Figure 1.
Investigations on the chemical structure of E. bolteae WAL-16351 CPS.
Immunological evaluation of the autism-related bacterium Enterocloster bolteae capsular polysaccharide driven by chemical synthesis
Lingxin Li , Jing Hu , Chunjun Qin , Juntao Cai , Xiaopeng Zou , Guangzong Tian , Peter H. Seeberger , Jian Yin

Autism spectrum disorder (ASD) is a neurodevelopmental disorder with dramatically increased morbidity over the past two decades [1,2]. The origins of ASD are related to a variety of genetic and environmental factors [3], and gastrointestinal diseases strongly correlate with ASD [4]. The gut bacterial communities of individuals with ASD were found to differ from those of healthy people [5]. Enterocloster bolteae, formerly known as Clostridium bolteae [6], was found in fecal samples of children with ASD with significantly higher abundance [7]. Oral vancomycin treatments against E. bolteae infections of ASD patients can lead to significant improvements in both ASD symptoms and related gastrointestinal problems [8]. Since E. bolteae has been reported to be resistant to the most common antibiotics [9], an alternative for long-term antibiotic treatment is highly desirable for preventing the associated gastrointestinal diseases. An E. bolteae vaccine might be beneficial for understanding and treating ASD. Unique bacterial surface glycans, such as capsular polysaccharides (CPS), lipopolysaccharides (LPS), and other exopolysaccharides have been extensively employed in the development of carbohydrate-based antibacterial vaccines [10]. The marketed CPS-based vaccines against Haemophilus influenzae type b [11,12], Streptococcus pneumoniae [13], and Neisseria meningitidis [14] have significantly decreased the incidence of associated diseases [15,16]. Monteiro et al. had demonstrated the immunogenic activity of CPS from E. bolteae strain WAL-16351 in rabbits [17]. Nevertheless, the immunogenic epitopes of this carbohydrate antigen, which is important for the design of an effective vaccine, remain to be defined in detail. Synthetic carbohydrate chemistry is an effective approach for preparing structurally well-defined, pure oligosaccharides [18-23], that has greatly helped in identifying the key epitopes of glycan antigens [24-28]. Therefore, it is attractive to perform immunological evaluation of chemically synthetic oligosaccharides related to E. bolteae WAL-16351 CPS.
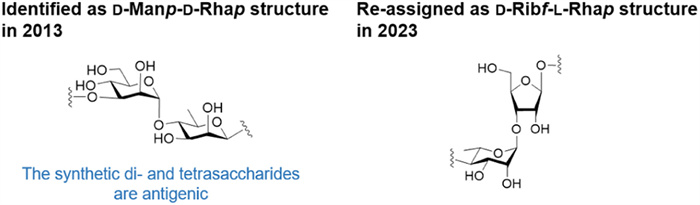
Notably, the structure elucidation of E. bolteae WAL-16351 CPS has been a challenging process (Fig. 1). Recently, the repeating unit of E. bolteae WAL-16351 CPS was re-assigned as [→3)-β-d-Ribf-(1→4)-α-l-Rhap-(1→] (D-Ribf-l-Rhap type structure), and the chemical synthesis of oligosaccharides 1–3 were also completed (Fig. 2a) [29]. The structural revision of E. bolteae WAL-16351 CPS prompted us to investigate the immunological activities concerning the d-Ribf-l-Rhap type structure-related oligosaccharides.
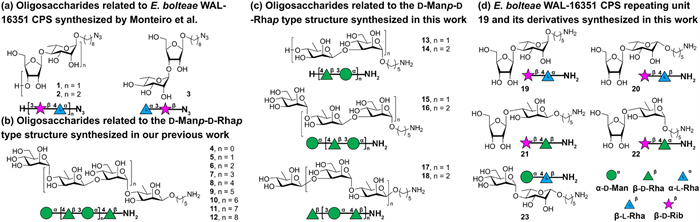
The [→3)-α-d-Manp-(1→4)-β-d-Rhap-(1→] (d-Manp-d-Rhap type structure) was designated as a disaccharide repeating unit of E. bolteae WAL-16351 CPS components, which is on average contained nine times in the polymer [17,30,31]. We synthesized a series of α-d-Manp-β-d-Rhap dimers as long as an 18-mer based on a stereoselective β-d-rhamnosylation and a modular assembly strategy (Fig. 2b). IgG antibodies in rabbit sera inoculated with E. bolteae bacteria can specifically bind to the synthetic di- (4) and tetrasaccharide (5) containing α-d-Manp-β-d-Rhap [32]. Besides, the gas chromatography-mass spectrometry (GC–MS) results confirmed the presence of mannose and galactose in native glycans of E. bolteae WAL-16351 cell wall in addition to ribose and rhamnose. It is suggested that the d-Manp-d-Rhap type structure may be included in other components of E. bolteae, such as glycolipids [29]. Therefore, further investigation is still needed to assess the immunogenicity of the d-Manp-d-Rhap type structure.
The immunological activity of oligosaccharides may be affected by the chain length and frameshifts of the repeating units [33,34]. Therefore, oligosaccharides 13–18 (Fig. 2c) related to the d-Manp-d-Rhap type structure with different lengths and frameshifts were synthesized. To investigate the structure-activity relationships of E. bolteae CPS, disaccharide 19 β-d-Ribf-(1→4)-α-l-Rhap-(1→O(CH2)5NH2 and a series of structurally similar disaccharides including 20 containing a β-l-rhamnoside, 21 containing a β-d-rhamnoside, 22 containing an α-d-rhamnoside, 23 containing an α-d-mannoside were synthesized (Fig. 2d). These synthetic oligosaccharides formed the basis for immunological investigations of this bacterial glycan by performing glycan microarray analyses, animal immunization, and IgG-bacteria binding test.
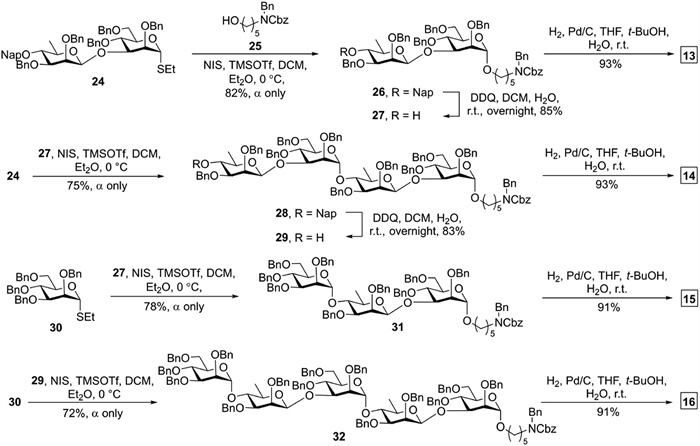
Oligosaccharides 13–16 were assembled from building blocks 24 and 30 (Scheme 1) [32]. Disaccharide 24 with a 1,2-cis-β-glycosidic linkage was efficiently synthesized employing the Crich β-mannosylation approach using 4,6-O-benzylidenated thioglycoside/1-benzenesulfinyl piperidine (BSP)/trifluoromethanesulfonic anhydride (Tf2O) [35,36]. To introduce an aminopentyl linker, donor 24 was glycosylated with linker 25 in dichloromethane (DCM) and Et2O (1:1 v/v) to provide exclusively α-linked disaccharide 26 in 82% yield. Cleavage of the O4 Nap in 26 using 2,3-dichloro-5,6-dicyano-1,4-benzoquinone (DDQ) produced disaccharide acceptor 27 in 85% yield. Disaccharide 24 was coupled with 27 upon N-iodosuccinimide (NIS) and trimethylsilyl trifluoromethanesulfonate (TMSOTf) activation in DCM and Et2O (1:1, v/v) to provide 28 in 75% yield. Removal of O4 Nap furnished tetrasaccharide acceptor 29 in 83% yield. The union of thioglycoside donor 30 and acceptor 27 activated by NIS and TMSOTf led to an α-selective synthesis of trisaccharide 31 in 78% yield. NIS/TMSOTf catalyzed glycosylation of donor 30 with 29 in DCM and Et2O afforded 32 in 72% yield and high α-stereoselectivity. Finally, global deprotection of oligosaccharides 26, 28, 31, 32 by palladium on carbon (Pd/C) hydrogenolysis gave target disaccharide 13 (93%), tetrasaccharide 14 (93%), trisaccharide 15 (91%), pentasaccharide 16 (91%). Target trisaccharide 17 and pentasaccharide 18 were obtained by Pd/C hydrogenolysis of the known compounds (Scheme S1 in Supporting information) [32].
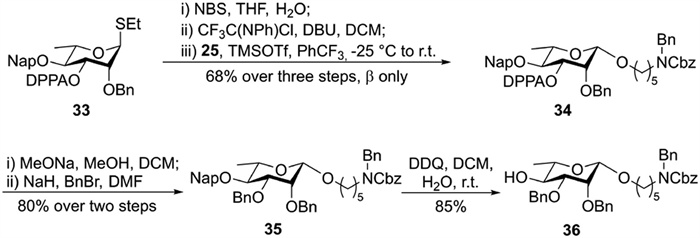
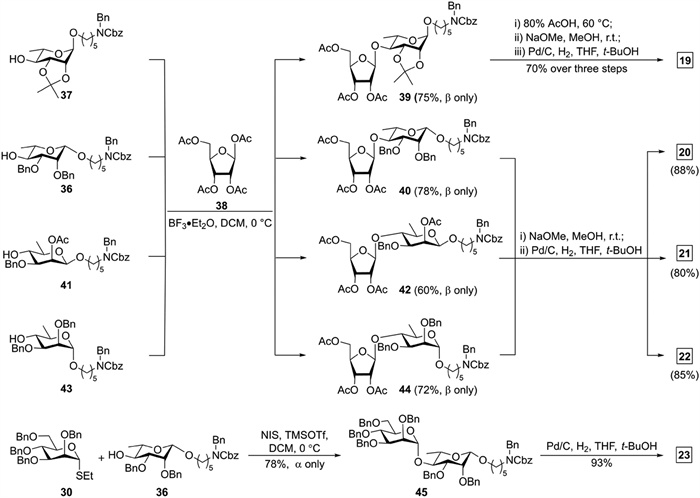
Construction of the β-rhamnosidic linkage commenced with the hydrolysis of l-thiorhamnoside 33 (Scheme S2 in Supporting information) bearing an O3 2-(diphenylphosphinoyl)acetyl (DPPA) group that can promote the formation of the 1,2-cis-β-glycosidic bond [37]. The hydrolyzed product was converted to a trifluoroacetimidate donor [38], which was glycosylated with linker 25 under TMSOTf catalytic conditions to afford 34 with complete β-selectivity in 68% overall yield (Scheme 2). Cleavage of the DPPA group and subsequent benzylation at the O3 afforded 35 in 80% overall yield. Removal of the Nap group in 35 produced glycosyl acceptor 36 in 85% yield.
The union of tetraacetylriboside 38 and acceptor 37 (Scheme S3 in Supporting information) promoted by boron trifluoride diethyl etherate (BF3·Et2O) at 0 ℃ generated disaccharide 39 in 75% yield (Scheme 3). Acceptors 36, 41 [32] and 43 (Scheme S4 in Supporting information) were smoothly glycosylated with 38 to furnish disaccharides 40, 42 and 44, respectively. Small 3JH1, H2 (0–2 Hz) and chemical shifts of C1 (105–110 ppm) of ribose residues revealed the β-stereoselectivity of the four ribosylation reactions [39]. Coupling of thiomannoside 30 and acceptor 36 in the presence of NIS and TMSOTf gave desired α-linked disaccharide 45 (1JC1, H1 = 168.2 Hz) in 78% yield. Global deprotection of 39 commenced with removal of the isopropylidene group in 80% yield. Alkaline hydrolysis and subsequent Pd/C hydrogenolysis furnished the target disaccharides 19 (70%), 20 (88%), 21 (80%), and 22 (85%). Global deprotection of 45 by hydrogenolysis afforded target disaccharide 23 in 93% yield.
A comparison of 1H nuclear magnetic resonance (NMR) and 13C NMR data of isolated E. bolteae 16351 CPS and the synthetic disaccharide units was performed. For the 13C NMR data of the native CPS and disaccharide 4 related to the d-Manp-d-Rhap type structure, the most significant difference arose from the chemical shifts of the anomeric carbons, which was previously thought to be caused by the installation of linker [32]. Notably, the 1H NMR and 13C NMR data of 19 bearing the linker are identical with those of the natural isolate (Fig. S1 and Table S1 in Supporting information).
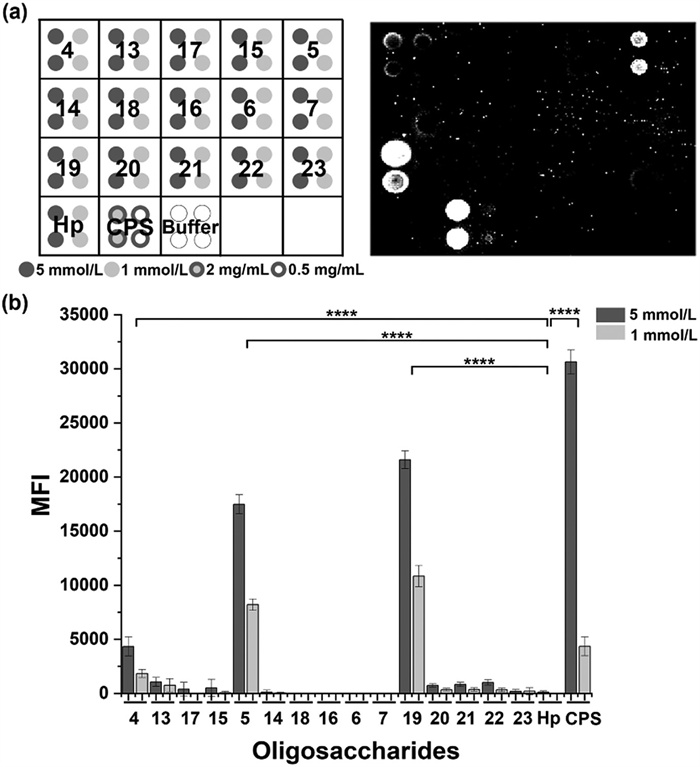
Synthetic oligosaccharides 4–7 [32] and 13–23 were printed on microarray slides to assess the antigenicity of the synthetic oligosaccharides. The glycan arrays were screened with the antisera [32] from rabbits immunized with inactivated E. bolteae WAL-16351. All the animal experiments were approved by the Experimental Animal Ethics Committee of Jiangnan University (Animal Ethics Committee Number: JN. No. 20230615b0300930[317]). The isolated E. bolteae CPS and a synthetic trisaccharide Hp (α-(1→6)-glucan) of Helicobacter pylori serotype O1 core oligosaccharide [40] were used as a positive and a negative control, respectively. Disaccharide β-d-Ribf-α-l-Rhap 19 is well recognized by IgG antibodies in the rabbit sera (Fig. 3). However, the other two β-l-Rha-containing disaccharides 20 and 23, as well as the d-Rha-containing disaccharides 21 and 22 showed almost no binding to the IgG antibodies. It is apparent that the α configuration of the l-rhamnosidic bond is vital for the binding with the anti-E. bolteae antibodies. The strong and highly specific binding between disaccharide 19 and IgG antibodies in the antisera showed great potential of this disaccharide in further immunological investigation.
Both disaccharide 4 and tetrasaccharide 5 can be also detected by IgG antibodies in the rabbit sera, which is consistent with previous results. However, the other oligosaccharides related to the d-Manp-d-Rhap type structure including 6, 7 and 13–18 were not recognized by the IgG antibodies. Different chain lengths and frameshifts impact the antigenicity of the d-Manp-d-Rhap type structure. Therefore, three synthetic oligosaccharides 4, 5, and 19 were further studied with an immunogenicity evaluation to identify promising antigens of E. bolteae WAL-16351.
Conjugation of a carbohydrate hapten to a carrier protein can elicit a T cell-dependent immune response. Three glycoconjugates 4-CRM197, 5-CRM197 and 19-CRM197 were obtained by conjugation of synthetic oligosaccharides 4, 5, and 19 with nontoxic diphtheria toxin mutant CRM197 for further immunological evaluation (Fig. S2a in Supporting information). Conjugation and relative purity of the glycoconjugates were confirmed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). Matrix-assisted laser desorption ionization-time of flight (MALDI-TOF) mass spectrometry analysis was utilized to determine the molar ratio of oligosaccharide to CRM197 (Figs. S2b and c in Supporting information). The average number of oligosaccharide fragments carried by each protein were calculated to be 6, 4 and 7 for glycoconjugates 4-CRM197, 5-CRM197 and 19-CRM197, respectively.
To test the immunogenicity of three glycoconjugates, four groups of BALB/c mice were immunized with phosphate-buffered saline (PBS) (control group), 4-CRM197, 5-CRM197 and 19-CRM197, respectively (Fig. 4a). Each mouse received one priming dose with conjugates containing 5 µg of glycan, formulated with complete Freund's adjuvant (CFA), and two boosting doses of the conjugates in incomplete Freund's adjuvant (ICFA). The antibody titers in post-immune sera collected on day 35 were monitored by enzyme-linked immunosorbent assay (ELISA) using E. bolteae WAL-16351 CPS as the coating antigen.
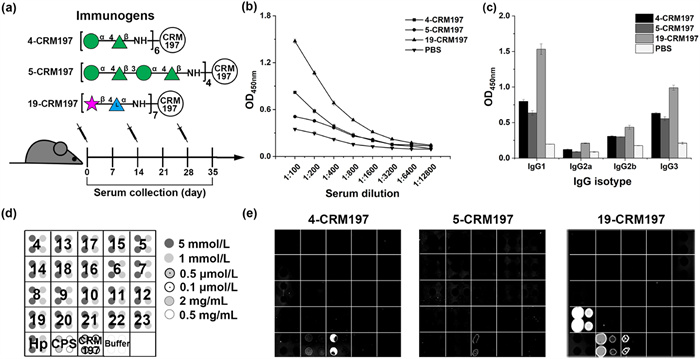
The ELISA data showed that the conjugate 19-CRM197 stimulates high titers of CPS-specific IgG antibodies, whereas mice vaccinated with conjugates 4-CRM197 and 5-CRM197 exhibited a very weak antibody response (Fig. 4b). The IgG isotypes in post-immune sera were further determined (Fig. 4c). IgG1 and IgG3 contributed the bulk of the CPS-specific IgG titer of sera from mice immunized with 19-CRM197, while a weak IgG2a and IgG2b response was observed. In comparison with mice inoculated with 19-CRM197, mice vaccinated with glycoconjugates 4-CRM197 and 5-CRM197 generated lower titers of IgG1 and IgG3 antibodies. The PBS control group was unable to elicit significant levels of CPS-specific IgG isotypes. These results indicated that 19-CRM197 induced a strong immune response in mice and produced high titers of T-cell-dependent CPS-specific antibodies.
The binding capacity of synthetic oligosaccharides with elicited IgG antibodies was evaluated with post-immune sera using glycan arrays (Fig. 4d). The microarrays also included CRM197 and an H. pylori serotype O1 O-antigen trisaccharide (Hp) to assess antibody responses to the carrier protein and an unrelated synthetic glycan. The antibodies generated against glycoconjugate 19-CRM197 were able to recognize disaccharide 19, carrier protein CRM197 and the isolated E. bolteae WAL-16351 CPS. Despite the minor structural changes between all printed oligosaccharides, there was almost no cross-reaction of the post-immune sera with other oligosaccharides (Fig. 4e and Fig. S3 in Supporting information). This result showed that disaccharide 19 can induce a robust and E. bolteae CPS-directed immune response. Moreover, the glycan microarrays results demonstrated that the anti-4-CRM197 and anti-5-CRM197 IgG antibodies cannot recognize any printed synthetic oligosaccharides, confirming that disaccharide 4 and tetrasaccharide 5 are weakly immunogenic.
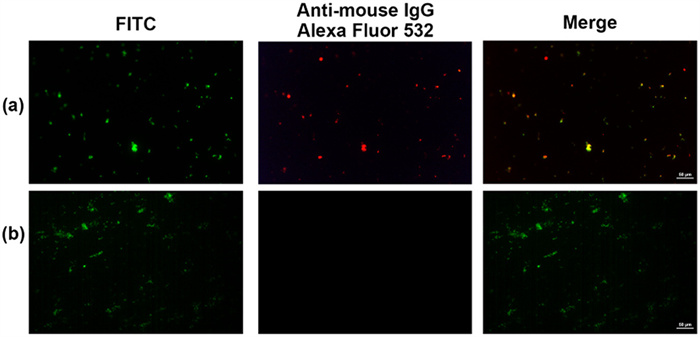
The capacity of antisera IgG antibodies to recognize E. bolteae WAL-16351 bacteria was investigated using immunofluorescence microscopy while H. pylori O6 bacteria served as control. All bacteria were labeled on the surface with fluorescein isothiocyanate (FITC) and can be detected with green fluorescence. The IgG antibodies bound to bacteria were observed with red fluorescent goat anti-mouse IgG-Alexa Fluor 532. The anti-19-CRM197 IgG antibodies significantly bound to the surface of inactivated E. bolteae WAL-16351 bacteria, whereas they showed no binding affinity towards H. pylori O6 bacteria, indicating the binding specificity of IgG antibodies (Fig. 5). In contrast, the anti-4-CRM197 and anti-5-CRM197 IgG antibodies bound weakly to both bacteria (Fig. S4 in Supporting information). Disaccharide 4 and tetrasaccharide 5 are recognized by whole-cell antisera but exhibit poor immunogenicity against E. bolteae WAL-16351. Disaccharide 19 of β-d-Ribf-α-l-Rhap is strongly antigenic and immunogenic, and an immunogenic epitope lead for E. bolteae WAL-16351 vaccine development.
In summary, the repeating unit disaccharide 19 and four derivatives 20–23 were synthesized for the investigation of structure-activity relationships of E. bolteae WAL-16351 CPS. Based on the former antigenicity evaluation of E. bolteae related oligosaccharides, oligosaccharides 13–18 related to the d-Manp-d-Rhap type structure were also chemically prepared. A 1,2-cis-β-glycosylation was efficiently achieved by hydrogen-bond mediated TMSOTf-catalyzed glycosylation. The Crich β-mannosylation approach relying on 4,6-O-benzylidene-induced conformational locking is essential for the formation of β-d-rhamnoside. Glycan microarray analysis showed that disaccharide 19 is antigenic and the configuration plays a role in the recognition of anti-E. bolteae antibodies and carbohydrate antigens. Among the oligosaccharides related to the d-Manp-d-Rhap type structure, only disaccharide 4 and tetrasaccharide 5 are recognized by the anti-E. bolteae IgG antibodies. Glycoconjugate 19-CRM197 elicited a strong T-cell-dependent immune response in mice that can specifically detect the CPS and bacteria. However, 4-CRM197 and 5-CRM197 induced a low level of IgG antibodies that can barely bind to the CPS and bacteria. These results demonstrated that disaccharide 19 related to the CPS repeating unit is a key motif for designing glycoconjugate vaccines against E. bolteae WAL-16351.
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Lingxin Li: Writing – original draft, Methodology, Investigation, Formal analysis, Data curation. Jing Hu: Writing – review & editing, Supervision, Methodology, Investigation, Funding acquisition, Formal analysis, Conceptualization. Chunjun Qin: Writing – review & editing, Methodology, Investigation, Formal analysis. Juntao Cai: Investigation, Formal analysis. Xiaopeng Zou: Investigation. Guangzong Tian: Investigation, Funding acquisition. Peter H. Seeberger: Writing – review & editing, Formal analysis, Conceptualization. Jian Yin: Writing – review & editing, Supervision, Resources, Project administration, Funding acquisition, Formal analysis, Conceptualization.
The authors are grateful to the National Natural Science Foundation of China (Nos. 22325803, 22277042, 22177041, 22207042, 22107037), the Max Planck Society International Partner Group Program, the China Scholarship Council (CSC), and the Fundamental Research Funds for the Central Universities for funding. P.H.S. thanks the Max-Planck Society for generous financial support.
Supplementary material associated with this article can be found, in the online version, at doi:
G. Sharon, N.J. Cruz, D.W. Kang, et al., Cell 177 (2019) 1600–1618. doi: 10.1016/j.cell.2019.05.004
M.J. Maenner, Z. Warren, A.R. Williams, et al., MMWR Surveill Summ. 72 (2023) 1–14. doi: 10.15585/mmwr.ss7202a1
T. Hirota, B.H. King, JAMA 329 (2023) 157–168. doi: 10.1001/jama.2022.23661
F. Strati, D. Cavalieri, D. Albanese, et al., Microbiome 5 (2017) 24–34. doi: 10.1186/s40168-017-0242-1
Y. Wan, T. Zuo, Z. Xu, et al., Gut 71 (2022) 910–918. doi: 10.1136/gutjnl-2020-324015
K.N. Haas, J.L. Blanchard, Int. J. Syst. Evol. Microbiol. 70 (2020) 23–34. doi: 10.1099/ijsem.0.003698
Y. Song, C. Liu, D.R. Molitoris, et al., Syst. Appl. Microbiol. 26 (2003) 84–89. doi: 10.1078/072320203322337353
R.H. Sandler, S.M. Finegold, E.R. Bolte, et al., J. Child Neurol. 15 (2000) 429–435. doi: 10.1177/088307380001500701
Y.A. Warren, K.L. Tyrrell, D.M. Citron, et al., J. Clin. Microbiol. 44 (2006) 2416–2422. doi: 10.1128/JCM.00116-06
R. Rohokale, Z. Guo, ACS Infect. Dis. 9 (2023) 178–212. doi: 10.1021/acsinfecdis.2c00559
F. Boisnard, C. Manson, L. Serradell, et al., Expert Rev. Vaccines 22 (2023) 1196–1213. doi: 10.1080/14760584.2023.2280236
C. Qin, J. Hu, W. Tong, et al., Chin. J. Nat. Med. 20 (2022) 633–640.
M. Shirley, Drugs 82 (2022) 989–999. doi: 10.1007/s40265-022-01733-z
S.R. Parikh, H. Campbell, J.A. Bettinger, et al., J. Infect. 81 (2020) 483–498. doi: 10.1016/j.jinf.2020.05.079
F. Khatun, I. Toth, R.J. Stephenson, Adv. Drug. Deliv. Rev. 165 (2020) 117–126.
M. Vella, D. Pace, Expert Opin. Biol. Ther. 15 (2015) 529–546. doi: 10.1517/14712598.2015.993375
B. Pequegnat, M. Sagermann, M. Valliani, et al., Vaccine 31 (2013) 2787–2790. doi: 10.1016/j.vaccine.2013.04.018
P.H. Seeberger, Chem. Rev. 121 (2021) 3598–3626. doi: 10.1021/acs.chemrev.0c01210
G. Tian, J. Hu, C. Qin, et al., J. Am. Chem. Soc. 146 (2024) 18427–18439. doi: 10.1021/jacs.4c03814
C. Qin, M. Ding, G. Tian, et al., Chin. J. Nat. Med. 20 (2022) 401–420.
C. Qin, B. Schumann, X. Zou, et al., J. Am. Chem. Soc. 140 (2018) 3120–3127. doi: 10.1021/jacs.8b00148
C. Qin, H. Hou, M. Ding, et al., Chin. J. Nat. Med. 20 (2022) 387–392.
Z. Hou, J. Wang, X. Zhang, et al., Chin. Chem. Lett. 34 (2023) 107804. doi: 10.1016/j.cclet.2022.107804
G. Tian, J. Hu, C. Qin, et al., Angew. Chem. Int. Ed. 59 (2020) 13362–13370. doi: 10.1002/anie.202004267
X. Zhou, L. Li, Z. Zhang, et al., Angew. Chem. Int. Ed. 61 (2022) e202204420. doi: 10.1002/anie.202204420
P. Shen, H. Lin, Y. Bao, et al., Chin. Chem. Lett. 34 (2023) 107679. doi: 10.1016/j.cclet.2022.07.022
J. Sianturi, P. Priegue, J. Hu, et al., Angew. Chem. Int. Ed. 61 (2022) e202209556. doi: 10.1002/anie.202209556
C. Qin, L. Li, G. Tian, et al., J. Am. Chem. Soc. 144 (2022) 21068–21079. doi: 10.1021/jacs.2c05953
N.W. Frame, M.J. Allas, B. Pequegnat, et al., Carbohydr. Res. 526 (2023) 108805. doi: 10.1016/j.carres.2023.108805
B. Pequegnat, M.A. Monteiro, Curr. Med. Chem. 26 (2019) 6341–6348. doi: 10.2174/0929867326666190225164527
J. Davidson, C. Gauthier-Signore, K.P. Bishop, et al., Org. Biomol. Chem. 20 (2022) 2964–2980. doi: 10.1039/d2ob00131d
J. Cai, J. Hu, C. Qin, et al., Angew. Chem. Int. Ed. 59 (2020) 20529–20537. doi: 10.1002/anie.202007209
M. Zhao, G. Tian, C. Qin, et al., Chin. J. Chem. 42 (2024) 243–251. doi: 10.1002/cjoc.202300510
B. Schumann, H.S. Hahm, S.G. Parameswarappa, et al., Sci. Transl. Med. 9 (2017) eaaf5347. doi: 10.1126/scitranslmed.aaf5347
D. Crich, Q. Yao, J. Am. Chem. Soc. 126 (2004) 8232–8236. doi: 10.1021/ja048070j
J. Cai, X. Yuan, Y. Kong, et al., Chin. J. Nat. Med. 21 (2023) 886–901.
X. Liu, Y. Lin, W. Peng, et al., Angew. Chem. Int. Ed. 61 (2022) e202206128. doi: 10.1002/anie.202206128
B. Yu, H. Tao, Tetrahedron Lett. 42 (2001) 2405–2407. doi: 10.1016/S0040-4039(01)00157-5
A. Imamura, T. Lowary, Trends Glycosci. Glycotechnol. 23 (2011) 134–152. doi: 10.4052/tigg.23.134
X. Zou, J. Hu, M. Zhao, et al., J. Am. Chem. Soc. 144 (2022) 14535–14547. doi: 10.1021/jacs.2c03068

Figure 1 Synthesis of oligosaccharides 13–16 related to the d-Manp-d-Rhap type structure.

Figure 3 Antigenicity evaluation of synthetic glycan fragments using a glycan microarray. (a) The printing pattern and representative scan of glycan microarray with inactivated E. bolteae bacteria immunized rabbit sera (day 35, 1:100 dilution). Buffer: sodium phosphate (50 mmol/L, pH 8.5). (b) Quantification of the mean fluorescence intensity (MFI) values of each oligosaccharide fragment. Data are individual values from n = 3 rabbits. The data of oligosaccharide groups with 5 mmol/L concentration were analyzed by one-way ANOVA with Dunnett's multiple comparisons. Hp served as a control group. ****P < 0.0001. Error bars represent the mean ± SD of six spots of three separate arrays.

Figure 4 Evaluation of glycoconjugates 4-CRM197, 5-CRM197, and 19-CRM197 in mice. (a) Schematic immunization schedule. The CPS-specific (b) IgG titer and (c) IgG isotypes (1:50 dilution) of D35 sera analyzed by ELISA. Data from triplicate determinations were plotted as mean ± SD. (d) Printing pattern of glycan microarrays. Buffer: 50 mmol/L of sodium phosphate, pH 8.5. (e) Representative scan of glycan microarray with sera from mice immunized by 4-CRM197, 5-CRM197 and 19-CRM197 (day 35, 1:50 dilution).

 扫一扫看文章
扫一扫看文章

扫一扫关注我们
 DownLoad:
DownLoad:

 下载:
下载:




 下载:
下载: