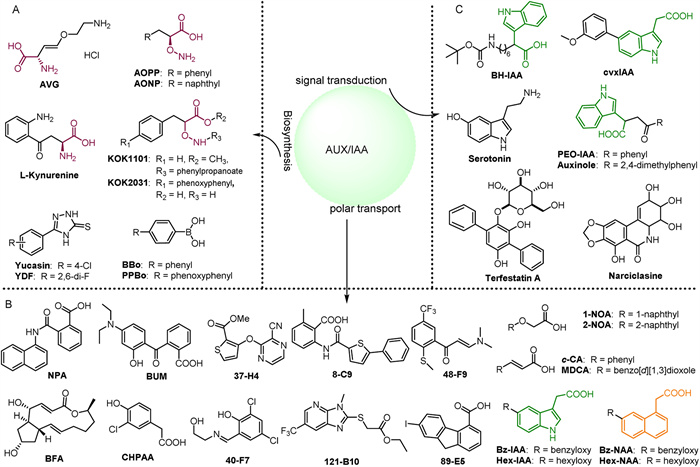
Figure 1.
(A) Small molecules that regulate auxin biosynthesis. (B) Small molecules regulating the polar transport of auxin. (C) Small molecules that regulate auxin signal transduction.
Small molecule chemical scaffolds in plant growth regulators for the development of agrochemicals
Ali Dai , Zhiguo Zheng , Liusheng Duan , Jian Wu , Weiming Tan

Plants are dynamic organisms constantly influenced by intricate environments and integrate cellular signaling hubs. Agrochemicals play a vital role in regulating plant growth and function, thereby enhancing crop yield and quality [1,2]. Plant growth regulators (PGRs) which can be either natural or synthetic compounds, are essential for controlling the development and metabolic processes of higher plants. Plant growth inhibitors, a key subset of PGRs, significantly influence plant growth and development by inhibiting hormone synthesis, or blocking receptors [3,4]. PGRs govern all aspects of plant development and growth, allowing for the manipulation of hormone balance to influence crop growth [5,6]. By strategically modulating endogenous hormone signals, PGRs serve as valuable tools in agriculture, enabling precise control over plant growth and development.
Various natural PGRs with distinct structures have been identified, each playing a crucial role in regulating plant growth, development, and defense. The first PGRs to be identified were auxins (IAA), followed by cytokinins (CTK), abscisic acid (ABA), ethylene (ETH), gibberellins (GA), brassinosteroids (BR), jasmonates (JA), and more recently, strigolactones (SL). These PGRs, whether acting individually or in combination with other hormones, are essential for various physiological processes [7]. The diverse chemical structures of plant hormones underpin their physiological functions, with each structure conferring specific biological activities. For instance, the indole ring serves as the core structure of auxin, facilitating its binding to plant cell receptors and initiating physiological responses [8]. Additionally, hormones like GA, ABA, and CTK feature terpene or adenine ring structures that enable them to interact with specific cellular receptors and initiate signal transduction pathways [8,9]. SL consists of one tricyclic lactone and one monocyclic lactone connected by enol ether bonds [10]. JA, with a cyclopentanone ring and a carboxylic acid group, and the steroidal compound BR contribute to the rich chemical diversity crucial for agricultural research [8]. Beyond their roles as PGRs, compounds with these structures may also function as insecticides or fungicides. Examples include carbendazim, azadirachtin, matrine, artemisinin, and 20-hydroxyecdysone, all of which are widely utilized in agriculture [11,12].
However, the practical application of natural PGRs is limited due to difficulties in extracting them from plants and the complexity of their synthesis methods, making them insufficient for modern agriculture needs. Developing new PGRs through modification and/or design synthesis, guided by key enzymes, target proteins, and maternal structures, has the potential to replace natural PGRs [13]. Meanwhile, small molecules can be employed conditionally, in a dose-dependent manner, and reversibly, providing advantages in bypassing the constraints of lethality and functional redundancy associated with conventional mutation screening [14]. Small molecules are useful in plant research because they allow for the investigation of gene functions without encountering issues of gene redundancy. For instance, the unique characteristics of the ABA receptor can be attributed to its agonist, pyrabactin, which selectively binds to a certain subset of receptors out of a total of 14 homologues [15,16]. By synthesizing small molecules that mimic the actions of endogenous hormones, researchers can elucidate many of their functional characteristics. By tailoring small molecules to target specific functions or receptors, scientists can modify hormone chemistry and gain valuable insights into protein binding pockets. This can lead to the development of a wide range of potent derivatives [17].
This paper explores research on the unique chemical structures of various plant hormones. It aims to deepen our understanding of how these hormones regulate plant growth and development, including their roles in anabolic processes and signaling pathways. The paper also examines the biological properties of structure/activity analogs and the relationship between structure and function. These insights provide a strong scientific foundation for studying PGRs and employing small molecules to enhance agricultural practices.
Auxin, a class of compounds including indole-3-acetic acid (IAA), has been studied since Darwin first observed its effects on plant growth and development in 1880 [18]. Auxin physiologically influences various processes in plants, such as cell differentiation, elongation, lateral root formation, growth, and development. It also plays a role in regulating apical dominance, senescence, and responses to abiotic stresses [19,20]. In recent years, significant research has focused on the distribution and influencing factors of auxin in plants, as well as its biological functions and applications in agriculture. Small molecules serve as chemical probes for studying tissue and organ functions without disrupting overall development, facilitating the swift and effective elucidation of the molecular pathways regulating auxin responses. These small molecules, whether mimicking auxin or modulating auxin-related processes, including agonists, antagonists, synthesis inhibitors, and transport inhibitors, are extensively employed in scientific investigations.
Several small molecules play significant roles in auxin biosynthesis, as depicted in Fig. 1A. For instance, both AVG and AOPP share the common characteristic of impeding the function of pyridoxal enzymes and/or aminotransferases [21]. AOPP specifically inhibits various growth processes in Arabidopsis thaliana (A. thaliana), such as root elongation, gravitropism, root tilt, and root hair formation. Treatment of A. thaliana seedlings with the derivative AONP results in an auxin-deficient phenotype, which can be reversed by exogenous IAA [21,22]. l-Kynurenine (L-kyn) exhibits greater efficacy than AVG as an alternative substrate, competitively inhibiting TAA/TAR aminotransferase activity and suppressing auxin production. Its binding site closely resembles that of the substrate (I-Trp), forming a hydrogen bond with Arg117 to secure the L-kyn molecule in the pocket. Application of L-kyn reduces ethylene-induced auxin synthesis in the root system [23]. Additionally, Yucasin (5-(4-chlorophenyl)-4H-1, 2, 4-triazole-3-thiol) and its derivatives YDF, compounds BBo, and PPBo (with Ki values of 67 and 56 nmol/L, respectively) inhibit the YUC synthase, leading to a decrease in IAA content and subsequent inhibition of primary root elongation and lateral root formation [24-26]. Compounds KOK1101 and KOK2031 decrease IAA levels and plant dry weight in tomatoes and rice, respectively. The Ki value of KOK2031 is 276 nmol/L, and its inhibitory effect can be reversed by exogenous IAA [27,28].
The polar transport of auxin is influenced by numerous small molecules (Fig. 1C). One significant inhibitor of auxin transport is N-1-naphthylphthalamic acid (NPA), which impacts processes such as lateral root development, apical dominance, phyllotaxis, shoot apical meristem integrity, tillering, and adventitious root formation in monocots [29]. Auxin transport in plants is regulated by various inhibitors and facilitators. BUM, similar to NPA, has a lower IC50 value, while BFA affects auxin efflux by impacting efflux carriers on the plasma membrane [30,31]. ATIs like 37-H4 and 48-F9 inhibit IAA transport, leading to its accumulation, with 37-H4 showing similar efficacy to NPA. Other compounds like 13-F11 and 54-E11 have minimal effects on IAA transport. 8-C9 acts like BFA, causing internalization of PIN1 and PIN2 proteins, while 40-F7, 89-E5, and 121-B10 influence the cellular trafficking of IAA transporters [32]. On the other hand, auxin transport is facilitated by plasma membrane-localized PIN, ABCB, and AUX1/LAX transporters. Auxin transport is facilitated by plasma membrane-localized PIN, ABCB, and AUX1/LAX transporters. Alkoxyauxins, such as hexyloxy (Hex-) and benzyloxy (Bz-), are potent modulators of asymmetric auxin distribution in plants. For instance, Hex-IAA and Bz-IAA can block the functions of PIN, ABCB, and AUX1, whereas Hex-NAA and Bz-NAA specifically affect PINs and ABCBs [33,34]. Compounds like 1-NOA can impede both auxin influx and efflux carriers, whereas 2-NOA and CHPAA primarily hinder auxin influx [35]. Introducing the auxin efflux inhibitor c-CA has been shown to promote cell division and expansion in A. thaliana leaves [36,37]. Additionally, its derivative MDCA not only suppresses primary root growth but also promotes the formation of lateral and adventitious roots [38].
Auxin antagonists, also known as inhibitors of auxin signaling, such as BH-IAA, specifically target TIR1/AFB (Fig. 1B). The IAA component of BH-IAA shares a similar conformation to IAA, with its long alkyl chain directed towards the Aux/IAA binding site and its benzene ring forming strong interactions with Phe82 of TIR1. This critical residue for Aux/IAA recognition allows BH-IAA to competitively inhibit various auxin responses in plants [39,40]. PEO-IAA and auxinole, developed using the crystal structure of the TIR1-BH-IAA complex, are potent antiauxins with comparable specificity but greater affinity than BH-IAA [41]. The inhibitors terfestatinA (TrfA) and narciclasine (NCS) regulate the degradation of Aux/IAA proteins. TrfA affects root auxin responses such as main root inhibition, lateral root initiation, root hair promotion, and root gravitropism [42,43]. Serotonin, structurally similar to IAA, acts as a natural antagonist by competitively inhibiting the expression of genes induced by auxin. Conversely, the compound cvx-IAA can block downstream auxin signaling [44,45].
The leucine-rich repeat domain of TIR1 can detect both auxin and Aux/IAA peptide substrates with a single surface pocket. Auxin is specifically recognized at the base of this pocket, interacting with a partially flexible site capable of accommodating various auxin analogues (Fig. S1A in Supporting information). For further details, please refer to Supporting information. In addition, there are small molecules synthesized based on the indole structure, as depicted in Fig. S1B (Supporting information), including several compounds with distinct functions. Compound 1 serves as a key biosynthetic precursor. Compound 2 disrupts AUX1, leading to IAA deficiency, while compound 3 stimulates seed germination and impedes radicle growth post-germination [46]. Compounds 4a and 4b significantly inhibit Brassica napus roots, with inhibition rates of 96% and 95% at 100 mg/L, and 92% and 93% at a tenfold lower concentration, respectively [46,47]. Chlorinated indole-3-tetrazoles (5) exhibit selectivity in binding to TIR1 and enhance in vivo efficacy. Notably, 6-chloroindole-3-tetrazole (5a) has an IC50 value of 19 µmol/L in a primary root growth inhibition assay, compared to the commercial herbicide picloram, which has an IC50 of 5 mmol/L [48].
Cytokinins (CTK) are adenine derivatives with a side chain at the N6 position. Variations in the side chain structure can significantly alter their chemical properties, particularly lipophilicity and hydrophobicity. These molecules play a crucial role in regulating various plant growth and development processes, including cell division, differentiation, stem and root growth, leaf senescence, apical dominance, fruit and seed development, nutritional signaling, and responses to both biotic and abiotic stresses [49].
Decreasing cytokinin levels through cytokinin oxidase (CKX) can benefit plant growth, development, and yield. Urea compounds such as TDZ, APPU, CPPU, and CPBU, exhibit greater resistance to CKX inactivation or breakdown compared to naturally occurring isoprenoid cytokinins [50,51]. TDZ is used as a defoliant in cotton by promoting the natural formation and detachment of the abscission zone between the petiole and the stem. Its derivatives, 2HE-TDZ and 3TFM-TDZ, inhibit CKX activity more effectively than TDZ, with ZmCKX1 and ZmCKX4a in maize showing lower IC50 values. The binding mechanisms of 2HE-TDZ and 3TFM-TDZ to the active site of ZmCKX4a differ [52]. CPPU is employed to increase fruit size in crops like kiwis and grapes. Its derivatives have varying affinities for HK receptors, with CPPU being the most effective, followed by DCPPU, ACPPU, and APPU. However, TDZ remains a more effective ligand than CPPU [50]. INCYDE and F-INCYDE have similar inhibitory IC50 values to TDZ. The presence of electron-withdrawing substituents in these compounds enhances their inhibitory activity against AtCKX2. However, the effectiveness of INCYDE and F-INCYDE is reduced by the formation of a hydrogen bond between the F atom substituent and the electron donor of the cell division protein receptor (Fig. 2A) [53,54].
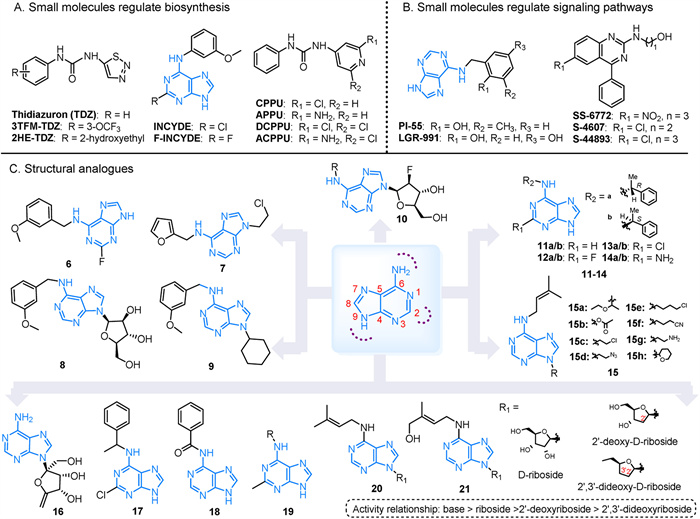
Due to the impact of CTK on root development, new PGRs called anti-cytokinins have emerged, demonstrating a beneficial effect on root growth. One such inhibitor is PI-55, which is structurally similar to 6-benzylaminopurine (BAP). PI-55 hinders the interaction between BAP and specific receptors CRE1/AHK4 in A. thaliana. This interference, along with competitive binding of AHK3 and suppression of the cytokinin response gene ARR5 induction, accelerates seed germination, enhances root growth, and promotes lateral root formation in A. thaliana [55]. Similarly, LGR-991, which has the same potency, blocks the CRE1/AHK4 receptor and acts as a competitive inhibitor of AHK3. This results in quicker seed germination and longer hypocotyl length in seedlings grown in the dark [56]. The CRE1 gene, pivotal in root cell division, has been used to identify compounds that impede receptor activity, such as the phenylquinazoline antagonists SS-6772 and S-4607 [57]. Additionally, the chemically modified S-4893 non-competitively inhibits the binding of 2-isopentenyladenine to CRE1. This inhibition hinders callus formation, suppresses root elongation induced by CTK, and promotes the growth of rice seeds, crowns, and lateral roots [57]. Interestingly, while the competitive inhibitors PI-55 and LGR-991 are purine derivatives, the non-competitive inhibitor S-4893, featuring a phenylquinazoline structure, does not share structural similarity with CTK (Fig. 2B).
Cytokinins play a crucial role in agrochemistry, biotechnology, and pharmaceuticals. Synthetic plant growth regulators are preferred due to their efficacy, cost-effectiveness, and lower toxicity. Developing new synthetic growth regulators by synthesizing various derivatives based on purine scaffolds is a versatile approach. By introducing substitutions at different positions on the adenine moiety, a diverse range of compounds with significant biological properties can be synthesized (Fig. 2C) [58].
The ability of isopentenyladenine (iP) to interact with cytokinin receptors such as A. thaliana CRE1/AHK4, and maize ZmHK1 allows certain synthesized derivatives to induce the transcription of the cytokinin response regulator ARR5 in A. thaliana seedlings. Compounds 6 and 7 have been shown to increase tiller number and grain yield in spring barley and winter wheat while also affecting endogenous CTK content [59]. Compound 8 specifically stimulates growth in spring barley. However, although the antagonist PI-55 increases tiller number and grain yield in winter wheat, it inhibits tiller growth in spring barley [59]. Compound 9 promotes root growth and overall organ development at nanomolar concentrations by increasing root/leaf length and dry root/leaf mass production [60]. Drought and salinity negatively impact seed germination, seedling emergence, and early seedling establishment, reducing crop yields due to their effects on plant metabolism. Nevertheless, compound 10 has shown promise in mitigating these effects in challenging environments [61]. Substituents on AHK3-specific cytokinins 11–14, particularly 11b, 12b, and 13b, include H, F, or Cl substitutions at the C-2 position of the purine heterocycle. Compound 13 has a significant impact on AHK3 [62]. Compounds 15a, 15b, and 15d-15e have demonstrated high efficacy in promoting cytokinin-dependent tobacco callus proliferation, with similar activities to iP in compounds 15c and 15g. Compound 15h can activate the CRE1/AHK4 receptor up to 25% at 50 µmol/L, while 15b, 15d, and 15e, despite their potency, do not activate CRE1/AHK4 [63]. Additionally, while neither 15c nor 15e can activate the ZmHK1 receptor, concentrations above 10 µmol/L can initiate the signaling pathway through the ZmHK3a receptor [63].
Additionally, a novel agrochemical product known as decoyinine (16) was approved in China in 2021 (registration number PD20212929). This compound significantly boosts root and coleoptile growth, tillering, and early maturation. For example, treating corn with compound 16 via seed soaking enhances various growth parameters from the seedling stage to maturity in the field, including plant height, root length, mesocotyl length, and ear weight [64,65]. A high-throughput screening of N-(phenylmethyl)-9H-purin-6-amine (6-BA) derivatives against the complete set of A. thaliana cytokinin receptors (AHK2–4) identified potent compounds with selective affinities for specific receptors. For instance, compound 17 demonstrates a strong binding affinity for the AHK3 receptor, while compound 18 shows a preference for the AHK2 receptor [66]. Additionally, compound 19 exhibits cytokinin activity at concentrations ranging from 10 mol/L–6 to 10–5 mol/L, depending on the nature of the substituent on the N atom of the outer ring [67]. Compounds 20 and 21, which involve adding 2′-deoxy-D-ribose or 2′,3′-dideoxy-D-ribose to the purine N9 position of isoprenyl cytokinin, exhibit the highest activity in their free base form. Compared to free bases, 2′-deoxynucleoside derivatives show no toxicity towards tobacco callus at concentrations up to 10–4 mol/L, with a hierarchy of activity: base > ribose > 2′-deoxyribose > 2′,3′-dideoxyribose [68].
Brassinosteroids (BR) are a class of steroids characterized by a cyclopentanoperhy-drophenanthrene skeleton, which includes three six-membered rings (A, B, and C) and one five-membered ring (D). The structural variations in BRs mainly arise from differences in the A/B carbocyclic rings and the substituents on the alkane side chain [69]. Brassin, initially discovered in the pollen extract of Brassica napus L., is known for its ability to promote plant growth and elongation of soybean stems, leading to its recognition as a brassinosteroid [70]. BRs play a crucial role in regulating various physiological processes in plants, including cell division and elongation in roots and stems, reproductive development, leaf expansion and senescence, seed germination, and pollen tube growth [71].
Triazole compounds inhibit cytochrome P450, a widely distributed plant enzyme essential for oxidation and hydroxylation reactions [72]. These compounds play a crucial role in the biosynthesis and signal transduction pathways of brassinosteroids. Further details can be found in Supporting information and Fig. S2A (Supporting information).
The synthesis of BR is motivated by their possible use as steroidal plant growth regulators in agriculture. Researchers have explored various modified B-ring analogs, including brassinosteroid lactams, thiolactones, cyclic ethers, ketones, hydroxyl groups, and cyclic methylene derivatives. Compounds with polar functional groups generally retain significant biological activity, with those containing 2α/3α-dihydroxy groups showing the highest levels of activity. The presence and configuration of the hydroxyl group on the A ring greatly influence the compound's activity. Loss of the seven-membered B-ring lactone and specific modifications to the B-ring are crucial for optimizing activity levels [73,74]. BRs are derived from the 5-cholestane skeleton, with α-hydroxyl, β-hydroxyl, and ketone groups at the C-3 position serving as precursors for compounds with 2α, 3α-o-hydroxyl groups. The most active BR are characterized by the adjacent 2α and 3α hydroxyl groups on the A ring. Structure-activity relationships indicates that the order of activity is 2α, 3α > 2α, 3β > 2β, 3α > 2β, 3β (Fig. S2B in Supporting information) [90,91]. The α-substituted hydroxyl group on C-2 is particularly important for enhancing biological activity. Derivatives 22 and 24a/24b exhibit significant activity, though not as potent as BR. Compound 23 shows lower activity. B-ring lactam and thiolactam analogs 25a/25b have weak activity against rice plants, while 29a/29b are approximately 5–7 times more potent than BR [75,76]. Compounds 26a, 26b, 26e, 27e, 28a, and 28e promote growth in A. thaliana at low concentrations [77]. Molecular docking studies reveal that compounds 26c, 26d, 26e, 28c, and 28d have similar or better binding energies than BR, suggesting their potential to bind effectively in the BRI1 receptor cavity. Additionally, derivatives 26a, 26b, 26e, and 27e, with shorter side chains, cause slight root elongation at concentrations of 0.1 and 1 nmol/L, while, 28a and 28e, with longer side chains, show a similar trend [77]. Analog 29c induces various growth effects in soybean second internodes, similar to BR. Compounds 25a/25b partially mitigate growth defects induced by Brz2001, with compound 25a being more effective in reversing growth inhibition, possibly due to structural differences [78]. Most compounds stimulate cell division and elongation at low concentrations. Lactones 31a-31d exhibit the highest activity (IC50 range: 2.56 × 10–6 ~ 1.4 × 10–6 mol/L), with medium activity observed in BR ≥ 31a> 32b > 32a in sensitivity tests affecting roots and hypocotyls in A. thaliana (Fig. S3 in Supporting information) [79].
Gibberellin (GA), a tetracyclic diterpenoid compound, was first isolated from rice in 1938 as a metabolite of the pathogenic fungus Gibberella fujikuroi [80]. Active forms of Gibberellin, including GA1, GA3, GA4, and GA7, share several structural features: a 3β-hydroxyl group, a non-hydroxyl group at the C-2 position, a C-6 carboxyl group, and a C-lactone ring. These compounds are crucial for various plant processes, including seed dormancy and germination, stem elongation, leaf expansion and senescence, fruit senescence, sexual expression, enzyme induction, and hormone stabilization [81].
Gibberellin's role in promoting plant hypocotyl elongation, seed germination, and flowering regulation can enhance plant yield but also creates the issue of "high yield and lodging". To address this, synthetic small molecules are used as gibberellin inhibitors (Fig. 3A) [82]. Notably, triazole plant growth regulators like uniconazole (UNI) and paclobutrazol (PAC) inhibit the enzyme ent-kaurene oxidase (KO) in plants, which interferes with the formation of GA precursor and subsequent gibberellin biosynthesis [83]. Both UNI and PAC retard growth, including stem elongation inhibition, internode shortening, enhanced stress tolerance, increased tillering, and stimulation of lateral and flower buds, while also promoting yield. However, UNI is significantly more effective than PAC. UNI adjusts endogenous hormone levels to improve lodging resistance and stress resilience in crops such as petunias, barley, wheat, soybeans, and buckwheat [84,85]. In contrast, PAC increases the activity of indole acetic acid oxidase in rice, which reduces IAA levels, suppresses top growth, facilitates lateral bud growth, and mitigates oxidative stress by enhancing the antioxidant system and reducing lipid peroxidation under adverse environmental conditions [86].
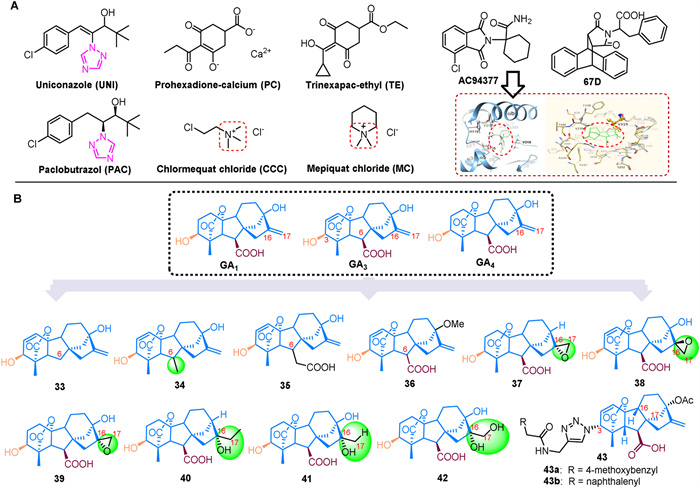
At the same time, GA inhibitors such as chlormequat chloride (CCC) and trinexapac-ethyl (TE) affect gibberellin metabolism in plants, leading to suppressed cell elongation and growth in stems and leaves. This results in shorter stems, which enhances stem strength, helps prevent lodging and increases crop yields [87,88]. The reduced form of TE disrupts the electron transport chain more effectively than the oxidized form, potentially enhancing stress tolerance in treated plants by lowering respiration rates [89]. Prohexadione-calcium (PC) is known for its high efficacy, low toxicity, and lack of residuals. It reduces plant height, increases survival rates, and, when applied before flowering, helps prevent flower withering and bud blight [90]. PC also enhances tillering, photosynthetic efficiency, osmotic regulation, and antioxidant enzyme activity in rice, while reducing membrane lipid peroxidation, thus improving the tillering phase and salt tolerance [91]. Similarly, mepiquat chloride (MC) slows plant vegetative growth by inhibiting gibberellin activity and preventing cell elongation, leading to a reduction in internode length.
The crystal structures of GA3/GA4-AtGID1A-DELLA in A. thaliana and GA3/GA4 bound to OsGID1 in rice have been reported, providing a foundation for discovering new functional analogs of GA based on receptor structures [92]. Early synthesized compounds such as AC-94377, which is similar to GA4, modulate plant growth and exhibit GA activity [93]. The AC94377-AtGID1A-DELLA complex promotes DELLA protein degradation, with the Val-319 residue playing a key role in the interaction between the GID1 protein and the benzene ring of AC94377 [93]. AC-94377 disrupts seed dormancy and enhances germination when applied to the soil surface, as seen in dormant wild mustard seeds [94,95]. It also stimulates stem elongation, flowering, fruiting, and sexual organ development in male fern plants. Acting as a GID1 agonist, AC-94377 has a carboxamide group that is crucial for its binding affinity to various GID1 subtypes [96]. Another GA analog, 67D, functions similarly to AC-94377 as a GID1 agonist and engages in the same signaling pathway as GA [97]. 67D induces DELLA protein RGA degradation, suppresses GA3ox1 expression, and binds to GID1 to counter the non-germinating phenotype of ga1–3 mutant seeds (Fig. 3A) [97,98].
One approach to obtaining active compounds is through structural modification. Biologically active GAs are characterized by a 3β-hydroxyl group, a non-hydroxyl group at the C-2 position, a C-6 carboxyl group, and a C-lactone ring. To study the impact of the C-6 carboxyl group on activity, experiments were conducted with dwarf peas, corn, rice, and barley. Compounds 33–35 were bio-assayed, revealing significantly lower activity compared to GA3. Specifically, compound 34 showed no activity, compound 33 had some activity in dwarf rice, and compound 36 demonstrated activity in dwarf peas, cucumbers, and lettuce [99]. GA1 and GA4 are the primary endogenous bioactive GAs in vascular plants, with their derivatives 37–42 also showing bioactivity in rice seedlings. However, 16, 17-epoxidation of GA1 and GA4 reduces their ability to promote elongation in rice leaf sheaths, indicating that these epoxy groups may cause a deactivation process through 16, 17-epoxidation and subsequent hydrolysis [100]. Additionally, a structural analogue, compound 43, was synthesized by adding an amide group at the 3-OH position, based on GA3. At a concentration of 100 µmol/L, compounds 43a and 43b exhibited inhibitory activities of 48.6% and 41.4% against rice and A. thaliana, respectively (Fig. 3B) [101].
Ethylene (CH2=CH2, ETH) is a well-known small molecule gas that acts as a plant hormone, widely utilized in agriculture. Found in various organs and tissues across angiosperms, gymnosperms, ferns, and mosses, ethylene plays a vital role in numerous plant physiological processes and responses to biotic and abiotic stimuli [102,103]. It influences processes such as seed germination, inhibition of root elongation, regulation of plant hypocotyl elongation and apical hook development, as well as the stimulation of flower and leaf senescence and fruit ripening [102,103].
Controlling ethylene biosynthesis and signaling is a major challenge in plant biotechnology. This can be managed by either slowing down ethylene production or using specific inhibitors. Although ethylene receptors and their associated signaling pathways have been identified, the exact molecular mechanisms of signal detection and transmission are not yet fully understood. Additionally, the gaseous nature of ethylene complicates biochemical studies involving both proteins and small molecules with similar functions. Further details are available in Supporting information and Fig. S2C (Supporting information).
Abscisic acid (ABA) is a plant hormone composed of 15 carbon atoms and is classified as isoprenoid or terpenoid [104]. Initially discovered in mature dry cotton husks, ABA was identified for its role in regulating cotton fruit shedding. This growth inhibitor significantly affects plant growth and development, influencing processes such as seed dormancy, embryonic development, leaf senescence, abscission, stomatal movement, and responses to biotic and abiotic stressors [105].
Inhibitors of ABA biosynthesis can trigger the release of dormancy in seeds, buds, and tubers of non-model plants, thereby enhancing agricultural yields. Additionally, specific inhibitors of ABA catabolism are essential for maintaining ABA homeostasis. The key small molecules involved in these processes are illustrated in Fig. 4A, with their regulatory effects detailed in Supporting information.
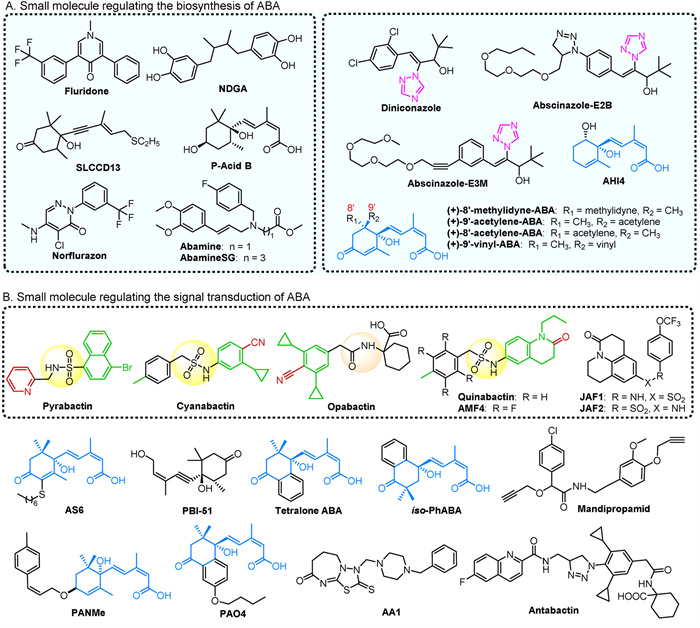
The functional redundancy within the ABA receptor protein family has drawn considerable interest. Small molecules are essential for distinguishing and characterizing receptor-specific regulatory mechanisms and uncovering other potential functions. Pyrabactin (Fig. 4B), an ABA agonist from the naphthalene sulfonamide class, strongly activates the PYR1 and PYL1 receptors, which are predominantly found in seeds [106]. Pyrabactin features a U-shaped structure that fits into the ligand pocket of PYL1, with its bromo ring and pyridine group occupying critical sites. The interaction between pyrabactin and PYL1 includes polar and van der Waals contacts, with the Glu121 carboxylate forming hydrogen bonds with pyrabactin's sulfonamide group [106]. The complex formed by OsPYL/RCAR3 with pyrabactin and OsPP2C50 closely mimics the OsPYL/RCAR3:ABA: OsPP2C50 complex, as pyrabactin binds in a similar pocket to ABA [107]. Despite the high sequence similarity between PYL1 and PYL2, pyrabactin preferentially binds to PYL1 due to a single amino acid difference between valine and isoleucine [108]. Pyrabactin also binds to PYL3 in a nonproductive manner, leading to gate closure and a decreased PP2C binding affinity. This compound inhibits seed germination, and hypocotyl growth, and may also affect stomatal function, suggesting its potential as an anti-transpiration agent or as a means to enhance drought tolerance [109]. While pyrabactin effectively triggers ABA response in seeds, its impact on vegetative tissues of A. thaliana is limited. In contrast, quinabactin, a screened compound, alleviates various phenotypes in the ABA-deficient mutant aba2 [110]. Upon binding, the dihydroquinolinone ring of quinabactin rotates, enhancing its binding within the active site of PYL [151]. Quinabactin primarily acts through PYR1 and PYL1, leading to responses such as guard cell closure, inhibited seed germination, reduced leaf water loss, and improved drought tolerance [111]. This effect promotes stomatal closure, induces stress response gene expression, and has the potential to enhance drought resistance through overexpression in PYL2 transgenic plants [112].
Cyanabactin, designed to interact with the PYL receptors, is a small molecule that shows improved selectivity in reducing transpiration and water loss compared to pyrabactin and quinabactin [112]. It is particularly effective at activating subfamily III ABA receptors, with a preference for PYR1. Structural analysis of the PYR1-cyanabactin complex reveals that its cyclopropyl ring extends into the 3′-tunnel. The arylnitrile and sulfonamide groups of cyanabactin mimic the cyclohexanone oxygen and carboxylic acid of ABA, facilitating hydrogen bonding in the tryptophan lock, which is essential for stable receptor activation [112]. Opabactin also activates PYR1, PYL1, and PYL2 receptors, demonstrating more potent and lasting anti-transpiration effects compared to existing sulfonamide-based ligands. It is effective in protecting wheat plants against drought and has a significant inhibitory effect on seed germination in A. thaliana, approximately ten times greater than ABA [113]. In contrast, JFA1 and JFA2 excessively stimulate PYR1 and PYL1, leading to inhibited seed germination and cotyledon greening [114]. The compound (+)-3′-butyl ABA induces transcriptional responses and stomatal closure but has only a modest impact on seed drought resistance (Fig. 4B) [114]. A new ABA analog, ASn (where n denotes the length of the alkyl chain), particularly AS6 with a six-carbon alkyl chain, disrupts the PYL/PP2C interaction and inhibits various stress-triggered ABA responses in vivo through steric hindrance. Despite its six-carbon alkyl chain, ASn closely resembles ABA in structure [115]. PBI-51, an acetyl analog of ABA, has been shown to suppress the expression of ABA-regulated genes during cabbage seed germination and inhibit ABA-induced stomatal closure [116]. Mandipropamid, used to manipulate ABA signaling, enhances plant survival under drought conditions by interacting with mutated residues in the PYR1 pocket. It mimics ABA effects by promoting stomatal closure in transgenic plants with altered PYR1 and inhibiting seed germination (Fig. 4B) [117].
4′-Phenylpropyl ABA analogs, known as PANs, are novel PYL antagonists. PANMe effectively disrupts the ABA-induced interaction between PYL-PP2C in vitro and shows stronger inhibition of stress-induced ABA responses compared to AS6 [118]. At concentrations three times higher than ABA, PANMe can also reverse the delayed seed germination in A. thaliana caused by ABA. Another analog, AA1, binds to the ligand-binding pocket of PYL2, blocking ABA signaling. This blockage results in delayed ripening of tomatoes, postponed senescence in rice, and inhibition of leaf chlorophyll degradation [119]. Antabactin (ANT), a derivative of opabactin, demonstrates how ANT sterically hinders access to the 4′ channel, a critical site where a conserved tryptophan residue binds to PP2C [120]. ANT exhibits greater potency than PANMe and AA1 in antagonizing ABA-mediated responses. The monocyclohexenone ring of ABA is expanded into a bicyclic ring to form tetealone ABA, which shows higher activity than ABA by extending into the ligand binding pocket typically occupied by the 7′-methyl group of ABA. Tetealone ABA and its isomer, iso-PhABA, engage in hydrogen bonding and multiple hydrophobic interactions with PYL10, demonstrating superior coordination within the binding pocket compared to ABA [121]. Additionally, the AS6 analog PAO4 shows enhanced binding affinity to PYL5, leading to antagonistic effects on seed germination and root growth. PBI686 acts as an antagonist of the PYL-PP2C interaction (Fig. 4B) [122,123].
ABA is a key stress signaling molecule in plants, and modifying its structure can enhance its effectiveness in managing abiotic and biotic stresses in agriculture. Several ABA derivatives have been synthesized based on its core structure (Fig. 5). For example, compound 44 shows a moderate effect on corn cell growth but is less potent than ABA. At a concentration of 33 µmol/L, its inhibitory effect is comparable to ABA at 10 µmol/L [124]. In contrast, compound 45 can induce stomatal closure without affecting seed germination or root growth. Its inhibition of PP2C is linked to its interaction with PYL5 and/or PYL6, with a binding affinity (Ka) of 0.88 µmol/L, making it 5–20 times more effective than ABA [125]. Among compounds 47–49, compound 47b, at 2 mg/L promotes stomatal closure and reduces water loss, enhancing drought tolerance in tobacco seedlings. Compounds 47a and 49 affect the elongation of rice seedlings and the germination of lettuce seeds, respectively [126]. Compound 47 specifically triggers ABA-mediated stomatal responses in A. thaliana, while compound 48 shows increased receptor binding affinity with a longer alkoxy chain [127,128]. Compounds 50a and 50b are used for detecting PYL. The hydroxyl group at position 1 is crucial for seed germination and development, and replacing it with –OCH3 reduces inhibitory efficacy by disrupting hydrogen bonds [129,130]. iso-PhABA analog 50c has slightly lower efficacy in inhibiting seed germination and rice seedling growth compared to iso-PhABA [129]. The structure-activity relationship indicates that the number and placement of -CH3 groups, as well as the substitution of hydrogen with -CH3, significantly impact the compound's activity [129]. A quaternary carbon at the 2′ or 3′ position is essential for maintaining ABA-like biological activity, particularly with demethylation at the 3′ position enhancing efficacy [129]. Due to ABA's photolability, its agricultural use is limited. However, photostable analogues 52a/52b, with the α, β-unsaturated carbonyl group of 52a reduced to 52b, show similar ABA activity. These analogues inhibit seed germination in tomatoes, lettuce, and rice, and promote drought tolerance in A. thaliana. Compound 52b does not activate PYL in vitro and may have higher membrane permeability due to ester substitution for the hydrophilic group (-COOH) in compound 52a [131].
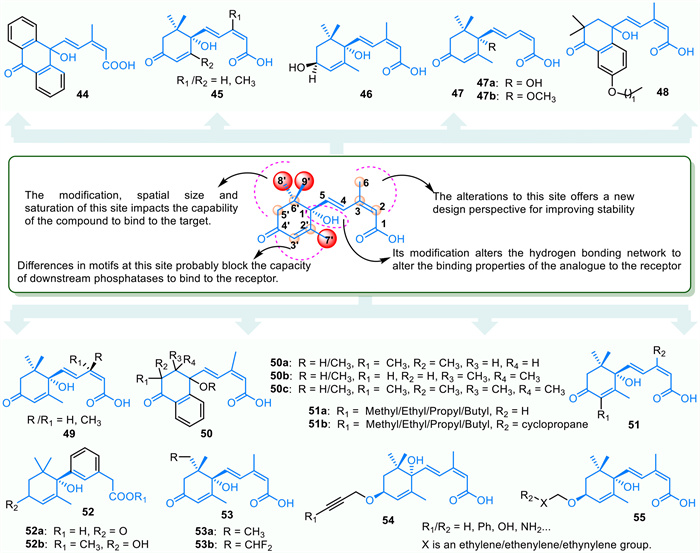
Compounds 51a/51b feature a cyclopropyl compound at the 3-position, enhancing their binding affinity and specificity by interacting with a specific receptor group in the C-6 cleft of the ABA binding pocket. These compounds increase the inhibitory activity of PYR1 and PYL1 on PP2C, with 3′-butyl ABA showing the highest drought stress resilience [132]. Compounds 53a/53b act as potent competitive inhibitors of ABA8'-hydroxylase, with 53a being a promising candidate for developing selective inhibitors [133]. Additionally, compounds 54 and 55, patented by Todoroki et al., are ABA derivatives modified at the 4′ position. They exhibit antagonistic effects on ABA receptors and promote seed germination in A. thaliana [134].
Jasmonic acid and its derivatives, collectively known as jasmonates (JA), including methyl jasmonate (Me-JA) and isoleucine jasmonate (JA-Ile), play crucial roles in regulating plant cell defense and development [135]. Me-JA was first isolated in 1962, with its physiological function confirmed in 1980. Jasmonic acid itself exhibits low biological activity, whereas JA-Ile demonstrates high activity [136]. JA extensively regulates various plant growth and development processes, including root elongation, anthocyanin accumulation, epidermal trichome development, male fertility, leaf senescence, and responses to abiotic stresses such as drought, high temperature, cold, salinity, and heavy metals [137].
Phytohormones play a crucial role in regulating various physiological processes in plants, such as development, differentiation, reproduction, stress tolerance, and immune response. Consequently, there is a growing interest in understanding the molecular mechanisms that govern growth and defense. Discovering small molecules through chemical biology techniques is essential for manipulating pathways in specific tissues, developmental stages, or species lacking mutants. Utilizing and absorbing small molecules by plants presents a viable alternative [138].
Inhibitors of JA biosynthesis, such as diethyldithiocarbamic acid (DIECA), can block JA production by targeting the octadecane signaling pathway. This inhibition causes hormonal changes and alters gene expression in plants [139]. Applying 10 mmol/L DIECA two days before infection has been shown to enhance wheat's resistance to powdery mildew, improving plant immunity and reducing disease severity [140]. Another JA synthesis inhibitor, salicyl hydroxamic acid (SHAM), impedes the increase in Rg1 levels induced by UV-B, highlighting the important roles of nitric oxide (NO) and JA in UV-B responses [141]. Moreover, Jarin-1, a selective inhibitor of the JAR1 enzyme, has been identified as an effective outcome of screening efforts in the JA signaling chemical library. This compound disrupts JA-Ile biosynthesis and selectively interferes with various JA-mediated processes in A. thaliana (Fig. 6B) [142].
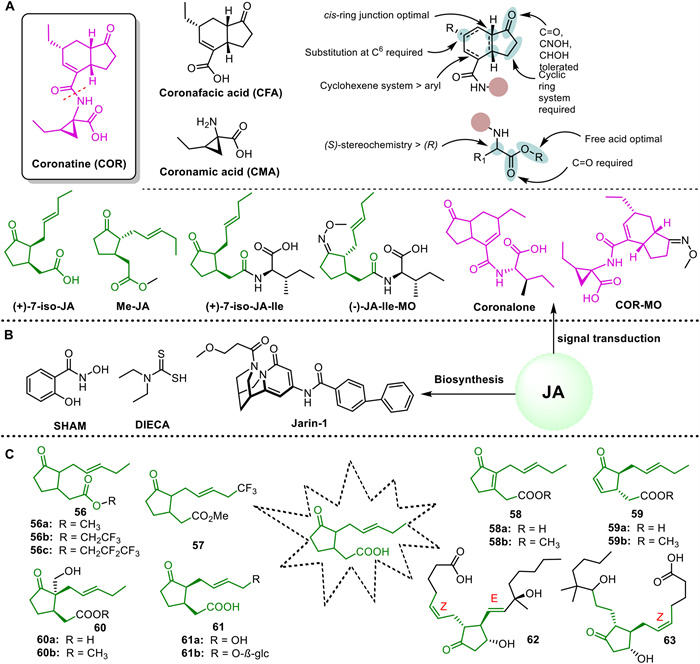
Coronatine (COR), derived from Pseudomonas syringae pv. atropurpurea, acts as a non-host-specific agonist of JA-Ile. It consists of coronanic acid (CMA) fused with α-amino acids and coronafacic acid (CFA) with a polyketide structure, connected by amide bonds. The biological activity of COR is influenced by the cis stereochemistry of the CMA ring, the C-6 position substitution, the α-carbon substitution retaining S-stereochemistry of CMA, and the free carboxyl terminus of the amino acid (Fig. 6A) [143]. Similar to JA-Ile, COR inhibits the degradation pathway of JA-Ile, resulting in increased stability and binding affinity to COI1. Consequently, COR is preferred over JA in various applications [144,145]. Applying COR on wheat under drought and normal conditions enhances resistance by improving water metabolism, photosynthesis, antioxidant enzyme activity, and reducing MDA and superoxide content [146]. Additionally, coronalone, an analog of COR, elicits JA-like responses in plants, stimulating volatile compound production and defense mechanisms against insect predation [142]. Building upon COR, JA-Ile-MO, and COR-MO have been synthesized, with COR-MO proving to be a potent antagonist, impeding COI1-JAZ interaction, JAZ degradation, and obstructing the JA signaling pathway as a competitive antagonist [147].
Derivatization of the JA structure is a method used to produce highly active compounds. For instance, JA derivative 1c has been shown to trigger oxidation reactions in plant defense mechanisms and activate L-phenylalanine ammonia lyase. Compared to 56a (Me-JA) and 56b, compound 56c serves as a chemical signal for inducing secondary metabolism in plant cell cultures [148]. Analog 57 of Me-JA effectively promotes potato tuber formation while inhibiting the growth of rice seedlings and the germination of lettuce and radish seeds [149]. Analog 58b exhibits stronger inhibition of lettuce seed germination than Me-JA, whereas 59b shows comparable activity to Me-JA [150]. Compounds 60a and 60b demonstrate activity in inducing the expansion of potato tuber cells, while 61a and 61b are significant JA metabolites [151]. The JA receptor COI1 is a key target involved in JA-mediated immunity through signal interaction. Virtual screening has identified JA analogues 62 and 63 with high binding affinity to COI1, with compound 62 being particularly efficient, stable, and exhibiting good cell permeability (Fig. 6C) [152].
Strigolactones (SLs) consist of a tricyclic lactone (ABC ring) and a monocyclic lactone (D ring) connected by an enol ether bond. Typical SLs, as shown in Fig. S4 (Supporting information), are classified into strigol-type SLs (A) and orobanchol-type SLs (B). Noncanonical SLs (C) lack a tetracyclic structure but contain a D ring [153]. SL governs various facets of plant development, including root architecture, seed germination, leaf senescence promotion, and secondary growth regulation [154]. Notably, SL inhibit lateral branching and significantly influence root growth.
Nitrogen-containing heterocyclic compounds, such as triazole and imidazole, are known for their ability to inhibit various P450 enzyme activities and are frequently found in inhibitors of SL biosynthesis (Fig. 7A) [155]. The triazole compound TIS13 promotes the growth of secondary tiller buds in wild-type seedlings while decreasing SL levels in roots and their exudates, an effect reversible by the application of GR24 [82]. TIS108 is more effective, reducing SL levels, particularly epi-5DS, in roots and their secretions without significantly affecting leaf sheath length [156]. Its derivative KK5, demonstrates even greater inhibition of SL levels compared to TIS108. Treatment with these compounds in A. thaliana increases branching and up-regulates the expressions of AtMAX3 and AtMAX4 genes [157]. Triflumizole has also been shown to effectively decrease the levels of 4-deoxyorobanchol (4DO) in rice root exudates by inhibiting Os900 enzyme activity, which converts the SL intermediate carlactone into 4DO [158]. Despite the limited literature on SL biosynthesis inhibitors, this area holds significant research potential and value.
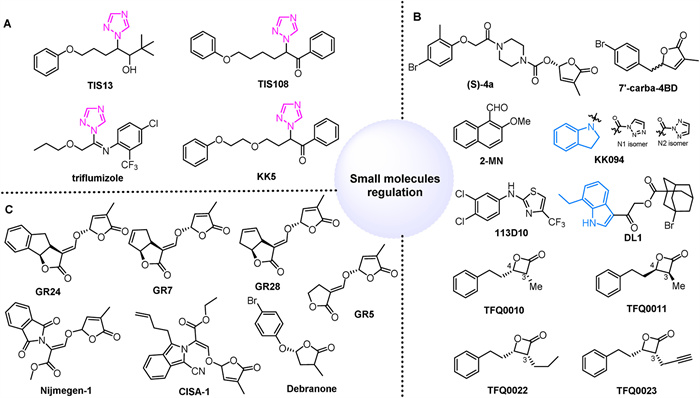
The development of selective agonists or antagonists for specific SL receptors is crucial for improving crop production due to the diverse receptors and functions of SL (Fig. 7B). SLs significantly influence the seed germination of most parasitic weeds, highlighting the need for potent SL agonists to control root parasitic weeds. A small molecule agonist, (S)-4a, identified through virtual screening and structure optimization, effectively induces seed germination in Phelipanchcea egyptiaca and Striga hermonthica at concentrations as low as 10–8-10–17 mol/L, demonstrating superior germination activity compared to GR24 [159]. Notably, it is important to note that the majority of SL agonists contain D-ring or D-ring-like structures. However, compound 113D10 functions as a unique SL agonist in a D14-dependent manner without an enol ether bond connected to the methylbutenolide moiety. Unfortunately, although it does not trigger seed germination in root parasitic plants [160].
Strigolactone antagonists can block the biological reactions triggered by agonist-activated receptors, regulate crop yields by altering tiller numbers, and prevent seed germination in root parasitic plants for crop protection [161]. Carba-SL inhibits the interaction between D14 and D53 by suppressing D14 hydrolysis activity, thereby inhibiting SL-induced tillering in rice. The D ring of 7′-carba-4BD remains attached to the skeleton upon hydrolysis by D14, functioning as an antagonist rather than an agonist [162]. KK094, a regioisomeric mixture (KK094-N1 and KK094-N2), forms a covalent bond with D14, and upon degradation, the product binds covalently to the Ser residue of the D14 catalytic triad, with its indole structure playing a crucial role [163]. Additionally, 2-methoxy-1-naphthaldehyde (2-MN) inhibits the interaction between D14-SLR1 and D14-D53, restoring the growth of rice tiller buds inhibited by SL [164]. DL1 inhibits SL-induced expression of BRC1 at submicromolar levels, acting locally within buds to suppress rosette branch outgrowth. Structure-activity relationship studies reveal that DL1's ethyl indole moiety is positioned close to the D14 catalytic site, significantly enhancing its inhibitory potency [165]. TFQ0010 has been identified as a potent inhibitor of AtD14, while its enantiomer TFQ0011 exhibits lower potency, indicating a specific chiral environment in the binding pocket. Compounds TFQ0022 and TFQ0023, with propyl and propargyl groups at C-3, inhibit the SL-induced interaction between AtD14 and MAX2 in vitro, consequently restraining hypocotyl elongation in A. thaliana seedlings [166].
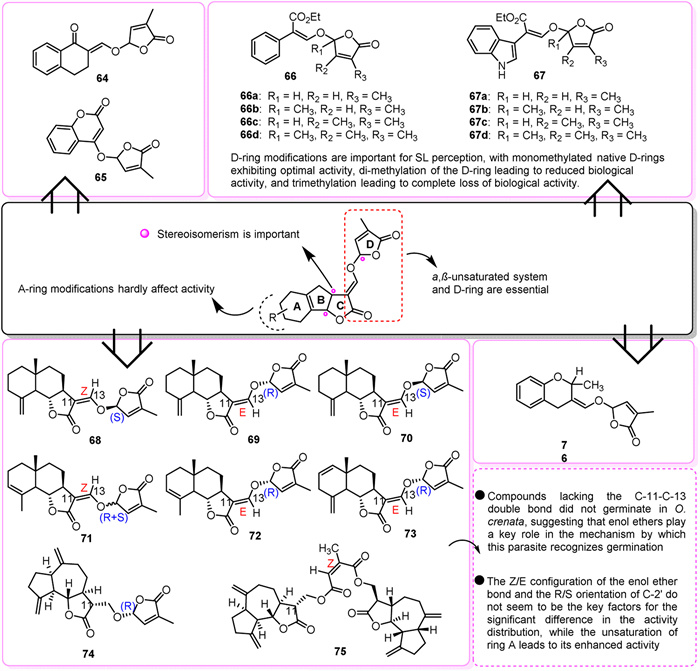
Given the complexity of strigolactones (SL) and limited synthetic options, efforts have focused on identifying the minimal structure required to trigger seed germination. A D-ring linked by an enol ether group is recognized as the essential core for creating SL analogues, with GR24 being a prominent example (Fig. 7C). GR24 is involved in various physiological processes, including the germination of A. thaliana seeds, potato tuber formation, and bud branching. It also suppresses lateral root density in alfalfa [167]. Application of GR24 results in altered photosynthesis and transpiration rates, stomatal conductance, leaf water potential, and oxidase activity, enhancing drought resistance and yield [168,169]. Simplified compounds like GR7, GR5, and GR28 also stimulate germination, with the C-2′ (R configuration) stereochemistry being crucial for optimal activity. Among these, GR24 has shown the highest effectiveness in stimulating the germination of parasitic weed seeds and greater hydrolysis resistance compared to GR7 and GR28, making it a commonly used compound in SL research [170]. However, the structurally simplified debranone is less effective in inducing the germination of Stringa hermonthica seeds and is unstable in alkaline soil, limiting its agriculture use [170]. Consequently, Nijmegen-1 was synthesized based on the structure-activity relationship of GR-type compounds and used as a suicide germination agent for tobacco infected with Orobanche ramosa L [171]. Similarly, CISA-1 is a fluorescent alternative to GR24 that has shown enhanced effectiveness in inhibiting adventitious roots and promoting the germination of orobanchaceae plants. Both compounds are equally potent in suppressing branching in A. thaliana and pea plants [172].
Several analogs, such as Nijmegen-1, were designed and synthesized based on the structural model of natural SLs with a D ring connected by an enol ether group. In pot experiments, compounds 64 and 65 showed promise as suicidal germinants [173]. Compounds 66a and 67a, both containing monomethylated native D rings, proved exceptionally effective, demonstrating potency comparable to GR24 against P. ramosa and significantly higher activity against O. minor. In contrast, compounds 66d and 67d, featuring trimethylated D rings, displayed no activity [174]. Additionally, compounds 66b/66c showed higher activity compared to 67b/67c, highlighting the significance of D-ring modification in SL sensing. Dimethylation resulted in decreased biological activity, while trimethylation led to a complete loss of activity [174]. Among the tested analogs, compounds 69, 70, 72, 74, and 75 were most effective in promoting P. ramosa germination. For O. cumana, compounds 71 and 72 showed the highest effectiveness, and for O. crenata, compounds 71 and 72 displayed the highest germination stimulation rates [175]. The presence of a 2-alkyl group in the dihydroflavone skeleton was found to be crucial for seed germination activity. Compound 76, containing a -CH3 group, exhibited tenfold higher germination activity compared to the control GR24 [176,177]. Strigolactones play a role in signaling the development of arbuscular mycorrhizal fungi and act as triggers for the germination of parasitic weed seeds (Fig. 8).
Recent research highlights the increasing importance of PGRs in pesticides due to their low dosage requirements, rapid action, high efficacy, minimal residual toxicity, and precise targeting. The biosynthesis, degradation, transportation, and signal transduction of natural plant hormones involve various enzymes and receptors, presenting numerous opportunities for chemical intervention. Studying these interactions may lead to the design of novel chemical compounds that promote plant growth and enhance stress resilience.
In conclusion, for ABA, modifications in spatial dimensions, and saturation at the 5′/6′ and 8′/9′ positions are crucial for determining binding capacity. Differences at the 2′/3′ position can hinder the downstream interaction of phosphatase with the receptor, while changes at the 2/3/6′ position can enhance stability. Modifications at the 1′ position alter the hydrogen bond network, impacting the interaction between the analog and the receptor. The small molecules analyzed in this research feature active fragments/groups like sulfonamide, amide, and triazole, essential for ABA synthesis and signaling pathways. For GAs, key biological activity traits include the 3β-hydroxyl group, the absence of a hydroxyl group at the C-2 position, the C-6 carboxyl, and a C-lactone ring. SLs exhibit structural and functional diversity due to various groups in different configurations or positions on the ABC-ring while maintaining a conserved D-ring structure essential for biological activity. CTK shows varying activities with substituents at different positions of the adenine structure, especially at the 2/4/6 positions, and non-adenine compounds like urea derivatives also act as cytokinin analogs. Structural variations in BRs predominantly are mainly due to differences in substituents on the A/B carbon ring and the alkane side chain. The cyclopentanone in JA and the indole ring in IAA are pivotal core structures, with their free carboxylic acid groups being significant.
Plant hormones, their inhibitors, and analogs contribute to the diverse effects, broad biological activities, low toxicity, and environmental compatibility of pesticides derived from them. The investigation of these PGRs with varied chemical structures remains a prominent research area. Although many compounds exhibit PGR activity, their suitability for agricultural use is currently limited. The complexity of PGRs activity mechanisms and targets further constrains their application. To address these challenges involves developing and characterizing highly selective and potent small molecule inhibitors or agonists through integrated approaches such as synthetic chemistry, computational chemistry, biology, high-throughput screening, and molecular modeling. Additionally, employing diverse methods including biochemistry, molecular biology, structural biology, genetics, and bioinformatics can elucidate the underlying regulatory mechanisms of plant hormone signaling networks. These research methodologies and findings offer valuable theoretical insights for drug discovery and present novel strategies for plant manipulation and the advancement of sustainable agricultural practices.
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Ali Dai: Writing – original draft, Visualization, Validation, Investigation, Conceptualization. Zhiguo Zheng: Writing – original draft, Visualization, Validation, Investigation, Conceptualization. Liusheng Duan: Writing – original draft, Supervision, Conceptualization. Jian Wu: Writing – review & editing, Supervision, Conceptualization. Weiming Tan: Writing – review & editing, Supervision, Funding acquisition, Conceptualization.
The financial support from the National Key Research and Development Program of China (No. 2023YFD1700600).
Supplementary material associated with this article can be found, in the online version, at doi:
Y. Wang, S.X. Guo, L.J. Yu, et al., Chin. Chem. Lett. 35 (2024) 108207.
A.L. Dai, Z.G. Zheng, Y.Q. Huang, et al., Heliyon 8 (2022) e12391.
S. Katel, H.R. Mandal, S. Kattel, S.P.S. Yadav, B.S. Lamshal, Heliyon 8 (2022) e11959.
D.K. Tripathi, S.R. Yadav, K. Mochida, L.S.P. Tran, Plant Cell Physiol. 63 (2023) 1757–1760. doi: 10.1093/pcp/pcac170
B. Desta, G. Amare, Chem. Biol. Technol. Agric. 8 (2021) 15.
B. Noein, A. Soleymani, J. Plant Growth Regul. 41 (2022) 672–681. doi: 10.1007/s00344-021-10332-3
J. Takeuchi, K. Fukui, Y. Seto, Y. Takaoka, M. Okamoto, Plant J. 105 (2021) 290–306. doi: 10.1111/tpj.15115
A. Bajguz, A. Piotrowska-Niczyporuk, Metabolites 13 (2023) 884. doi: 10.3390/metabo13080884
S. Hagihara, R. Yamada, K. Itami, K.U. Torii, Curr. Opin. Plant Biol. 47 (2019) 32–37.
V. Gomez-Roldan, S. Fermas, P.B. Brewer, et al., Nature 455 (2008) 189–194. doi: 10.1038/nature07271
R.D. Crapnell, P.S. Adarakatti, C.E. Banks, Anal. Methods 15 (2023) 4811–4826. doi: 10.1039/d3ay01053h
Z.Y. Lv, L. Zhang, K.X. Tang, Plant Signaling Behav. 12 (2017) e1366398. doi: 10.1080/15592324.2017.1366398
S.A.J. Messing, S.B. Gabelli, I. Echeverria, et al., Plant Cell 22 (2010) 2970–2980. doi: 10.1105/tpc.110.074815
A. Rigal, Q. Ma, S. Robert, Front. Plant Sci. 5 (2014) 373.
K. Jiang, T. Asami, Biosci. Biotechnol. Biochem. 82 (2018) 1265–1300. doi: 10.1080/09168451.2018.1462693
S.Y. Park, P. Fung, N. Nishimura, et al., Science 324 (2009) 1068–1071. doi: 10.1126/science.1173041
R. Varela, A.E. Cleves, R. Spitzer, A.N. Jain, J. Comput-Aided. Mol. Des. 27 (2013) 917–934. doi: 10.1007/s10822-013-9688-9
M. Lavy, M. Estelle, Development 143 (2016) 3226–3229. doi: 10.1242/dev.131870
H. Kasahara, Biosci. Biotechnol. Biochem. 80 (2016) 34–42. doi: 10.1080/09168451.2015.1086259
O. Leyser, Plant Physiol. 176 (2018) 465–479. doi: 10.1104/pp.17.00765
W. He, J. Brumos, H. Li, et al., Plant Cell 23 (2011) 3944–3960. doi: 10.1105/tpc.111.089029
J. Basran, E.S. Booth, L.P. Campbell, et al., J. Inorg. Biochem. 225 (2021) 111604.
D.M. Eklund, K. Ishizaki, E. Flores-Sandoval, et al., Plant Cell 27 (2015) 1650–1669. doi: 10.1105/tpc.15.00065
Y. Kakei, C. Yamazaki, M. Suzuki, et al., Plant J. 84 (2015) 827–837. doi: 10.1111/tpj.13032
T. Nishimura, K. Hayashi, H. Suzuki, et al., Plant J. 77 (2014) 352–366. doi: 10.1111/tpj.12399
S. Tsugafune, K. Mashiguchi, K. Fukui, et al., Sci. Rep. 7 (2017) 13992.
Y. Kakei, A. Nakamura, M. Yamamoto, et al., Plant Cell Physiol. 58 (2017) 598–606.
P. Xu, H. Jinbo, W. Cai, New Phytol. 236 (2022) 1748–1761. doi: 10.1111/nph.18459
W. Teale, K. Palme, J. Exp. Bot. 69 (2018) 303–312. doi: 10.1093/jxb/erx323
J.Y. Kim, S. Henrichs, A. Bailly, et al., J. Biol. Chem. 285 (2010) 23309–23317.
N. Geldner, J. Friml, Y.D. Stierhof, G. Jürgens, K. Palme, Nature 413 (2001) 425–428.
T. Nishimura, N. Matano, T. Morishima, et al., Cell. Physiol. 53 (2012) 1671–1682. doi: 10.1093/pcp/pcs112
K. Fukui, K.I. Hayashi, Plant Cell Physiol. 59 (2018) 1500–1510. doi: 10.1093/pcp/pcy076
E. Tsuda, H. Yang, T. Nishimura, et al., J. Biol. Chem. 286 (2011) 2354–2364. doi: 10.1074/jbc.M110.171165
M. Lanková, R.S. Smith, B. Pesek, et al., J. Exp. Bot. 61 (2010) 3589–3598. doi: 10.1093/jxb/erq172
W. Steenackers, I. El Houari, A. Baekelandt, et al., J. Exp. Bot. 70 (2019) 6293–6304. doi: 10.1093/jxb/erz392
W. Steenackers, P. Klíma, M. Quareshy, et al., Plant Physiol. 173 (2017) 552–565. doi: 10.1104/pp.16.00943
W. Steenackers, I. Cesarino, P. Klíma, et al., Plant Physiol. 172 (2016) 874–888.
B. Hartwig, G.V. James, K. Konrad, K. Schneeberger, F. Turck, Plant Physiol. 160 (2012) 591–600. doi: 10.1104/pp.112.200311
K. Hayashi, X. Tan, N. Zheng, et al., Proc. Natl. Acad. Sci. U. S. A. 105 (2008) 5632–5637. doi: 10.1073/pnas.0711146105
K. Hayashi, J. Neve, M. Hirose, et al., ACS Chem. Biol. 7 (2012) 590–598. doi: 10.1021/cb200404c
Y. Hu, L. Yang, X. Na, et al., Planta 236 (2012) 597–612. doi: 10.1007/s00425-012-1632-z
A. Yamazoe, K. Hayashi, S. Kepinski, O. Leyser, H. Nozaki, Plant Physiol. 139 (2005) 779–789. doi: 10.1104/pp.105.068924
R. Pelagio-Flores, R. Ortíz-Castro, A. Méndez-Bravo, et al., Plant Cell Physiol. 52 (2011) 490–508. doi: 10.1093/pcp/pcr006
N. Uchida, K. Takahashi, R. Iwasaki, et al., Nat. Chem. Biol. 14 (2018) 299– 305. doi: 10.1038/nchembio.2555
P. Sun, Y.Q. Huang, X.Y. Yang, A.J. Liao, J. Wu, Front. Plant Sci. 13 (2023) 1120613.
P. Sun, Y.Q. Huang, S.H. Chen, et al., Chin. Chem. Lett. 35 (2024) 109005.
M. Quareshy, J. Prusinska, M. Kieffer, et al., ACS Chem. Biol. 13 (2018) 2585–2594. doi: 10.1021/acschembio.8b00527
B. Wybouw, B. De Rybel, Trends Plant Sci. 24 (2019) 177–185.
D. Kopecný, P. Briozzo, H. Popelková, et al., Biochimie 92 (2010) 1052–1062.
N. Murvanidze, J. Nisler, O. Leroux, S.P.O. Werbrouck, Plant Growth Regul. 94 (2021) 195–200. doi: 10.1007/s10725-021-00708-6
J. Nisler, D. Kopečný, R. Končitíková, et al., Plant. Mol. Biol. 92 (2016) 235–248. doi: 10.1007/s11103-016-0509-0
M. Zatloukal, M. Gemrotová, K. Dolezal, et al., Bioorg. Med. Chem. 16 (2008) 9268–9275.
M. J. van Voorthuizen, J. Song, O. Novák, P.E. Jameson, Plants 10 (2021) 2309.
L. Spíchal, T. Werner, I. Popa, et al., FEBS J. 276 (2009) 244–253. doi: 10.1111/j.1742-4658.2008.06777.x
J. Nisler, M. Zatloukal, I. Popa, et al., Phytochemistry 71 (2010) 823–830.
Y. Arata, A. Nagasawa-Iida, H. Uneme, et al., Plant Cell Physiol. 51 (2010) 2047–2059. doi: 10.1093/pcp/pcq163
M.S. Oshchepkov, A.V. Kalistratova, E.M. Savelieva, et al., Russ. Chem. Rev. 89 (2020) 787–810. doi: 10.1070/rcr4921
R. Koprna, J.F. Humplík, Z. Spísek, et al., Agronomy-Basel 11 (2021) 15.
K. Podlešáková, D. Zalabák, M. Cudejková, et al., PLoS One 7 (2012) e39293. doi: 10.1371/journal.pone.0039293
M. Bryksová, A. Hybenová, A.E. Hernándiz, et al., Front Plant Sci. 11 (2020) 599228.
E.M. Savelieva, A.A. Zenchenko, M.S. Drenichev, et al., Int. J. Mol. Sci. 23 (2022) 11334. doi: 10.3390/ijms231911334
V. Mik, L. Szücová, L. Spíchal, et al., Bioorg. Med. Chem. 19 (2011) 7244–7251.
B. Zhang, H. Gao, G. Wang, et al., Front. Plant. Sci. 13 (2022) 1025634.
C. Liu, L. Bai, P. Cao, et al., J. Agric. Food Chem. 70 (2022) 16229–16240. doi: 10.1021/acs.jafc.2c07072
E.M. Savelieva, B.V.E. Oslovsky, D.S. Karlov, et al., Phytochemistry 149 (2018) 161–177.
H. Iwamura, N. Masuda, K. Koshimizu, S. Matsubara, Phytochemistry 18 (1979) 217–222.
V. Matušková, M. Zatloukal, T. Pospíšil, et al., Phytochemistry 205 (2023) 113481.
A. Bajguz, A. Tretyn, Phytochemistry 62 (2003) 1027–1046.
J.W. Mitchell, N. Mandava, J.F. Worley, J.R. Plimmer, M.V. Smith, Nature 225 (1970) 1065–1066. doi: 10.1038/2251065a0
C.J. Yang, C. Zhang, Y.N. Lu, J.Q. Jin, X.L. Wang, Mol. Plant 4 (2011) 588–600. doi: 10.1093/mp/ssr020
M. Mizutani, D. Ohta, Annu. Rev. Plant Biol. 61 (2010) 291–315. doi: 10.1146/annurev-arplant-042809-112305
T.G. Back, R.P. Pharis, J. Plant Growth Regul. 22 (2003) 350–361.
S. Takatsuto, N. Yazawa, N. Ikekawa, T. Morishita, H. Abe, Phytochemistry 22 (1983) 1393–1397.
M.A. Hussain, S. Fahad, R. Sharif, et al., Plant Growth Regul. 92 (2020) 141–156.
L.Y. Zou, M.H. Qu, L.J. Zeng, G.S. Xiong, Plant Growth Regul. 91 (2020) 263–276. doi: 10.1007/s10725-020-00604-5
M.V. Diachkov, K. Ferrer, J. Oklestkova, et al., Int. J. Mol. Sci. 22 (2020) 155. doi: 10.3390/ijms22010155
L.M. Mazorra, M. Núñez, M.C. Nápoles, et al., Plant Growth Regul. 44 (2004) 183–185.
M. Kvasnica, J. Oklestkova, V. Bazgier, et al., Org. Biomol. Chem. 14 (2016) 8691–8701.
S. Salazar-Cerezo, N. Martínez-Montiel, J. García-Sánchez, et al., Microbiol. Res. 208 (2018) 85–98.
Niharika, N.B. Singh, A. Singh, et al., Plant Mol. Biol. Rep. 39 (2021) 34–49. doi: 10.1007/s11105-020-01231-0
Z.K. Yang, J.H. Xu, Y.M. Bai, et al., J. Plant Growth Regul. 41 (2022) 1845–1853. doi: 10.1007/s00344-021-10425-z
S. Wang, H. Zhou, N. Feng, et al., J. Plant Physiol. 268 (2022) 153579.
I. Ahmad, S. Ahmad, X.N. Yang, et al., Plant Biol. 23 (2021) 485–496. doi: 10.1111/plb.13235
H. Zhou, X. Liang, N. Feng, D. Zheng, D. Qi, Ecotoxicol. Environ. Saf. 224 (2021) 112619.
M. Waqas, C. Yaning, H. Iqbal, et al., J. Agron. Crop Sci. 203 (2017) 315–322. doi: 10.1111/jac.12217
N.M. Kleczewski, C. Whaley, Crop Prot 104 (2018) 60–64.
J.A.K. Trethewey, M.P. Rolston, B.L. McCloy, R.J. Chynoweth, N. Z. J. Agric. Res. 59 (2016) 113–121. doi: 10.1080/00288233.2015.1134590
N.L. Heckman, T.E. Elthon, G.L. Horst, R.E. Gaussoin, Crop Sci. 42 (2002) 423–427.
J.H. Grabber, Agron. J. 108 (2016) 726–735. doi: 10.2134/agronj2015.0466
R. Zhang, D. Zheng, N. Feng, Q.S. Qiu, H. Zhou, F. Meng, X. Huang, A. Huang, Y. Li, PLoS One 18 (2023) e0279192. doi: 10.1371/journal.pone.0279192
K. Murase, Y. Hirano, T.P. Sun, T. Hakoshima, Nature 456 (2008) 459–463. doi: 10.1038/nature07519
K. Jiang, M. Otani, H. Shimotakahara, et al., Plant Physiol. 173 (2017) 825–835. doi: 10.1104/pp.16.00937
J.D. Metzger, Weed Sci. 31 (1983) 285–289. doi: 10.1017/s0043174500069010
W.W. Donald, R.A. Hoerauf, Weed Sci. 33 (1985) 894–902.
K. Jiang, J.W. Wang, S. Ito, et al., J. Plant Growth Regul. 42 (2023) 2637–2645. doi: 10.1007/s00344-022-10733-y
K. Jiang, H. Shimotakahara, M. Luo, et al., Bioorg. Med. Chem. Lett. 27 (2017) 3678–3682.
T. Ariizumi, C.M. Steber, Plant Cell 19 (2007) 791–804. doi: 10.1105/tpc.106.048009
C. Bergner, M. Lischewski, G. Adam, G. Sembdner, Planta 155 (1982) 231–237.
T. Ishida, B. Watanabe, K. Mashiguchi, S. Yamaguchi, Phytochem. Lett. 49 (2022) 162–166.
H. Tian, Y. Xu, S. Liu, et al., Molecules 22 (2017) 694. doi: 10.3390/molecules22050694
Y.Y. Xiao, J.Y. Chen, J.F. Kuang, et al., J. Exp. Bot. 64 (2013) 2499–2510. doi: 10.1093/jxb/ert108
W. Zhang, Y. Hu, J. Liu, et al., Saudi J. Biol. Sci. 27 (2020) 1667–1673.
Y. Liu, S. Chen, P. Wei, S. Guo, J. Wu, Front. Chem. 10 (2022) 967404.
B.P. Brookbank, J. Patel, S. Gazzarrini, E. Nambara, Genes 12 (2021) 1936. doi: 10.3390/genes12121936
Q. Hao, P. Yin, C. Yan, et al., J. Biol. Chem. 285 (2010) 28946–28952. doi: 10.1074/jbc.M110.149005
S. Han, Y. Lee, E.J. Park, et al., Plant Mol. Biol. 100 (2019) 319–333. doi: 10.1007/s11103-019-00862-6
X.Q. Yuan, P. Yin, Q. Hao, et al., Plant Biol. 285 (2010) 28953–28958. doi: 10.1074/jbc.M110.160192
M. Okamoto, F.C. Peterson, A. Defries, et al., Proc. Natl. Acad. Sci. U. S. A. 110 (2013) 12132–12137. doi: 10.1073/pnas.1305919110
F. Dupeux, J. Santiago, K. Betz, et al., EMBO J. 30 (2011) 4171–4184. doi: 10.1038/emboj.2011.294
M. Cao, X. Liu, Y. Zhang, et al., Cell Res. 23 (2013) 1043–1054. doi: 10.1038/cr.2013.95
A.S. Vaidya, F.C. Peterson, D. Yarmolinsky, et al., ACS Chem. Biol. 12 (2017) 2842–2848. doi: 10.1021/acschembio.7b00650
A.S. Vaidya, J.D.M. Helander, F.C. Peterson, et al., Science 366 (2019) eaaw8848.
K. Nemoto, M. Kagawa, A. Nozawa, et al., Sci. Rep. 8 (2018) 4268.
J. Takeuchi, M. Okamoto, T. Akiyama, et al., Nat. Chem. Biol. 10 (2014) 477–482. doi: 10.1038/nchembio.1524
R.W. Wilen, D.B. Hays, R.M. Mandel, S.R. Abrams, M.M. Moloney, Plant Physiol. 101 (1993) 469–476. doi: 10.1104/pp.101.2.469
P.L. Rodriguez, J. Lozano-Juste, Trends Plant Sci. 20 (2015) 330–332.
J. Takeuchi, N. Mimura, M. Okamoto, et al., ACS Chem. Biol. 13 (2018) 1313–1321. doi: 10.1021/acschembio.8b00105
Y. Ye, L. Zhou, X. Liu, et al., Plant Physiol. 173 (2017) 2356–2369. doi: 10.1104/pp.16.01862
A.S. Vaidya, F.C. Peterson, J. Eckhardt, et al., Proc. Natl. Acad. Sci. U. S. A. 118 (2021) e2108281118.
X. Han, L. Jiang, C. Che, et al., Sci. Rep. 7 (2017) 43863.
N. Rajagopalan, K.M. Nelson, A.F. Douglas, et al., Biochemistry 55 (2016) 5155–5164. doi: 10.1021/acs.biochem.6b00605
J. Takeuchi, T. Ohnishi, M. Okamoto, Y. Todoroki, Org. Biomol. Chem. 13 (2015) 4278–4288.
N.M. Irvine, P.A. Rose, A.J. Cutler, T.M. Squires, S.R. Abrams, Phytochemistry 53 (2000) 349–355.
J. Takeuchi, T. Ohnishi, M. Okamoto, Y. Todoroki, Bioorg. Med. Chem. Lett. 25 (2015) 3507–3510.
T. Liu, C.X. Li, J. Zhong, et al., Int. J. Mol. Sci. 22 (2021) 2555. doi: 10.3390/ijms22052555
C.L. Benson, M. Kepka, C. Wunschel, et al., Phytochemistry 113 (2015) 96–107.
C. Che, Y. Zeng, Y. Xu, et al., J. Agric. Food Chem. 68 (2020) 8524–8534. doi: 10.1021/acs.jafc.0c02154
C. Wan, Q. Hong, X. Zhang, et al., Agric. Food Chem. 67 (2019) 4995–5007. doi: 10.1021/acs.jafc.8b07068
C. Wan, D.Y. Yang, R.Y. Liu, et al., J. Mol. Struct. 1249 (2022) 131650.
J. Takeuchi, S. Mimura, T. Ohnishi, Y. Todoroki, J. Agric. Food Chem. 70 (2022) 869–876. doi: 10.1021/acs.jafc.1c06321
K. Yoshida, Y. Kondoh, T. Nakano, et al., ACS Chem. Biol. 16 (2021) 1566–1575. doi: 10.1021/acschembio.1c00451
K. Ueno, H. Yoneyama, S. Saito, et al., Bioorg. Med. Chem. Lett. 15 (2005) 5226–5229.
K.A. Abu Safieh, A.K. Hasan, M.T. Ayoub, M.S. Mubarak, Res. Chem. Intermed. 41 (2015) 1863–1872. doi: 10.1007/s11164-013-1315-9
K. Gomi, Int. J. Mol. Sci. 21 (2020) 1261. doi: 10.3390/ijms21041261
J. Yan, S. Li, M. Gu, et al., Plant Physiol. 172 (2016) 2154–2164. doi: 10.1104/pp.16.00906
Y. Hu, Y. Jiang, X. Han, et al., J. Exp. Bot. 68 (2017) 1361–1369. doi: 10.1093/jxb/erx004
W. Zhang, S.X. Guo, L.J. Yu, et al., Chin. Chem. Lett. 34 (2023) 108123.
X.B. Wei, W.L. Guan, Y.J. Yang, Y.L. Shao, L.C. Mao, Postharvest Biol. Technol. 175 (2021) 111472.
Y. Li, L. Qiu, Q. Zhang, et al., Plant Direct 4 (2020) e00212.
J. Zhou, Z.F. Ran, X.T. Yang, J. Li, Molecules 24 (2019) 1462. doi: 10.3390/molecules24081462
M. Ueda, T. Kaji, W. Kozaki, Int. J. Mol. Sci. 21 (2020) 1124. doi: 10.3390/ijms21031124
M.M. Littleson, C.M. Baker, A.J. Dalençon, et al., Nat. Commun. 9 (2018) 1105.
S. Fonseca, A. Chini, M. Hamberg, et al., Nat. Chem. Biol. 5 (2009) 344–350. doi: 10.1038/nchembio.161
L.B. Sheard, X. Tan, H. Mao, et al., Nature 468 (2010) 400–405. doi: 10.1038/nature09430
Y.J. Wang, Z.H. Wang, S. Wang, et al., Acta Agric. Scand., Sect. B 70 (2020) 467–473.
M. Cui, K. Zhang, R. Wu, J. Du, J. Comput. Aided. Mol. Des. 36 (2022) 141–155. doi: 10.1007/s10822-022-00441-w
Z.G. Qian, Z.J. Zhao, Y. Xu, X. Qian, J.J. Zhong, Appl. Microbiol. Biotechnol. 68 (2005) 98–103. doi: 10.1007/s00253-004-1835-8
H. Kiyota, M. Saitoh, T. Oritani, T. Yoshihara, Phytochemistry 42 (1996) 1259–1262.
H. Kiyota, Y. Yoneta, T. Oritani, Phytochemistry 46 (1997) 983–986.
N. Kitaoka, E. Fukushi, Y. Koda, K. Nabeta, H. Matsuura, Biosci. Biotechnol. Biochem. 73 (2009) 1872–1876. doi: 10.1271/bbb.90148
R.K. Pathak, M. Baunthiyal, R. Shukla, et al., Front Plant Sci. 8 (2017) 609.
R.J. Chesterfield, C.E. Vickers, C.A. Beveridge, Trends Plant Sci. 25 (2020) 1087–1106.
A. Bhoi, B. Yadu, J. Chandra, S. Keshavkant, Planta 254 (2021) 28.
I. Takahashi, H. Koishihara, T. Asami, J. Pestic. Sci. 47 (2022) 197–202. doi: 10.1584/jpestics.d22-022
S. Ito, M. Umehara, A. Hanada, S. Yamaguchi, T. Asami, Plant Signal. Behav. 8 (2013) e24193. doi: 10.4161/psb.24193
K. Kawada, I. Takahashi, M. Arai, et al., J. Agric. Food Chem. 67 (2019) 6143–6149. doi: 10.1021/acs.jafc.9b01276
K. Kawada, Y. Uchida, I. Takahashi, et al., Molecules 25 (2020) 5525. doi: 10.3390/molecules25235525
D. Wang, Z. Pang, H. Yu, et al., Nat. Commun. 13 (2022) 3987.
R. Yasui, Y. Seto, S. Ito, et al., Bioorg. Med. Chem. Lett. 29 (2019) 938–942.
I. Takahashi, T. Asami, J. Exp. Bot. 69 (2018) 2241–2254. doi: 10.1093/jxb/ery126
J. Takeuchi, K. Jiang, K. Hirabayashi, et al., Plant Cell Physiol. 59 (2018) 1545–1554. doi: 10.1093/pcp/pcy087
H. Nakamura, K. Hirabayashi, T. Miyakawa, et al., Mol. Plant 12 (2019) 44–58.
O. Mashita, H. Koishihara, K. Fukui, H. Nakamura, T. Asami, J. Pestic. Sci. 41 (2016) 71–78.
M. Yoshimura, S.F. Kim, R. Takise, et al., Chem. Commun. 56 (2020) 14917–14919. doi: 10.1039/d0cc01989e
H. Xiang, R. Yao, T. Quan, et al., Cell Res. 27 (2017) 1525–1528. doi: 10.1038/cr.2017.105
C. De Cuyper, J. Fromentin, R.E. Yocgo, et al., J. Exp. Bot. 66 (2015) 137–146. doi: 10.1093/jxb/eru404
Y. Krasylenko, G. Komis, S. Hlynska, et al., Front. Plant Sci. 12 (2021) 675981.
M. Sedaghat, Y. Emam, A. Mokhtassi-Bidgoli, et al., Plants 10 (2021) 1223. doi: 10.3390/plants10061223
C. Screpanti, R. Fonné-Pfister, A. Lumbroso, et al., Bioorg. Med. Chem. Lett. 26 (2016) 2392–2400.
B. Zwanenburg, A.S. Mwakaboko, C. Kannan, Pest Manag. Sci. 72 (2016) 2016–2025. doi: 10.1002/ps.4222
H. Goossens, T.S.A. Heugebaert, B. Dereli, et al., Eur. J. Org. Chem. 2015 (2015) 1211–1217. doi: 10.1002/ejoc.201403457
A.S. Mwakaboko, B. Zwanenburg, European J. Org. Chem. 2016 (2016) 3495–3499. doi: 10.1002/ejoc.201600576
D. Blanco-Ania, J.J. Mateman, A. Hýlová, et al., Pest Manag. Sci. 75 (2019) 3113–3121. doi: 10.1002/ps.5553
J.G. Zorrilla, A. Cala, C. Rial, et al., J. Agric. Food Chem. 68 (2020) 9636–9645. doi: 10.1021/acs.jafc.0c02361
Y. Kang, Z. Pang, N. Xu, et al., J. Agric. Food Chem. 68 (2020) 11077–11087. doi: 10.1021/acs.jafc.9b08044
S.D. Fornier, A. de Saint Germain, P. Retailleau, et al., J. Nat. Prod. 85 (2022) 1976–1992. doi: 10.1021/acs.jnatprod.2c00282

Figure 1 (A) Small molecules that regulate auxin biosynthesis. (B) Small molecules regulating the polar transport of auxin. (C) Small molecules that regulate auxin signal transduction.

Figure 2 (A) Small molecules that regulate CTK biosynthesis. (B) Small molecules that regulate CTK signal transduction. (C) Structural analogues based on cytokinin skeleton modification.

Figure 3 (A) The regulatory role of small molecules in GA biosynthesis and signal pathways. (B) Structural analogues.

Figure 4 (A) Small molecule that regulates ABA biosynthesis. (B) Small molecule that regulates ABA signal transduction.

Figure 6 (A) Small molecules modulating the JA signaling pathway. (B) Small molecules involved in JA biosynthesis. (C) JA structural analogues.

Figure 7 (A) Small molecule regulating SL biosynthesis. (B) Small molecules modulating the SL signaling pathway. (C) SL analogues.

 扫一扫看文章
扫一扫看文章

扫一扫关注我们
 DownLoad:
DownLoad:
 下载:
下载:


 下载:
下载: