Table 1.
Procedures and extractants used for a sequential five-step chemical extraction.
Citation:
Jianju Li, Xinwei Chen, Yang Yu, Hao Ma, Xinhui Xia, Zixuan Zhao, Junqiu Jiang, Qingliang Zhao, Yingzi Lin, Liangliang Wei. Insights into bioavailable heavy metal impact driven by sludge application on soil nitrification: Toxicity thresholds and influential factors[J]. Chinese Chemical Letters,
2025, 36(7): 110410.
doi:
10.1016/j.cclet.2024.110410

Insights into bioavailable heavy metal impact driven by sludge application on soil nitrification: Toxicity thresholds and influential factors
English
Insights into bioavailable heavy metal impact driven by sludge application on soil nitrification: Toxicity thresholds and influential factors
Abstract:
Strict regulations on heavy metal (HM) limits impede the sludge land utilization for carbon emission reduction. This study aimed to evaluate the impact of bioavailable HMs (Cd, Cu, and Zn) on soil nitrification and determine toxicity thresholds via two cycles of sludge land application tests over 185 days. HMs inhibited gene abundance in their labile fractions, with the most affected being nitrite-oxidizing bacteria (NOB)-nxrB, followed by ammonia-oxidizing bacteria (AOB)-amoA, NOB-nxrA, and ammonia oxidizing archaea (AOA)-amoA. Toxicity thresholds for incremental labile fractions of HMs (in mg/kg) were determined as 0.35 for Cd, 21.73 for Cu, and 84.04 for Zn. Additionally, AOB, as the core nitrifiers, significantly correlated (P < 0.05) with ammonia nitrogen, soil organic matter, total phosphorus, and total potassium, playing a pivotal role in maintaining intricate interactions within HMs-spiked sludge-treated soil systems. The acute toxicity effects of HMs on potential ammonia oxidation (PAO), measured by inhibition rates, were 77.04%, 73.63%, and 67.06% for Cd, Cu, and Zn, with labile fractions contributing 33.79%, 40.19%, and 28.37%, respectively. Long-term sludge land application revealed chronic toxicity of HMs to PAO through the reshaping of ammonia-oxidizing microorganisms, particularly Cu and Zn. These findings provide insights into HM toxicity thresholds and their impact on nitrification, supporting sustainable sludge land management.
-
Key words:
- Labile heavy metals
- / Sludge land management
- / Nitrifiers
- / Ammonia oxidation capacity
- / Dynamic effects
-
The land application of digested (treated) sludge offers a meaningful route to recycle nutrients and reduce carbon emissions. However, it may also introduce unwanted contaminants such as heavy metals (HMs) into the soil [1]. The presence of HMs in treated sludge, with a total concentration of 0.5%–2% (dry weight) [2, 3], poses a significant threat to the soil ecological environment [4]. These HMs, known for their high toxicity and persistence, can cause irreversible damage to the abundance, activity, and diversity of soil microorganisms during long-term sludge application [5]. Hence, countries, especially China, have set strict limits on total metal content in treated sludge and led only 29.3% of sludge for land utilization.
It is estimated that the increasing of the contribution of land utilization to 50% in China would reduce as high as 14.0% of CO2-equivalent greenhouse gas emissions from sludge management in 2030 [6]. Consequently, there is a pressing need to establish precise strategies for HMs toxicity control in sludge land utilization. Although the total metal content in sludge serves as an indicator of contamination, the toxicity, bioavailability, and mobility of HMs are heavily dependent on their bioavailable forms rather than the total content [7-9]. Therefore, clarifying the toxicity threshold of bioavailable HMs on microbial communities in sludge land utilization would provide valuable recommendations for enhancing control standards.
Nitrification, a rate-limiting step in the microbially mediated N cycle, is primarily controlled by several functional genes. Ammonia-oxidizing archaea (AOA)-amoA and ammonia-oxidizing bacteria (AOB)-amoA play a primary role in the conversion of ammonium (NH4+-N) to nitrite (NO2−-N) [10]. Subsequently, nitrite-oxidizing bacteria (NOB) with genes nxrA and nxrB facilitate the transformation of NO2−-N into nitrate (NO3−-N) [11, 12]. This process is reported to be highly sensitive to the HM contamination [13], suggesting the existence of a toxicity threshold for nitrifiers. Extensive efforts have been focused on understanding the metal toxicity to nitrifiers and determining the thresholds for soil nitrification [14, 15]. This is primarily achieved through the assessment of nitrifiers' abundance and their potential ammonia oxidation (PAO) rate. The PAO rate is commonly recognized for its sensitivity and is employed as an indicator to assess the impacts of HMs stress on soil nitrogen transformation function [16, 17]. These efforts have revealed that the toxicity of HMs on nitrification varied widely across soils with different properties [18-20]. Filling this knowledge gap regarding the toxicity thresholds of bioavailable HMs in the context of sludge land application is crucial due to their direct impact on soil properties [21].
To identify toxicity thresholds, models like the threshold model, linear non-threshold model, and hormetic dose-response model have been widely employed via the detection of HMs concentration and dose responses of nitrifiers [22]. However, literatures revealed that the microorganisms' sensitivity, tolerance, or resistance to metal-contaminated soil varied significantly over different time scales [23-25]. Exogenous factors, such as extended aging time of HMs, can attenuate their toxicity on nitrification [20]. Endogenous factors, including variations in stress tolerances of nitrifiers and their differential soil niches over incubation time, have significant implications as well [26]. Thus, exploring the dynamics response of nitrifiers to cumulative sludge application with different HMs contamination and identifying the key factors influencing soil nitrification would contribute to a better understanding of the long-term impacts of digested sludge application on soil ecological functions.
Therefore, we opted to include three specific heavy metals (Cu and Zn, which are the highest in content according to Geng et al. [27], and Cd, known for its high toxicity according to Qin et al. [28]) from the list of controlled pollutants in sludge for agricultural use as outlined in "Control standards of pollutants in sludge for agriculture use (GB 4284–2018)" of China. These metals were then subjected to a 185-day greenhouse incubation, simulating two cycles of digested sludge land application. Specifically, three aspects were focused on: (1) Investigating the impact of Cd, Cu, and Zn derived from sludge on the nitrifiers and determining their bioavailable toxicity thresholds; (2) exploring the interplay and identifying core factors among soil properties, HM fractionations, and nitrification-related genes during consecutive sludge applications; and (3) investigating the dynamic response of PAO rate to different concentrations of Cd, Cu, and Zn, and identifying key contributing factors to this process. The findings could assist in refining HM concentration standards, promoting sustainable sludge management for resource recovery and carbon emission reduction within the framework of a circular economy.
Soil samples were collected from the surface layer (0–20 cm) of a clay-textured paddy field in Harbin, China, and their physicochemical properties are listed in Table S1 (Supporting information). Digested sludge used for experiments was acquired from a sludge treatment plant located in Dalian, China, with the corresponding properties for the sludge shown in Table S2 (Supporting information). The experimental preparation details were adapted from our previous work [29].
To simulate the addition of Cd, Cu, and Zn present in the sludge, air-dried digested sludge was applied to the soil at a rate of 7.5 tons per hectare. Subsequently, the soil was spiked with CdCl2·2.5H2O, CuCl2·2H2O, and ZnCl2 at concentrations 0.75, 1, 1.5, and 2 times the thresholds specified for grade B sludge (Table S3 in Supporting information). The different treatments were denoted by the specific types of HMs and their content relative to the permissible limit for grade B sludge. For example, Cd-0.75B, Cd-1B, Cd-1.5B, and Cd-2B represented treatments where Cd was present at concentrations ranging from 0.75 to 2 times the permissible limit for grade B sludge, respectively. Moreover, a control group was included in the study, where only digested sludge was applied to the soil.
Soil samples weighing 50 g were collected at various time intervals (1, 7, 14, 21, 35, 49, 63, 84, and 105 days), mixed thoroughly, and then employed for subsequent analysis. Once the NH4+-N was depleted, a secondary sludge application containing Cd, Cu, and Zn was performed using the same treatments as the initial sludge application. Subsamples of soil (50 g) were obtained on days 122, 136, 157, and 185, representing different time points during the second sludge land use, corresponding to 7, 21, 42, and 70 days, respectively.
Details of soil physicochemical analysis, including the measurement of soil pH, soil redox potential (Eh), NH4+-N, NO3–-N, soil organic matter (SOM), total phosphorus (TP), and total potassium (TK) are described in the Text S1 (Supporting information). HMs in the soil were fractionated using a sequential five-step chemical extraction method described by Tessier et al. [30], with the procedures and extractants listed in Table 1. This method divides the total metals into five fractions: exchangeable fraction (F1), carbonate-bound fraction (F2), Fe–Mn oxide bound fractions (F3), organic bound fraction (F4) and residual fraction (F5). Soil PAO rate was measured using the chlorate inhibition method according to Li et al. [31]. Quantitative polymerase chain reaction (qPCR) was performed to quantify the abundance of nitrifying functional marker genes: AOA-amoA, AOB-amoA, nxrA, and nxrB. Detailed information concerning DNA extraction, the qPCR assay mixture (Text S2 in Supporting information), primer sequences, and cycling parameters (Table S4 in Supporting information) is shown in Supporting information.
Table 1
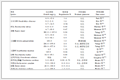
 DownLoad:
CSV
DownLoad:
CSV
Fractions
Extractants
Volume
(mL)Temp.
(℃)Time
(h)F1 1 mol/L MgCl2 (pH 7.0) 10 25 ± 1 1 F2 1 mol/L NaAc (pH 5.0) 10 25 ± 1 5 F3 0.04 mol/L NH2OH·HCl in 25% HAc 20 96 ± 3 6 F4 (a) 0.02 mol/L HNO3 and 30% H2O2 (a) 3 and 5 85 ± 2 (a) 2 (b) 30% H2O2 (b) 3 (b) 3 (c) 3.2 mol/L NH4Ac (c) 5 (c) 0.5 (d) distilled water (d) 4 (d) - F5 HNO3-HF-HClO4 5–3-2 190 0.5–2-1 The statistical analysis was performed using SPSS software (Version 25). Pearson's correlation test was employed to investigate the relationships between soil nitrifying microbe gene abundance and labile HM concentrations, as well as the relationship between HM fractionation concentrations and PAO inhibition rates. Multiple linear regression analyses were performed to evaluate the potential correlation between the log-transformed amoA gene abundances and PAO rates. The statistical analyses were conducted at significance levels of 0.05 and 0.01. Network analysis among soil properties, HM fractionation, and nitrification functional genes was employed by Spearman correlation analysis (correlation coefficient |r| > 0.5, P < 0.05) and visualized via Gephi software (v 0.10.1) [32]. Additionally, Random Forest analysis, relying on scikit-learn, was used to determine the relative importance of features [33, 34] in the suppression of PAO rate by Cd, Cu, and Zn in soil.
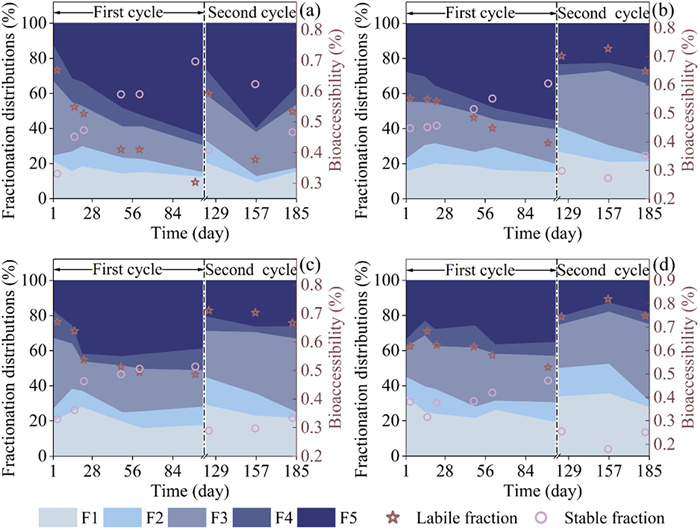
The toxicity of exogenous HMs to the soil microbial habitat primarily depends on their labile fractions (F1 + F2 + F3), which are influenced by the adsorption and diffusion processes with soil components [20, 35]. Therefore, fractional concentration distribution of HMs and their bioaccessibility in samples obtained from treatments involving the application of Cd, Cu, and Zn- spiked sludge was analyzed. The results, as illustrated in Fig. 1, Figs. S1 and S2 (Supporting information), present data obtained from samples collected throughout two application cycles.
Figure 1
 Figure 1. Variation in fractionation and bioavailability of Cd in soil over two cycles of land utilization with sludge amended at varying Cd concentrations: Cd-0.75B (a), Cd-1B (b), Cd-1.5B (c), Cd-2B (d). The scatter spots correspond to the right Y axis.
Figure 1. Variation in fractionation and bioavailability of Cd in soil over two cycles of land utilization with sludge amended at varying Cd concentrations: Cd-0.75B (a), Cd-1B (b), Cd-1.5B (c), Cd-2B (d). The scatter spots correspond to the right Y axis.For example, the bioavailability of Cd in treatments such as Cd-0.75B, Cd-1B and Cd-1.5B exhibited a decreasing trend with the transformation of the F3 fraction to the F5 fraction (Fig. 1). Eventually, F5 fraction became the dominant fraction in the soil at the end of the first sludge application, ranging from 38.91% to 64.52%. Regarding to the second sludge application, the stabilization of Cd was primarily attributed to the transformation of F2 to F3 fraction. The main fractionation of Cd was found to bind with iron and manganese oxides (ranging from 35.96% to 43.78%), consistent with findings reported by Liu et al. [36]. It is worth noting that at the end of the first and second sludge applications, the percentages of the F1 fraction for Cd were comparatively high, ranging from 12.26% to 19.10% and from 15.17% to 26.62%, respectively. These findings indicate a high bioavailability of Cd for the microbial habitat. Moreover, a similar consensus was reported by Yang et al. [37], suggesting that Cd exhibits weak non-specific adsorption at permanently charged sites on mineral surfaces [38]. Consequently, these findings underscore the high bioavailability of Cd in soil and its potential impact on microbial habitats.
During the two cycles of sludge application, a clear tendency of Cu immobilization was observed. The stable fraction of Cu in the soil ranged from 64.82% to 82.94% at the end of the first cycle, and from 49.52% to 76.35% at the end of the second cycle (Fig. S1). This suggests that specific adsorption plays a more significant role than nonspecific adsorption (i.e., cation exchange), indicating a strong bond between Cu and the soil matrix [39]. In terms of fractionation, the transformation of F2, F3, and F4 into the F5 fraction predominantly accounted for the occupation of the stable fraction following the initial application of digested sludge. Furthermore, in the course of the second sludge application, the F2 fraction transformed into both the F3 and F5 fractions. In relation to Zn, while there was stabilization following the first cycle of sludge application, involving a conversion from the F2 to the F5 fraction (Fig. S2), the labile fraction remained predominant (49.91%–60.85%). It is noteworthy that the labile fraction of Zn in the soil exhibited a considerable increase during the second cycle of sludge utilization. The proportion of the labile fraction, primarily composed of the F2 and F3 fraction [40], ranged from 76.0% to 83.5%. These observed changes in the labile fraction raise concerns about their potential impact on the nitrification in the soil.
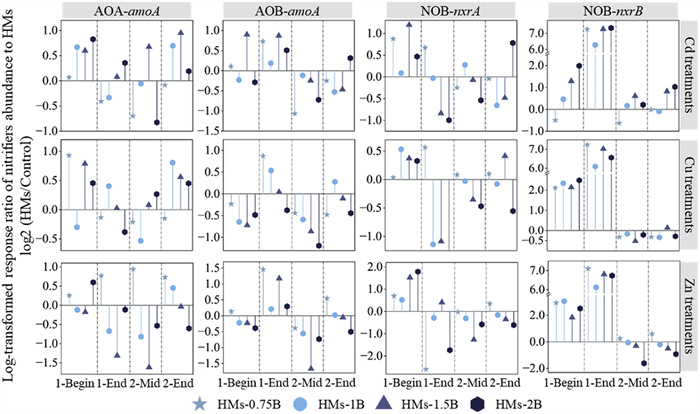
Quantitative PCR was employed to assess the impact of Cd-, Cu-, and Zn-spiked sludge application on the absolute abundance of nitrifying functional genes. The response ratios of their abundance to different levels of Cd, Cu, and Zn were calculated, and these ratios were normalized (log2 fold) for systematic assessment (Fig. 2). Overall, all four nitrification genes exhibited an increase in abundance following the initial cycle of sludge application (Fig. S3 in Supporting information), with nxrA exhibiting the highest order of magnitude increase (2.86), followed by AOB-amoA (1.70), nxrB (1.44), and AOA-amoA (0.69). Theoretically, the NH4+-N generated from sludge and the mineralization of organic N within the sludge (Fig. S4 in Supporting information) promote microbial growth [41]. However, most of these stimulatory effects gradually diminished following the second cycle of sludge application, except for nxrA (Fig. S3). Additionally, besides the impact of sludge addition, we also observed various response patterns exhibited by nitrifiers in soils spiked with Cd, Cu, and Zn to counteract metal toxicity. These response patterns varied depending on the exposure time, the specific types of HMs, and the degree of contamination.
Figure 2
 Figure 2. Log-transformed response ratios for the abundance of AOA-amoA, AOB-amoA, NOB-nxrA, and NOB-nxrB in soils amended with sludge spiked at varying Cd, Cu, and Zn concentrations over two cycles of land utilization. Values are for 1 and 105 days after the first cycle of sludge addition, as well as 42 and 70 days after the second cycle of sludge addition. Log transformation (Log2 fold) was employed to normalize and standardize the nitrifiers' response to HMs.
Figure 2. Log-transformed response ratios for the abundance of AOA-amoA, AOB-amoA, NOB-nxrA, and NOB-nxrB in soils amended with sludge spiked at varying Cd, Cu, and Zn concentrations over two cycles of land utilization. Values are for 1 and 105 days after the first cycle of sludge addition, as well as 42 and 70 days after the second cycle of sludge addition. Log transformation (Log2 fold) was employed to normalize and standardize the nitrifiers' response to HMs.On the first day of sludge application, the addition of HMs stimulated the growth of AOA and NOB. Such promotion was also observed in soil amended with cow manure under moderate metal contamination conditions [42]. In contrast, AOB were mostly inhibited by metals, particularly in Cu-spiked sludge-amended soil, indicating its greater sensitivity to Cu than AOA, which is consistent with a previous study [16]. As the incubation period progressed, the effect of HMs on nitrifiers varied widely. Initially, Cd and Cu stimulated nxrA expression while Cu inhibited AOB; this effect converted to a hormesis effect by the 105th day of the first sludge application cycle. For instance, the abundance of nxrA increased under a low dose of Cd (Cd-0.75B) but declined under higher Cd doses (Cd-1.5B, Cd-2B), demonstrating that extended exposure led to the occurrence of hormetic responses at low Cd dose [43]. Additionally, Cu treatments, with a broader promoting range from Cu-0.75B to Cu-1.5B, increased the abundance of AOB, whereas Cu-2B treatment exerted toxic effects. The hormetic responses of Cu on AOB were also observed in the study by Gong et al., suggesting that the hormesis effect might be due to Cu acting as an active center for some nitrifying enzymes [44]. Besides the hormesis pattern, promotion also occurred. Specifically, nxrB was consistently enriched under treatments with Cd, Cu, and Zn, even at concentrations twice the permissible limit for grade B sludge, with log-transformed response ratios of 7.32, 6.56, and 6.73 for Cd, Cu, and Zn, respectively. Moreover, the growth of AOB-amoA was promoted under the treatments with Cd and Zn, regardless of the metal concentrations in the sludge, with log-transformed response ratio of 0.51 and 0.29 under Cd-2B and Zn-2B conditions.
During the middle period (157th day) of the second cycle of sludge application, a comparable hormesis response was noted for nxrB under Zn treatments, as well as nxrA under Cu treatments. Notably, there was a noticeable decrease in the abundance of AOB-amoA in treatments involving Cd, Cu, and Zn, as well as nxrA under Zn stress and nxrB under Cu stress. This decline was attributed to the cumulative impact of repeated sludge applications, resulting in elevated HM levels that exerted increased toxicity on nitrification microorganisms. By the end (185th day) of the second sludge amendment, the response of nitrifiers to metal pressure displayed distinct patterns depending on the types of HM. It is interesting to note that a high dose of Cd (Cd-2B) led to an observable increase in the abundance of all nitrifies, contrary to the conventional expectation that higher levels of Cd addition would inhibit their growth [25]. In contrast, the response of nitrifiers to Cu-2B treatment revealed that the cumulative Cu from two cycles of sludge applications was effective in suppressing the proliferation of AOB and NOB, except for AOA. This observation is consistent with previous research that suggests AOA is not as responsive to Cu addition because their archaeal membranes are less permeable to ions, which can be attributed to their unique membrane lipid structure [13, 16]. In addition, all soil nitrifiers exhibited a hormesis effect in response to Zn treatments. This hormesis effect in AOMs due to Zn has been reported by several previous studies [45]. Specifically, the promotion effect of low concentrations of Zn can be attributed to two primary reasons. Firstly, Zn2+ could replace Cu2+ in the ammonia monooxygenase (AMO) active site, reducing its activity and necessitating increased amoA expression for AMO synthesis [46]. Secondly, low concentrations of Zn enhance RNA polymerase activity, thereby improving gene transcription [47]. Conversely, the decrease in amoA gene expression at high concentrations of Zn is attributed to the inhibition of AOMs' energy acquisition pathways [48]. The varied response patterns to HMs at different exposure times and contamination levels highlight the critical importance of investigating toxicity thresholds for individual HMs over the two cycles of sludge land applications.
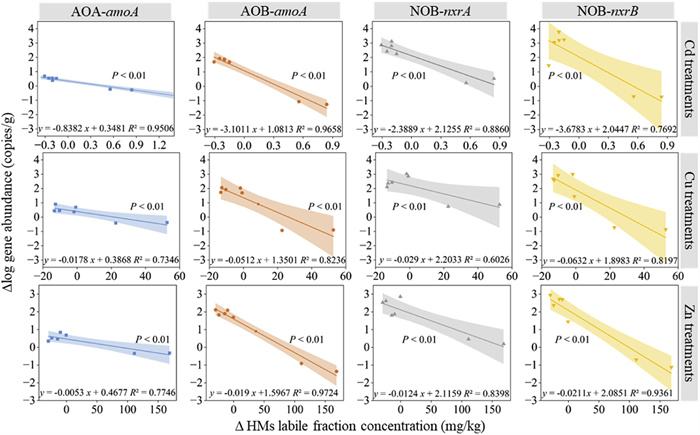
It is essential to accurately assess and monitor soil disturbances caused by the contamination of HMs by determining the toxicity threshold of nitrification microorganisms. This is particularly crucial when considering the prolonged impact of recurrent land use for treated sludge. Consequently, the toxicity threshold was explored by examining the correlation between the abundance of nitrifiers and concentration of labile fractions of Cd, Cu, and Zn (Fig. 3). In general, the concentration of the incremental labile (∆labile) fraction for metals had a negative correlation with the incremental log-transformed (∆log) abundances of AOA, AOB, and NOB. This indicates that raising the bioaccessible concentration of Cd, Cu, and Zn suppresses the proliferation of nitrifiers. To be specific, the most pronounced level of toxicity was indicated by the highest slope values in the Cd treatment, while Cu and Zn exhibited comparatively lower levels of toxicity.
Figure 3
 Figure 3. Correlation between soil nitrifying microbe gene abundance and the labile Cd, Cu, and Zn during the whole sludge application period. Nitrifiers response was assessed by comparing the change in log abundance from beginning and end of the initial sludge application, and between the end of the two cycles of sludge application.
Figure 3. Correlation between soil nitrifying microbe gene abundance and the labile Cd, Cu, and Zn during the whole sludge application period. Nitrifiers response was assessed by comparing the change in log abundance from beginning and end of the initial sludge application, and between the end of the two cycles of sludge application.Upon analyzing various nitrifying microorganisms, it was noted that NOB-nxrB exhibited the highest slope value when simulating the relationship between its log abundance variations and the changes in the labile fraction of the three metals. Therefore, NOB-nxrB was the most sensitive nitrifiers to the labile fraction of HMs, followed by AOB-amoA, NOB-nxrA, and AOA-amoA. The thresholds for Δlabile HMs that reduced nitrifier populations were 0.35 mg/kg for Cd, 21.73 mg/kg for Cu, and 84.04 mg/kg for Zn. These values were determined as the minimum concentrations that caused a reduction in the abundance of the four nitrifying functional genes to ensure the safe utilization of sludge in managing metal toxicity.
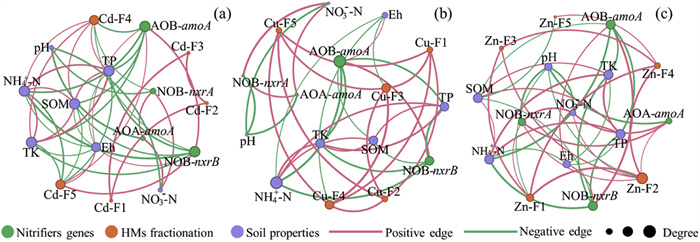
Previous studies have discovered that the microbial community in long-term polluted soils is predominantly shaped by the joint interactions of soil properties and HMs [32, 49]. Therefore, network analysis was conducted to illustrate interaction patterns among soil properties, HM fractions, and nitrification functional genes within the soil nitrification system, as well as to identify influential factors. Generally, network interactions were more complex in Cd-spiked sludge treatments compared to Cu and Zn treatments, characterized by a greater number of edges, higher average degree, and increased average clustering coefficient (Fig. 4 and Table S5 in Supporting information). This increased complexity is attributable to the higher participation of different Cd fractionations, which show more connections with soil properties and nitrifiers abundance.
Figure 4
 Figure 4. Network analysis among soil properties, fractional concentrations of Cd (a), Cu (b), Zn (c), and nitrifiers abundance in sludge-amended soils based on Spearman correlation analysis (|r| > 0.5, P < 0.05).
Figure 4. Network analysis among soil properties, fractional concentrations of Cd (a), Cu (b), Zn (c), and nitrifiers abundance in sludge-amended soils based on Spearman correlation analysis (|r| > 0.5, P < 0.05).The consecutive application of sludge to soil had a drastic impact on soil properties, including pH, Eh, and nutrients content [50, 51], which would inevitably influence soil microbial community structures [52]. Nutrients such as NH4+-N, SOM, TP, and TK were identified as the core soil properties in the network of Cd and Zn treatments. In contrast, NH4+-N was the most influential factor in the Cu treatments. It is noteworthy that NH4+-N, SOM, TP, and TK had a significant (P < 0.05) impact on AOB-amoA and NOB-nxrB for all HMs treated sludge amendments. On the other hand, NO3--N and pH were found to have a significant (P < 0.05) correlation with AOA-amoA and NOB-nxrA. Similarly, as indicated by Xing et al. [53], water content, NH4+-N, SOC, TN, and available phosphorus were found to potentially contribute to the distribution of AOB, and the community of AOA was correlated with NH4+-N and water content. Although pH and Eh were not core components in the soil nitrification system network, it is important to note that they were crucial factors that influenced the distribution of HM speciation [54]. Strong associations were observed between pH and both Zn-F1 and Zn-F2 fractions, while Eh significantly influenced Cd-F5 and Cu-F3 fractions (P < 0.05). Notably, it was observed that speciation of HM affected the growth of nitrifying microorganisms, especially in Zn treatments. For example, Zn-F4 and Zn-F5 presented positive correlations with AOB-amoA (P < 0.05), while Zn-F1 and Zn-F2 demonstrated positive correlations with NOB-nxrA (P < 0.05). These results suggest that HM speciation and soil properties jointly regulate nitrifiers [15]. Although the distribution of nitrifiers displayed dynamic variations across different HMs-spiked sludge treatments, the core nitrification functional genes (AOB and NOB-nxrB) remained consistent for Cd and Zn treatments. Moreover, AOB was also the core microbiome in Cu treatments, indicating its crucial role in maintaining the complex interactions within sludge-amended soil systems.
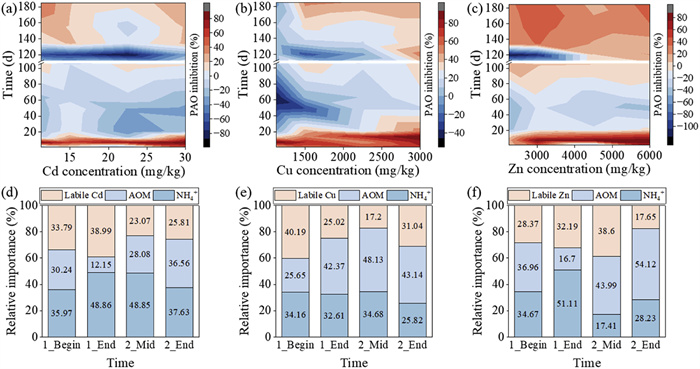
To uncover the dynamic response of PAO in soil to the application of sludge spiked with Cd, Cu, and Zn, we conducted an evaluation of PAO inhibition rates. The results are shown in Figs. 5a–c. It is evident that the inhibition patterns of PAO rate in response to Cd, Cu, and Zn were time-dependent. To be specific, acute toxicity was observed on the 7th day, regardless of the types and concentration levels of HMs. The average inhibition rates were 77.04% for Cd, 73.63% for Cu, and 67.06% for Zn, respectively. As the incubation period extended to the 49th day, there was a partial alleviation of inhibition, with concomitant increases in PAO rate noted in certain instances. However, contrary trends were observed towards the end of the first sludge amendment (105th day), with the PAO rate in soils treated with Cd (except for Cd-1B), all Cu treatments, and all Zn treatments being suppressed slightly.
Figure 5
 Figure 5. PAO inhibition rates and feature importance variation in soils for different Cd- (a, d), Cu- (b, e), and Zn- (c, f) spiked sludge application conditions.
Figure 5. PAO inhibition rates and feature importance variation in soils for different Cd- (a, d), Cu- (b, e), and Zn- (c, f) spiked sludge application conditions.In contrast to the first cycle of sludge application, PAO rate exhibited stimulation at the beginning (7th day) of the second cycle in all Cd treatments, as well as in the Cu-1B and Cu-1.25B treatments. Additionally, a hormesis response was noted under the treatments of Zn, with stimulation at lower concentrations (Zn-0.75B, Zn-1B) and inhibition at higher concentrations (Zn-1.25B, Zn-2B). Moreover, PAO was generally suppressed by the additions of Cd, Cu, and Zn at the middle (157th day) and the end (185th day) of the second cycle of sludge application. The average inhibition rates at the 185th day were 25.37%, 46.99%, and 32.09% for Cd, Cu and Zn, respectively, higher than those at the 157th day (21.11%, 23.83%, and 30.83%, respectively). In summary, acute toxicity towards PAO was observed at the beginning of the first sludge amendment, while chronic toxicity occurred at the middle and the end of the second sludge amendment in soils spiked with these three HMs. The temporal response pattern of PAO rate in this study contrasts with findings from previous research [55, 56], which reported acute toxicity on PAO, with subsequent recovery observed 20 days after initial inhibition in soils amended with Cu concentrations of 50–3200 mg/kg [55]. This comparison highlights the necessity for a more comprehensive understanding of the underlying mechanisms driving these time-dependent variations. To address this knowledge gap, NH4+-N, AOMs, and labile fractions of HMs were selected in combination with the previous findings [23, 25] to conduct a dynamic contribution analysis on the inhibition rate of PAO using a Random Forest model (Figs. 5d–f).
At the onset of the initial sludge application, it was evident that there was an immediate toxicity of labile Cu on soil PAO rate, as it accounted for a substantial contribution of 40.19%. Subsequently, the stabilization of HM was beneficial in alleviating the inhibition of ammonia oxidation [20, 25] with the contribution decreasing to 25.02%. However, a different trend was observed with Cd and Zn, where the contribution of their labile fraction to PAO inhibition rate increased during the first cycle of sludge utilization. This emphasizes the importance of further investigation into which specific fraction of HMs has a significant impact on PAO rate. This question is addressed in the below section.
In the course of the second sludge amendment, the factors influencing PAO inhibition rate exhibited temporary stability. For Cd treatments, NH4+-N was the restrictive factor for PAO inhibition rate, as demonstrated by Zhao et al. [25], indicating that the presence of NH4+-N significantly affects the impact of Cd on the nitrification process. Notably, AOMs were the key contributors to the variations in PAO inhibition rate observed in the Cu and Zn treatments, with the respective mean contributions of 45.64% and 49.06%. This suggests that the initial exposure to Cu- and Zn-spiked sludge may have reshaped the community of AOMs and potentially had a strong impact on the nitrification process during the second cycle of sludge application.
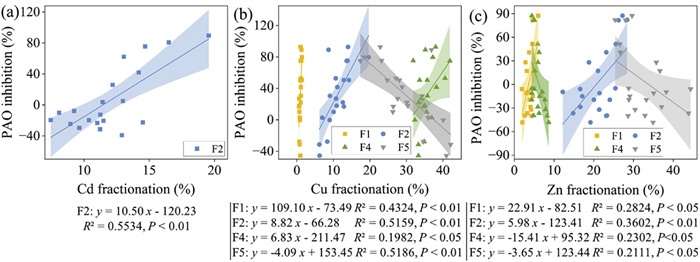
Clarifying the effects of HM fractionations on PAO rate is of vital importance for understanding the potential mechanisms behind the observed variations of PAO inhibition rate in soil. Thus, a linear regression analysis was conducted to evaluate the relationship between HM fractions and PAO inhibition rates (Fig. 6). In general, the F1 fraction concentration showed a positive correlation with PAO inhibition rate in soil treated with Cu- and Zn-spiked sludge. This implies that an increase in the proportion of the F1 fraction leads to a more serious PAO inhibition. This finding is consistent with Cela and Sumner [14, 56], who reported that the F1 fraction was strongly and negatively correlated with the nitrification inhibition. Similarly, the F2 fraction concentration of Cd, Cu and Zn exhibited a positive and significant correlation (P < 0.01) with the PAO inhibition, as observed in our previous research [29]. This indicates that carbonate-bound fraction of these three HMs is a reliable indicator of their inhibitory effects on nitrification in this study. According to the slope assessment, the inhibitory effect on PAO resulting from the increase in the F2 fraction decreased in the following order: Cd > Cu > Zn. Notably, the correlation between the F4 fraction of Cu and Zn and PAO inhibition was opposite. Specifically, an increase in the organic-bound fraction of Cu significantly (P < 0.05) inhibited the PAO rate in soil, whereas an increase in the organic-bound fraction of Zn significantly (P < 0.05) reduced PAO inhibition. In light of the non-toxic residual fraction and the observed correlations between HM fraction and PAO inhibition, it can be confidently stated that increasing the F5 fraction of Cu and Zn has a beneficial effect in reducing PAO inhibition.
Figure 6
 Figure 6. Relationship between fractionation concentration of Cd (a), Cu (b), Zn (c) and inhibition on PAO rates in soils during sludge land application.
Figure 6. Relationship between fractionation concentration of Cd (a), Cu (b), Zn (c) and inhibition on PAO rates in soils during sludge land application.To evaluate the predominant contribution of ammonia oxidizers to PAO rates, multiple linear regression analysis was conducted. Notably, a consistent positive correlation was observed between PAO rates variation and AOB abundance variation under all stress conditions of Cd, Cu, and Zn (P < 0.01, Table S6 in Supporting information). AOB, characterized by their high abundance and identified as the core nitrifiers in all treatments in this study, were found to significantly influence ammonia oxidation activity. Similarly, Mertens et al. [57] found that AOB community drove recovery of nitrification from Zn contamination in this soil. In contrast, AOA were negatively correlated with PAO rates in soil amended with Cu and Zn spiked sludge. These findings suggest that AOA and AOB may occupy different soil niches and employ different ammonia oxidation pathways [26, 58], possibly influenced by environmental drivers and HM stress [59]. The hypothesis is supported by the results of the network analysis (Fig. 4), which indicate that AOB and AOA were influenced by different soil properties, with AOB exhibiting higher susceptibility to HM speciation compared to AOA. This suggests that AOB primarily drive PAO during the application of Cd, Cu, and Zn-spiked sludge to land.
In conclusion, during the continuous two cycles of sludge land application, the presence of HMs in their labile fraction inhibited the abundance of nitrifiers, with NOB-nxrB experiencing the highest inhibition, followed by AOB-amoA, NOB-nxrA, and AOA-amoA. Furthermore, the rise of the labile fraction was strongly associated with the toxic effects of HMs, with threshold values of 0.35 mg/kg for Cd, 21.73 mg/kg for Cu, and 84.04 mg/kg for Zn. It is important to note that NH4+-N, SOM, TP, and TK had a significant (P < 0.05) impact on AOB and NOB-nxrB. On the other hand, NO3--N and pH were found to have a significant (P < 0.05) correlation with AOA and NOB-nxrA. Furthermore, AOB were identified as the core nitrifying microorganisms, essential for maintaining soil ecosystem functions amidst Cd, Cu, and Zn pollution and amendments. All three HMs exhibited both acute and chronic toxicity to PAO, with acute toxicity more pronounced. Acute toxicity, represented by the mean inhibition rate of PAO, was 77.04% for Cd, 73.63% for Cu, and 67.06% for Zn, whereas chronic toxicity levels were 25.37%, 46.99%, and 32.09%, respectively. In terms of the contribution of key factors, the labile fractions accounted for 33.79%, 40.19%, and 28.37% of the acute toxicity to PAO for Cd, Cu, and Zn, respectively. The chronic toxicity of Cu and Zn to PAO was closely related to the reshaping of the AOMs. In addition, NH4+-N was the primary factor restricting chronic toxicity to PAO in the Cd treatments. The inhibition of PAO rates showed a positive correlation with the F2 fraction concentration of HMs, with AOB being the main driver. These findings are crucial in refining HM concentration standards for sustainable sludge management, especially for determining the thresholds of labile Cd, Cu, and Zn.
CRediT authorship contribution statement
Jianju Li: Writing – review & editing, Writing – original draft, Methodology, Formal analysis, Data curation. Xinwei Chen: Writing – original draft, Methodology, Data curation. Yang Yu: Writing – original draft, Visualization. Hao Ma: Writing – review & editing, Formal analysis. Xinhui Xia: Methodology, Formal analysis, Data curation. Zixuan Zhao: Formal analysis, Data curation. Junqiu Jiang: Writing – review & editing, Writing – original draft. Qingliang Zhao: Resources, Conceptualization. Yingzi Lin: Funding acquisition, Resources. Liangliang Wei: Resources, Funding acquisition, Conceptualization.
Acknowledgments
This work received support from the National Key Research and Development Program of China (No. 2023YFC3207404) and the National Natural Science Foundation of China (No. 52270034). Additional funding was provided by the National Engineering Research Center for Safe Sludge Disposal and Resource Recovery (No. 2021A004) and the National Engineering Research Center for Bioenergy (No. 2021B007).
Supplementary materials
Supplementary material associated with this article can be found, in the online version, at doi:
10.1016/j.cclet.2024.110410 .
-
-
[1]
X. Li, S. Yuan, C. Cai, et al., Environ. Pollut. 341 (2024) 122907.
-
[2]
A. Pathak, M.G. Dastidar, T.R. Sreekrishnan, J. Environ. Manag. 90 (2009) 2343–2353.
-
[3]
K. Qin, L. Wei, J. Li, et al., Chin. Chem. Lett. 31 (2020) 2603–2613.
-
[4]
J. Zhang, Z. Liu, B. Tian, et al., J. Hazard. Mater. 441 (2023) 129891.
-
[5]
V. Kapoor, X. Li, M. Elk, et al., Environ. Sci. Technol. 49 (2015) 13454–13462. doi: 10.1021/acs.est.5b02748
-
[6]
L. Wei, F. Zhu, Q. Li, et al., Environ. Int. 144 (2020) 106093.
-
[7]
H. Yesil, R. Molaey, B. Calli, A.E. Tugtas, Water Res. 201 (2021) 117303.
-
[8]
Y. Xu, Y. Feng, Pol. J. Environ. Stud. 25 (2016) 405–412. doi: 10.15244/pjoes/60861
-
[9]
J. Jia, J. Bai, R. Xiao, et al., Sci. Total Environ. 807 (2022) 151725.
-
[10]
L. Li, Y. Ling, H. Wang, et al., Chin. Chem. Lett. 31 (2020) 28–38. doi: 10.5703/educationculture.36.1.0028
-
[11]
M. Kuypers, H.K. Marchant, B. Kartal, Nat. Rev. Microbiol. 16 (2018) 263–276. doi: 10.1038/nrmicro.2018.9
-
[12]
B. Xi, H. Yu, Y. Li, et al., J. Hazard. Mater. 403 (2021) 123853.
-
[13]
S. Tang, Y. Rao, S. Huang, et al., J. Environ. Manag. 326 (2023) 116641.
-
[14]
S. Cela, M.E. Sumner, Commun. Soil Sci. Plant Anal. 33 (2002) 19–30.
-
[15]
L. Lu, C. Chen, T. Ke, et al., Sci. Total Environ. 830 (2022) 154732.
-
[16]
H. He, H. Liu, T. Shen, et al., Geoderma. 321 (2018) 141–150.
-
[17]
Y. Wang, X. Zeng, Y. Zhang, et al. J. Environ. Sci. 127 (2023) 15–29.
-
[18]
J. Li, Y. Huang, Y. Hu, et al., J. Environ. Sci. 44 (2016) 131–140. doi: 10.26599/jgse.2016.9280016
-
[19]
C. Yanez, J. Verdejo, H. Moya, et al., Chemosphere. 300 (2022) 134517.
-
[20]
X. Zhang, X. Zhang, L. Li, et al., Environ. Res. 204 (2022) 111941.
-
[21]
R. Dhanker, S. Chaudhary, S. Goyal, V.K. Garg, Environ. Technol. Innov. 23 (2021) 101642.
-
[22]
M.Z. Hashmi, Naveedullah, H. Shen, et al., Environ. Int. 64 (2014) 28–39.
-
[23]
S. Ruyters, J. Mertens, D. Springael, E. Smolders, Soil Biol. Biochem. 42 (2010) 766–772.
-
[24]
J. Song, Q. Shen, L. Wang, et al., Environ. Pollut. 243 (2018) 510–518. doi: 10.3390/f9090510
-
[25]
H. Zhao, J. Lin, X. Wang, et al., Environ. Sci. Technol. 55 (2021) 14305–14315. doi: 10.1021/acs.est.1c04409
-
[26]
A.E. Taylor, L.H. Zeglin, T.A. Wanzek, D.D. Myrold, P.J. Bottomley, ISME J. 6 (2012) 2024–2032. doi: 10.1038/ismej.2012.51
-
[27]
Y. Geng, C. Zhang, Y. Zhang, et al., Sci. Pollut. Res. 28 (2021) 29146–29156. doi: 10.1007/s11356-021-12762-8
-
[28]
S. Qin, H. Liu, Z. Nie, et al., Pedosphere. 30 (2020) 168–180.
-
[29]
J. Li, H. Ma, H. Yu, et al., J. Hazard. Mater. 466 (2024) 133552.
-
[30]
A. Tessier, P.G.C. Campbell, M. Bisson, Anal. Chem. 51 (1979) 844–851. doi: 10.1021/ac50043a017
-
[31]
J. Li, H. Yang, K. Qin, et al., J. Hazard. Mater. 423 (2022) 126994.
-
[32]
Z. Cheng, J. Shi, Y. He, L. Wu, J. Xu, J. Hazard. Mater. 426 (2022) 128095.
-
[33]
S. Cipullo, B. Snapir, G. Prpich, P. Campo, F. Coulon, Chemosphere 215 (2019) 388–395.
-
[34]
T. Zeng, Y. Liang, Q. Dai, et al., Chin. Chem. Lett. 33 (2022) 5184–5188.
-
[35]
B. Wang, B. Huang, Y.B. Qi, W.Y. Hu, W.X. Sun, Chin. Chem. Lett. 23 (2012) 1287–1290.
-
[36]
G. Liu, Z. Yu, X. Liu, et al., J. Chem. 2020 (2020) 1–10.
-
[37]
J. Yang, Z. Guo, L. Jiang, et al., Ecotoxicol. Environ. Saf. 239 (2022) 113617.
-
[38]
G. Xueyuan, L.J. Evans, S.J. Barabash, Geochim. Cosmochim. Acta 74 (2010) 5718–5728.
-
[39]
H.B. Bradl, J. Colloid Interface Sci. 277 (2004) 1–18.
-
[40]
L. Mouni, L. Belkhiri, A. Bouzaza, J. Bollinger, Environ. Earth Sci. 75 (2016) 1–8.
-
[41]
H. Liu, Waste Manag. 56 (2016) 575–583.
-
[42]
M.M. Ali, A. Khanom, K. Nahar, et al., Environ. Contam. Toxicol. 106 (2021) 707–713. doi: 10.1007/s00128-021-03112-y
-
[43]
D. Fan, S. Wang, Y. Guo, et al., Sci. Total Environ. 757 (2021) 143771.
-
[44]
P. Gong, S.D. Siciliano, S. Srivastava, C.W. Greer, G.I. Sunahara, Hum. Ecol. Risk Assess. 8 (2002) 1067–1081. doi: 10.1080/1080-700291905828
-
[45]
Q. Wu, K. Huang, H. Sun, et al., J. Hazard. Mater. 343 (2018) 166–175.
-
[46]
T.S. Radniecki, R.L. Ely, Biotechnol. Bioeng. 99 (2008) 1085–1095. doi: 10.1002/bit.21672
-
[47]
J.E. Coleman, Biochem. Biophys. Res. Commun. 60 (1974) 641.
-
[48]
T.S. Radniecki, L. Semprini, M.E. Dolan, Biotechnol. Bioeng. 104 (2009) 1004–1011. doi: 10.1002/bit.22454
-
[49]
X. Zhao, J. Huang, J. Lu, Y. Sun, Ecotoxicol. Environ. Saf. 170 (2019) 218–226.
-
[50]
W. Longhua, C. Miaomiao, L. Zhu, et al., J. Soils Sediments 12 (2012) 531–541.
-
[51]
C.D.N.A. Tsadilas, T. Matsi, N. Barbayiannis, D. Dimoyiannis, Commun. Soil Sci. Plant Anal. 26 (1995) 2603–2619. doi: 10.1080/00103629509369471
-
[52]
A. Charlton, R. Sakrabani, S. Tyrrel, C.M. Rivas, S.P. McGrath, et al., Environ. Pollut. 219 (2016) 1021–1035.
-
[53]
Y. Xing, Y.X. Si, C. Hong, Y. Li, Arch. Environ. Contam. Toxicol. 69 (2015) 20–31. doi: 10.1007/s00244-015-0144-9
-
[54]
B. Shen, X. Wang, Y. Zhang, et al., Sci. Total Environ. 711 (2020) 135229.
-
[55]
J. Li, Y. Zheng, Y. Liu, et al., Microb. Ecol. 67 (2014) 931–941. doi: 10.1007/s00248-014-0391-8
-
[56]
S. Cela, M.E. Sumner, Water Air Soil Pollut. 141 (2002) 91–104.
-
[57]
J. Mertens, K. Broos, S.A. Wakelin, et al., ISME J. 3 (2009) 916–923. doi: 10.1038/ismej.2009.39
-
[58]
W. Qin, S.P. Wei, Y. Zheng, et al., Nat. Microbiol. 9 (2024) 524–536. doi: 10.1038/s41564-023-01593-7
-
[59]
A. Ginawi, L. Wang, H. Wang, B. Yu, Y. Yunjun, PeerJ 8 (2020) e8256. doi: 10.7717/peerj.8256
-
[1]
-
Figure 1 Variation in fractionation and bioavailability of Cd in soil over two cycles of land utilization with sludge amended at varying Cd concentrations: Cd-0.75B (a), Cd-1B (b), Cd-1.5B (c), Cd-2B (d). The scatter spots correspond to the right Y axis.
Figure 2 Log-transformed response ratios for the abundance of AOA-amoA, AOB-amoA, NOB-nxrA, and NOB-nxrB in soils amended with sludge spiked at varying Cd, Cu, and Zn concentrations over two cycles of land utilization. Values are for 1 and 105 days after the first cycle of sludge addition, as well as 42 and 70 days after the second cycle of sludge addition. Log transformation (Log2 fold) was employed to normalize and standardize the nitrifiers' response to HMs.
Figure 3 Correlation between soil nitrifying microbe gene abundance and the labile Cd, Cu, and Zn during the whole sludge application period. Nitrifiers response was assessed by comparing the change in log abundance from beginning and end of the initial sludge application, and between the end of the two cycles of sludge application.
Figure 4 Network analysis among soil properties, fractional concentrations of Cd (a), Cu (b), Zn (c), and nitrifiers abundance in sludge-amended soils based on Spearman correlation analysis (|r| > 0.5, P < 0.05).
Figure 5 PAO inhibition rates and feature importance variation in soils for different Cd- (a, d), Cu- (b, e), and Zn- (c, f) spiked sludge application conditions.
Figure 6 Relationship between fractionation concentration of Cd (a), Cu (b), Zn (c) and inhibition on PAO rates in soils during sludge land application.
Table 1. Procedures and extractants used for a sequential five-step chemical extraction.
Fractions
Extractants
Volume
(mL)Temp.
(℃)Time
(h)F1 1 mol/L MgCl2 (pH 7.0) 10 25 ± 1 1 F2 1 mol/L NaAc (pH 5.0) 10 25 ± 1 5 F3 0.04 mol/L NH2OH·HCl in 25% HAc 20 96 ± 3 6 F4 (a) 0.02 mol/L HNO3 and 30% H2O2 (a) 3 and 5 85 ± 2 (a) 2 (b) 30% H2O2 (b) 3 (b) 3 (c) 3.2 mol/L NH4Ac (c) 5 (c) 0.5 (d) distilled water (d) 4 (d) - F5 HNO3-HF-HClO4 5–3-2 190 0.5–2-1  下载: 导出CSV
下载: 导出CSV
-







 扫一扫看文章
扫一扫看文章
计量
- PDF下载量: 0
- 文章访问数: 34
- HTML全文浏览量: 3
 下载:
下载:
