity with AtKAO (an A. thaliana ent–kauren-19-oic acid oxidase) [44]. We therefore presumed that CYP88A176 is an ortholog of AtKAO. We respectively expressed CYP88A176 and AtKAO in LRD17 to afford LRD18 and LRD19 (Table S2). After silylation, we detected the silylated derivatives of ent–kauren-7β-ol-19-oic acid and gibberellin A12 from the extracts of both LRD18 and LRD19 (Fig. S14 in Supporting information). Hence, CYP88A176 is indeed an ent–kauren-19-oic acid oxidase.
Many CYPs involved in LRD biosynthesis exhibit substrate and/or catalytic promiscuity and thereby have been used for synthesis of diverse LRDs via combinatorial biosynthesis [45]. CYP71D450 can catalyze the hydroxylation of C-3 of ent–atiserene, ent–kaurene and ent–isopimara-7(8),15-diene; CYP701A148 can oxidize C-19 of ent–atiserene and ent–kaurene into the carboxyl groups, and hydroxylate C-19 of ent–isopimara-7(8),15-diene and both C-19 and C-3 of ent–neoabietadiene (Fig. S15 in Supporting information). To the best of our knowledge, this study for the first time reports CYPs which can oxidize C-3 of ent–isopimara-7(8),15-diene, ent–atisane and ent–abietane skeletons, and C-19 of ent–abietane and ent–pimarane skeletons. As illustrated in Fig. 1 and Fig. S15, the discovery of these CYPs may enable enzymatic/chemoenzymatic synthesis of not only anti-cancer LRDs from E. fischeriana, such as ent–3α-hydroxyatis-16-ene-2,14–dione [46] and ent–kaurane-3-oxo-16β,17-acetonide [47], but also other valuable ones, such as serofendic acid A [7,8], 7β,11α,17-trihydroxy-helioscopinolide E [48] and araucarone [49], thereby enriching the toolkit for combinatorial biosynthesis of diverse unnatural LRDs for drug development.
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Ke Gao: Writing – original draft, Investigation, Formal analysis, Data curation. Lin Zhao: Writing – original draft, Methodology, Investigation, Formal analysis. Lei Wang: Methodology. Rui Zhang: Methodology. Jianxun Zhu: Writing – original draft, Methodology, Investigation, Formal analysis, Conceptualization. Pengcheng Lin: Writing – original draft, Investigation, Funding acquisition, Conceptualization. Jiachen Zi: Writing – original draft, Project administration, Investigation, Funding acquisition, Formal analysis, Data curation, Conceptualization.
We thank Dr. David R. Nelson for annotation of cytochrome P450 enzymes. We thank Dr. Xiaoya Shang from Beijing Union University for LC-MS/MS analysis. The research was supported by grants from the National Natural Science Foundation of China [Nos. 82474024, 82293682 (82293680), 82073953], the Nonprofit Central Research Institute Fund of Chinese Academy of Medical Sciences (No. 2021-RC350–009), the CAMS Innovation Fund for Medical Sciences (No. 2023-I2M-2–006), and the Special Fund for the Construction of Qinghai Innovation Platform (No. 2024-ZJ-T02).
Supplementary material associated with this article can be found, in the online version, at doi:
K. Gao, W.L. Zha, J.X. Zhu, C. Zheng, et al., Chin. J. Nat. Med. 19 (2021) 666–674. doi: 10.1039/D4TA09029B
Y. Wang, P. Tang, W. Tu, Q. Gao, et al., Chin. Chem. Lett. 36 (2025) 109955. doi: 10.1016/j.cclet.2024.109955
B. Zeng, A.L. Wei, Q. Zhou, M.H. Yuan, et al., Phytother. Res. 36 (2022) 336–364. doi: 10.1002/ptr.7324
X.H. Chen, J.Z. Ren, J.D. Yang, Z.P. Zhu, et al., Medi. Plant Biol. 2 (2023) 15.
X. Li, C.T. Zhang, W. Ma, X. Xie, et al., Front. Pharmacol. 12 (2021) 645824. doi: 10.3389/fphar.2021.645824
S.I. Jeong, W.S. Han, Y.H. Yun, K.J. Kim, Phytother. Res. 20 (2006) 511–514. doi: 10.1002/ptr.1894
T. Kume, N. Asai, H. Nishikawa, N. Mano, et al., Proc. Natl. Acad. Sci. U. S. A. 99 (2002) 3288–3293. doi: 10.1073/pnas.052693999
M. Akao, T. Takeda, T. Kita, T. Kume, et al., Cardiovasc. Drug Rev. 25 (2007) 333–341. doi: 10.1111/j.1527-3466.2007.00026.x
Z.C. Wang, Y.M. Lin, D.Q. Feng, C.H. Ke, et al., Molecules 14 (2009) 414–422. doi: 10.3390/molecules14010414
X. Zheng, P. Li, X. Lu, J. Exp. Bot. 70 (2019) 4619–4630. doi: 10.1093/jxb/erz203
Jiangsu New Medical College, Dictionary of Chinese Materia Medica, Shanghai Science and Technical Press, Shanghai, 1986, pp. 1898–1900.
B. Jian, H. Zhang, J. Liu, Molecules 23 (2018) 935. doi: 10.3390/molecules23040935
Z.J. Zhan, S. Li, W. Chu, S. Yin, Nat. Prod. Rep. 39 (2022) 2132–2174. doi: 10.1039/d2np00047d
Y. Peng, Y. Chang, C. Sun, W. Wang, et al., Chin. Chem. Lett. 33 (2022) 4261–4263. doi: 10.1016/j.cclet.2022.03.003
J. Sang, L. Gan, M.F. Zou, Z.J. Lin, et al., Cancer Lett. 526 (2022) 352–362. doi: 10.1016/j.canlet.2021.11.014
Y. Wang, X.Q. Ma, S.S. Yan, S.S. Shen, et al., Cancer Res. 69 (2009) 7302–7310. doi: 10.1158/0008-5472.CAN-09-0462
J. Sang, W. Li, H.J. Diao, R.Z. Fan, et al., Cancer Lett. 509 (2021) 13–25. doi: 10.1016/j.canlet.2021.03.030
K. Gao, K.W. Xu, P.C. Lin, J.X. Zhu, et al., ACS. Catal. 14 (2024) 14233–14241. doi: 10.1021/acscatal.4c05274
R.H. Xie, L.S. Li, X.N. Fan, J.C. Zi, Chin. Chem. Lett. 31 (2020) 431–433. doi: 10.1016/j.cclet.2019.07.068
R.H. Xie, G.Y. Xia, J.X. Zhu, P.C. Lin, et al., Fitoterapia 149 (2021) 104810. doi: 10.1016/j.fitote.2020.104810
X.M. Ma, L.Y. Mo, Z.P. Ren, X.N. Fan, et al., J. Asian. Nat. Prod. Res. 25 (2023) 519–527. doi: 10.1080/10286020.2023.2197224
D. Chen, J. Li, X. Xu, Z. Sun, et al., Chin. Chem. Lett. 35 (2024) 109451. doi: 10.1016/j.cclet.2023.109451
H. Wang, M. Luo, Y. Wang, J. Yin, et al., Chin. Chem. Lett. 36 (2025) 110348. doi: 10.1016/j.cclet.2024.110348
K.A.G. Donald, R.Y. Hampton, I.B. Fritz, Appl. Environ. Microbiol. 63 (1997) 3341–3344. doi: 10.1128/aem.63.9.3341-3344.1997
D.K. Ro, E.M. Paradise, M. Ouellet, K.J. Fisher, et al., Nature 440 (2006) 940–943. doi: 10.1038/nature04640
H. Dong, S. Chen, J.X. Zhu, K. Gao, et al., J. Biotechnol. 307 (2020) 29–34. doi: 10.1016/j.jbiotec.2019.10.019
G. Scalcinati, C. Knuf, S. Partow, Y. Chen, et al., Metab. Eng. 14 (2012) 91–103. doi: 10.1016/j.ymben.2012.01.007
K.M. Murphy, L.T. Ma, Y.Z. Ding, E.A. Schmelz, et al., Front. Plant Sci. 9 (2018) 01542. doi: 10.3389/fpls.2018.01542
Y.J.J. Zhou, W. Gao, Q.X. Rong, G.J. Jin, et al., J. Am. Chem. Soc. 134 (2012) 3234–3241. doi: 10.1021/ja2114486
P. Urban, C. Mignotte, M. Kazmaier, F. Delorme, et al., J. Biol. Chem. 272 (1997) 19176–19186. doi: 10.1074/jbc.272.31.19176
M.L. Diaz-Chavez, J. Moniodis, L.L. Madilao, S. Jancsik, et al., PLoS One 8 (2013) e75053. doi: 10.1371/journal.pone.0075053
M. Hotze, G. Schröder, J. Schröder, FEBS Lett. 374 (1995) 345–350. doi: 10.1016/0014-5793(95)01141-Z
K.W. Henry, J.T. Nickels, T.D. Edlind, Eukaryot. Cell 1 (2002) 1041–1044. doi: 10.1128/EC.1.6.1041-1044.2002
B. Ozaydin, H. Burd, T.S. Lee, J.D. Keasling, Metab. Eng. 15 (2013) 174–183. doi: 10.1016/j.ymben.2012.07.010
Q. Wang, M.L. Hillwig, Y. Wu, R.J. Peters, Plant Physiol. 158 (2012) 1418–1425. doi: 10.1104/pp.111.187518
S. Croker, P. Gaskin, M. Beale, J. Lenton, Phytochemistry 39 (1995) 11–14. doi: 10.1016/0031-9422(94)00912-D
M.K.L.A.B. C, N.C.D. D, M.R. E, S.L.S. E, et al., J. Ethnopharmacol. 264 (2021) 113272. doi: 10.1016/j.jep.2020.113272
M. Grande, J.R. Morán, M.J. Macías, B. Mancheño, Phytochem. Anal. 4 (2010) 19–24. doi: 10.1021/np50044a011
D. Morrone, X.M. Chen, R.M. Coates, R.J. Peters, Biochem. J. 431 (2010) 337–344. doi: 10.1042/BJ20100597
S. Block, C. Baccelli, B. Tinant, L. Van Meervelt, et al., Phytochemistry 65 (2004) 1165–1171. doi: 10.1016/j.phytochem.2004.02.023
R.R. Ravu, M.R. Jacob, C. Jeffries, Y. Tu, et al., J. Nat. Prod. 78 (2015) 2255–2259. doi: 10.1021/acs.jnatprod.5b00470
S.M. Isyaka, M.K. Langat, E. Mas-Claret, B.M. Mbala, et al., Phytochemistry 170 (2020) 112217. doi: 10.1016/j.phytochem.2019.112217
M.H. Li, F. He, Y. Zhou, M.G. Wang, et al., Arch. Pharm. Res. 43 (2020) 982–982. doi: 10.1007/s12272-020-01267-6
C.A. Helliwell, P.M. Chandler, A. Poole, E.S. Dennis, et al., Proc. Natl. Acad. Sci. U. S. A. 98 (2001) 2065–2070. doi: 10.1073/pnas.98.4.2065
S. Mafu, M.R. Jia, J.C. Zi, D. Morrone, et al., Proc. Natl. Acad. Sci. U. S. A. 113 (2016) 2526–2531. doi: 10.1073/pnas.1512096113
X. Kuang, W. Li, Y. Kanno, N. Yamashita, et al., J. Nat. Med. 70 (2016) 120–126. doi: 10.1007/s11418-015-0940-6
Q. Shi, Y.-w. Sun, D. Meng, Bioorg. Med. Chem. Lett. 27 (2017) 266–270. doi: 10.1016/j.bmcl.2016.11.063
C.M. Chen, Q.L. Yan, M.X. Tao, H.Y. Shi, et al., Microb. Pathog. 131 (2019) 218–226. doi: 10.1016/j.micpath.2019.04.002
Q.Y. Zhang, Z.Y. Cai, Z.W. Yu, C. Di, et al., Cardiovasc. Drugs Ther. 39 (2025) 239–257.

Figure 1 Examples of active LRDs respectively from E. fischeriana (shadowed in pink) and other species (shadowed in blue).

Figure 2 Functional characterization of CYP71D450. GC–MS detection of the extracts of the yeast strains respectively expressing CYP71D450/EfTPS43 (A) and CYP71D450/EfTPS1 (B). GC–MS detection of the extracts of the E. coli strain expressing CYP71D450/EfTPS41 (C) and N. benthamiana expressing CYP71D450/EfTPS14 (D). (E) The structures of the products.

Figure 3 Functional characterization of CYP701A148. GC–MS detection of the extracts of the yeast strains respectively expressing CYP701A148/EfTPS43 (A) and CYP701A148/EfTPS1 (B). GC–MS detection of the extracts of the E. coli strains respectively expressing CYP701A148/EfTPS41 (C) and CYP701A148/EfTPS14 (D). (E) The structures of the products.

 扫一扫看文章
扫一扫看文章

扫一扫关注我们
 DownLoad:
DownLoad:
 下载:
下载:
 下载:
下载: