Citation:
Cheng Wang, Ji Wang, Dong Liu, Zhi-Ling Zhang. Advances in virus-host interaction research based on microfluidic platforms[J]. Chinese Chemical Letters,
2024, 35(12): 110302.
doi:
10.1016/j.cclet.2024.110302
Advances in virus-host interaction research based on microfluidic platforms
English
Advances in virus-host interaction research based on microfluidic platforms
School of Life Sciences, Co-Innovation Center of Neuroregeneration, Nantong Laboratory of Development and Diseases, Nantong University, Nantong 226019, China
b.
College of Chemistry and Molecular Sciences, Wuhan University, Wuhan 430072, China
c.
Centre for Immunology and Infection, Hong Kong Science and Technology Park, Hong Kong SAR 999077, China
* Corresponding authors. E-mail addresses: tom@ntu.edu.cn (D. Liu)
zlzhang@whu.edu.cn (Z.-L. Zhang). 1 These authors contributed equally to this work.
Received Date:
16 March 2024 Accepted Date:
29 July 2024 Revised Date:
28 June 2024 Available Online:
15 December 2024
Abstract:
Viral epidemics pose a serious threat to global public health, making it essential to explore virus-host interactions for uncovering the pathogenesis of viral diseases and developing effective antiviral strategies. Traditional in vitro cell infection models struggle to replicate physiological microenvironment, while animal infection models may encounter obstacles such as species gap, high-cost, and ethical issues. Additionally, potential heterogeneous infection outcomes are usually inaccessible by population-based experiments. Microfluidics, as an emerging interdisciplinary platform, has proven to be a powerful tool for inquiring virus-host interactions. In this review, conventional virological methods were introduced first and remarkable advantages of microfluidics in viral cell biology were highlighted. Next, the in-depth applications of microfluidics in analyzing heterogeneity of virus-host interplays, dynamic monitoring of events related to viral life cycle, and modeling of viral infectious diseases were fully elaborated from the perspective of single-cell chip, multi-cell culture chip and organ-on-a-chip (organ chip). Finally, the opportunities and challenges in developing robust microfluidic methods for virology were discussed. Overall, this review aims to provide an overview of microfluidic-based research on virus-host interaction and promote multidisciplinary collaborations for better understanding and responding to viral threats.
Viruses are highly prevalent in nature and lurk in our surroundings, posing an urgent threat to public health due to their high pathogenicity and contagiousness. Over the past decades, numerous serious viral epidemics such as influenza viruses, Ebola virus (EBOV), Zika virus (ZIKA) and norovirus emerging have broken out around the world, with some of them even resulted in periodic pandemics [1-3]. Meanwhile, the emergence of novel virus strains, including severe acute respiratory syndrome (SARS), Middle East respiratory syndrome coronavirus (MERS-CoV), and acute respiratory syndrome coronavirus 2 (SARS-CoV-2) caused severe manifestations in human beings [4]. Particularly, the COVID-19 pandemic caused by SARS-CoV-2 infection in 2019 has brought heavily negative impacts on global society and economy. Facing to the persistent challenges leveraging with emerging and reemerging viruses to human health, there is an urgent need to seek reliable strategies for effective prevention, rapid diagnosis and precise treatment of viral diseases.
Understanding of the mechanisms underlying virus-host interactions is crucial for the development of antiviral strategies. In vitro cell infection models and in vivo animal infection models are commonly used in virology research to gain insights into viral pathogenesis, immune responses, and the evaluation of antiviral drugs and vaccines [5,6]. However, these models have inherent limitations, such as the absence of extracellular environments, unavoidable interspecific differences, and the neglected viral and cellular heterogeneity, making them fail to reproduce realistic human viral infections [7,8]. Therefore, developing novel platforms capable to overcome these limitations will create opportunities for innovative findings of virus-host interactions.
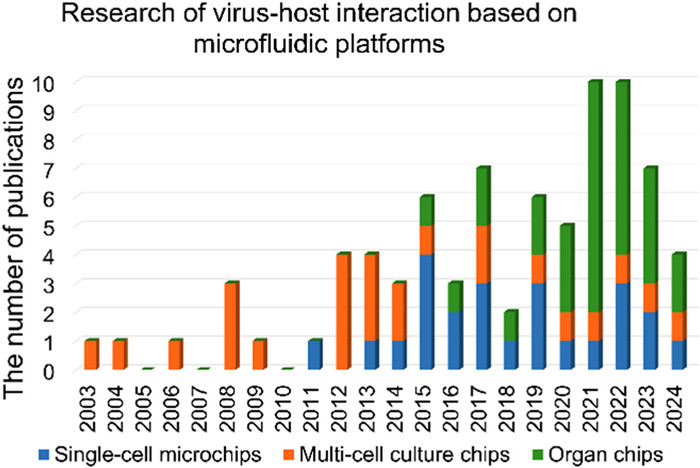
Microfluidics, which emerged in the 1990s, as an interdisciplinary technology platform, has played irreplaceable role in biological and medical fields, including point-of-care testing (POCT), physiological and pathological processes exploration and drug development [9-11]. Since Yin’s group first characterized vesicular stomatitis virus (VSV) propagation on micropatterned cell culture chip in 2003 [12], more and more attempts were made by scholars to explore the interactions between living viruses and host cells utilizing microfluidic techniques (Fig. 1). A statistical analysis of publications from the past two decades reveals a strong correlation between the advancements in cellular microchips and the increasing utilization of microfluidics in viral cell biology. The remarkable progress in single-cell chips and organ-on-a-chip (organ chip) technology has greatly facilitated the integration of microfluidics with single-cell virology and the modeling of viral diseases [8,13-15]. This integration is extensively documented, with reports dating back to 2011 and 2015, respectively, as depicted in Fig. 1. In particular, during COVID-19 pandemic, organ chip has revolutionized the modeling novel virus infection in human tissues [16-19]. This has significantly raised researchers’ awareness of its great potentials in dealing with emerging infectious diseases, as could be glimpsed from the sharp increase in publications after 2019 (Fig. 1). This review aims at fully presenting an overview of microfluidics applications in virus-host interactions. The opportunities and challenges of microfluidics in developing robust virological methods in vitro are also discussed. We hope that this review will provide useful guidelines to pursuers interested in this area, encouraging new fruitful exchanges and collaborations of specialists across multiple disciplines.
Figure 1
Figure 1.
Statistical analysis of virus-host interaction publications based on microfluidic platforms (as of April 2024). Note: This statistical analysis excludes studies that only use commercial single-cell RNA sequencing instruments without involving the fabrication or improvement of single-cell microchip.
2.
Conventional methods for studying virus-host interactions
Viruses, as restrictive parasitic microbe rely on the operating machinery within host cells to implement infection and dissemination. Generally, entire viral life cycle is composed of viral particles entry, replication, assembly and release [20]. Released virions perform cell-free virus spread through particle diffusion, whereas intracellular virions via directly delivery between contacting cells execute more efficient cell-to-cell spread [21]. Deciphering these complicated processes are basic issues in virus-host interactions. Plaque assay is classical method for quantifying and characterizing viral strains capable of forming plaques on host cell monolayers due to cytopathic effect (CPE) [22-24]. Infectious virion particle concentration of a virus sample, known as viral titer, can be calculated in plaque-forming units (pfu) or TCID50 by this method [23]. Virion cell-to-cell transmission can also be characterized through plaque shape and dimensional changes over the infection time [24]. The titer change of intracellular and extracellular viruses over infection time is known as one-step growth curve, reflecting viral infection dynamics [25,26]. Additionally, routine immunoassays such as ELISA, immunostaining and western blot are applicable for parsing viral and cellular protein interplays [27]. These classic virological approaches help researchers to explore viral events, based on virus infection models.
Despite the successful controlled human infection (CHI) trials with some viruses [28], humans are not ideal objects to model viral infections due to ethical issues. Currently, the mostly used models in viral cell biology are in vitro transformed cell line models and in vivo animal models (e.g., rodents and primates). This is because their prominent advantages, as described in Figs. 2a and b. Here, stable cell lines are cultivated in Petri dishes or well plates [29], providing convenient condition for open-ended virus incubation. In addition, the ease of handling, affordability, and the ability to directly and easily perform microscopic examinations have made cell line infection models a popular choice in virological research (Fig. 2a). Animal infection models, as a counterpart, are superior in analyzing interorgan communications, individual symptoms and organism immune responses (Fig. 2b). Rodents are widely used due to the availability of abundant standard strains and relative low-cost. Other small and large animals, such as ferrets, cats, dogs, cattle, sheep, and pigs, also provide a pool of valuable candidates [30,31]. Processing the closest relatives to humans, primates are widely considered the gold standard model for preclinical studies on vaccines and antivirals [31].
Figure 2
Figure 2.
Conventional methods and microfluidic technology in viral cell biology research. (a) Traditional cell infection models were constructed by culturing transformed cell lines, primary cells, or stem cells in Petri dishes or well plates, and their advantages and disadvantages were listed. (b) Animal infection models were established on a variety of animals ranging from small to large, and their superiority and inherent shortcomings were described. (c) Taking the technical development of cell microfluidic chips as a clue, the advantages and main applications of single-cell microchip, multi-cell culture chip and organ chip in addressing virus-host interactions were elaborated.
Conventional methods and models have greatly contributed to the understanding of virus-host interactions over the past decades. However, the inherent shortcomings of traditional cell culture techniques and animal models, as shown in Figs. 2a and b, also bring some obstacles to fully cognize virus-host interactions. For example, the planar and static culture conditions are far from real extracellular microenvironment under physiological conditions (Fig. 2a), and the resulted differences in cell morphology and function may affect the outcomes of viral infection [7,29]. Certainly, cells derived from primary tissue or human pluripotent stem cells (PSCs) can be alternatives for better maintaining phenotypic and genotypic features of physiological condition (Fig. 2a), but high-cost, technical barriers in obtaining and maintaining these cells, and difficulty in mimicking in vivo extracellular environment in conventional cell culture settings hinder their widespread use [29]. While for animal models, they may encounter challenges in terms of ethics, economics, and species gap (Fig. 2b). Species-determined differences in anatomy, pathophysiology, immune response would increase the failure risk in vaccines and antivirals development from laboratory experiments to clinical trials [29-31]. Moreover, complex procedures and longer experimental periods are unfavorable for the urgent investigation needs of emerging epidemics.
Cell-to-cell heterogeneity naturally exist in the genome, transcriptome, proteome, and metabolome [8,13]. Additionally, individual infectious units delivered to each cell could be variable. For example, RNA viruses, with their high mutation rates, often exist as viral quasispecies consisting of both intact and defective virions [8,32]. The demonstrated competition roles of defective viruses significantly affect viral production [32]. The multiple layers of heterogeneity from host cells to viral populations predicts unequal outcomes of viral infection and dissemination, while these variations are often overlooked and masked in bulk assays [8,13,33]. Improving current viral infection models to achieve accurate simulations of extracellular microenvironment, humanized tissues or organs, and robust single-cell infection mode presents unprecedented opportunities for new discoveries in viral cell biology.
3.
Advantages of microfluidic technology in viral cell biology
Microfluidics, also called as lab-on-a-chip (LOC) or micro total analysis system (µTAS) is characterized by having micro/nanochannel in at least one dimension where fluid exhibits special properties including laminar flow, high diffusion efficiency and rapid transfer of heat and mass [9]. With the traits of low sample consumption, miniaturization, automation, portability and high throughput, microfluidics exhibited remarkable benefits for chemical and biological analyses on the microscale [10,13]. Particularly, microfluidic platforms have commensurate characteristic sizes with biological entities (e.g., cells, bacterial and viruses) and enable flexible control of fluids, which are conducive to precise manipulation of cells and accurate simulation of extracellular microenvironment, providing powerful technique supports for improving viral infection models. Microfluidic technology owns unique advantages in addressing major concerns in viral cell biology, as shown in Fig. 2c. Firstly, microfluidic platforms offer flexible and customizable cell operation capabilities that surpass traditional in vitro cell culture methods. Up to now, various single-cell microchips have been developed to analyze tumor cell heterogeneity, bacterial drug resistance, stem cell differentiation, and immune response differences [34-37]. Cell patterning and multi-cells co-culture platforms can also be easily implemented through surface modification or microstructure control enabling the visualization of diverse biological processes [38,39]. Benefiting from microfluidic-based controllable cell culture, high-throughput single-cell infection mode and regionalized multi-cells infection mode can be achieved, facilitating to develop efficient single-cell virology and humanized tissues or organs infection models.
Next, microfluidics holds tremendous potentials for simulating extracellular microenvironment under physiological conditions (Fig. 2c). A series of biochemical and biophysical external cues can be reproduced on diverse microchips, including stable concentration gradients of cytokines, extracellular matrix (ECM) with different topographies, stiffness, and proteins, and various mechanical stresses (shear, stretch, tension) [40-42]. One notable advantage is the ability to achieve biomimetic fluid flows in vessels, lymphatics, and renal tubules, which further enhances the realism of the in vivo microenvironment [14]. Base on this, diverse organ chips designed to reconstruct physiological functions have developed, including heart, liver, spleen, lung, kidney, gut, blood-brain-barriers (BBB), skin, placenta and microvascular chips [15,19,29,43,44]. As an expected alternative to animal models, they have been widely used in disease modelling, drug development, personalized medicine and regenerative pharmacology [11,45].
In addition, microfluidic platforms provide access to integrated application with various testing facilities, among which optical microscope are mostly used, enabling real-time monitoring and efficient recording of viral events. On the contrary, these characterizations are often labor-intensive and technical difficult to achieve via routine assays. Finally, microfluidic technology provides a cost-saving, environmental- and animal-friendly option for modeling viral infection [7,29,31]. The miniaturized nature of the devices is ideal for studying rare cells and viruses, while low reagent consumption helps reduce costs and minimize environmental pollution. Applying organ chips to viral infection models can not only reduces the use of animals and the related ethical concern, but also offer a solution for studying viruses that lack suitable animal hosts.
Beyond viral cell biology, microfluidics has also proven to be a useful tool for virus detection and characterization. A large number of microfluidic platforms based on viral nucleic acid and antibodies immunoassays have been released, achieving rapid and sensitive virus detection and quantification, and enriching virus diagnostic methods [10,46,47]. Moreover, microfluidics has proven instrumental in addressing complex tasks for understanding viral variation and evolution, such as single-virus isolation and sequencing [13]. And this is realized by leveraging the potential of microfluidics in terms of high throughput, parallelization, and sensitivity, indicating that microfluidic technology has revolutionized conventional approaches and provided a versatile toolbox for virology research. In the following sections, we will exclude the application of microfluidics in virus detection and characterization, and focus on reviewing how microfluidics harnesses the development of cellular microchips as a chance to tackle key challenges in viral cell biology.
4.
Microfluidics applications in virus-host interactions
4.1
Single-cell chips for uncovering heterogeneity of virus-host interactions
Natural cell-to-cell differences and pre-existing diversity of viral populations are destined for heterogeneous outcomes, including but not limited to viral replication initiation time, viral mRNA production, and viral particle genotypic types. Unraveling these heterogeneities is vital to explain the mechanisms under antiviral resistance and zoonic diseases [8,13,33]. To obtain single-cell resolution, traditional methods are usually conducted on microwell plates via serial dilution [48], capillary micromanipulation [49] and fluorescence activated cell sorting (FACS) [50]. Manual dilution and micromanipulation have extremely laborious workflows, while FACS requires large amounts of pre-labeled cell samples. Furthermore, above approaches are limited in throughput and parallelization and not applicable for large-scale single-cells infection analysis. As the promising substitution, microfluidic technology allows for high-throughput and parallel capture and patterning of single cells using various capture strategies. As schematically shown in Fig. 2c, these strategies include microstructure traps, microwells, microvalves and droplets. Classified by capture strategy and heterogeneity analysis, a comprehensive summary of research on virus-host interactions using single-cell microchips was presented in Table S1 (Supporting information), while representative applications of different types of single-cell chips were shown in Fig. 3.
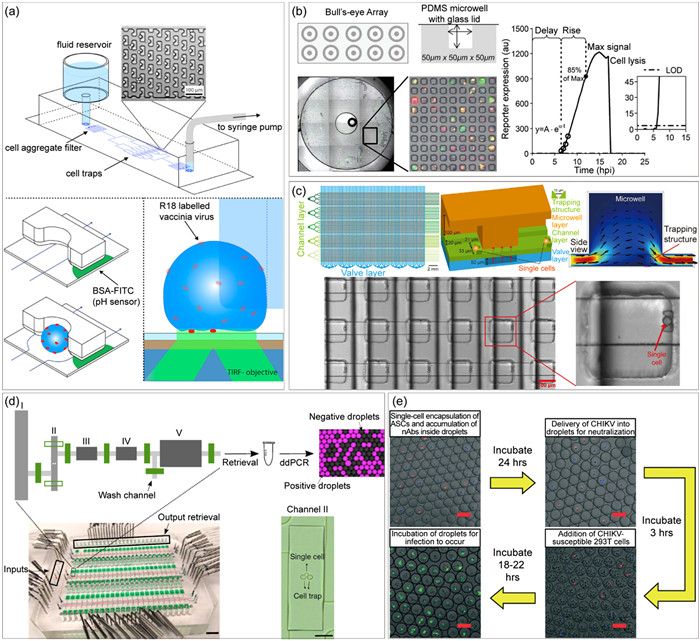
Figure 3
Figure 3.
Single-cell microfluidic platforms for virus-host interactions analysis. (a) Microstructure trap-based single-cell capture array for investigating acid-dependent membrane fusion of vaccinia virions. Reproduced with permission [51]. Copyright 2013, Biophysical Society. (b) A microwell array device for quantitative profiling of innate immune activation by viral infection in single cells. Reproduced with permission [56]. Copyright 2017, Royal Society of Chemistry. (c) Single-cell analysis of the activity of antiviral agents on a multilayer device equipped with microvalves and microstructure traps. Reproduced with permission [57]. Copyright 2019, The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science, under the CC BY-NC license. (d) An automated valve-based microfluidic system with cell-trapping microstructures for multi-parameter and ultra-sensitive protein/mRNA measurements. Reproduced with permission [81]. Copyright 2019, The Authors, under the CC BY license. (e) High-thought screening and enrichment of cells secreting virus neutralizing antibodies on a droplet-based microfluidic system. Reproduced with permission [87]. Copyright 2022, Royal Society of Chemistry, under the CC BY-NC license.
Monitoring viral infection at the single-cell level requires non-invasive and continuous acquisition of information, akin to the classical virus one-step curve that measures changes in viral particles and nucleic acid production [25,26]. Microfluidic platforms have emerged as ideal tools for coupling in vivo real-time imaging and testing technology to meet these demands. Several single-cell microchips embedded with microstructure traps array were developed to investigate the acid-dependent kinetic fusion of single vaccinia virus (VACV) on the plasma membrane (Fig. 3a) [51], evaluate the heterogeneous HIV activation in single Jurkat T cells stimulated by latency reversing agents (LRAs) [52], and explore infection heterogeneity of H1N1 in the presence of gradient bafilomycin A1 [53]. These microstructure traps-based devices have robust ability for single-cell capture but fail to offer separate compartments to avoid cross-talk among infected cells. To address this limitation, Eid et al. modified microstructure trap-based device by incorporating independent microchannels behind each trap site, and successfully monitored the kinetics of HIV-like particles (VLPs) releasing in single-cell level [54].
Meanwhile, strategies based on microwells, microvalves and droplets are preferred to provide compartmentalized space for unambiguous analysis of single-cell viral infection [26,32,55-59]. Given attractive simplicity of microwells, Yin’s group established a bull’s-eye polydimethylsiloxane (PDMS) array-based device covered with a glass slide lid, in which each bull’s-eye unit contained 2500 cubic microwells [55,56]. By the integration with a dual-color fluorescent reporter and imaging system, this platform enables the characterization of viral and cellular dynamic gene expression, and the visualization of innate immune activation in single-cell level during VSV infection (Fig. 3b) [56]. This platform also facilitated the investigation about the impact of defective interfering particles (DIPs) on VSV protein production, by the co-infection of BHK cells with different proportions of DIPs and other infectious particles [32]. Besides, microvalves-based array chips offer better automation and integration capabilities, supporting multi-step processing and multi-parameter simultaneous analysis. For instance, Guo et al. constructed a microvalve-based PDMS device featured 4 parallel unit, and had the capacity to accommodate up to 6400 individual microchambers [26]. By employing GFP-poliovirus (PV) as RNA virus model, their study revealed that the start time of viral replication was mostly determined by cellular factors, while the replication speed and titer yields depended on virus itself.
However, the single-cell occupancy in microwells- and microvalves-based devices generally does not exceed 30% due to reliance on the Poisson distribution. To maximize the compartments efficiency, Liu et al. combined the strategies of both microstructure traps and microvalves, and developed a multilayer PDMS device with 5700 microchambers, and each equipped a dam-type trap (Fig. 3c) [57]. Using this system, researchers evaluated the efficacy of a wide range of enterovirus inhibitors and antiviral candidates, which will help in antiviral drug clustering and resistance prediction. Recently, droplet-based microfluidics enabled single-cell-level high resolution viral genomes analysis of influenza A virus (IAV) during infection [58], as well as the kinetics of initial gene expression and replication of herpes simplex virus type 1 (HSV-1) [59]. Notably, while most single-cell studies of virus-host interactions were focused on manipulating single cells, the Arai team reported a novel approach by utilizing optical tweezers in conjunction with dielectrophoresis to manipulate single viruses and infect selected cells [60,61]. Although this approach is still in its early stages, it holds significant potential for exploring the intricacies of single-virus infection.
4.1.2
Viral and cellular transcriptomic
The differential phenotypes of viral infection are essentially caused by the heterogeneity of mRNA transcription and protein expression. Viral and cellular transcriptomic analysis will help us understand viral replication, diversity and evolution, and profile heterogeneous cellular response to viral infection, especially in identifying cell subpopulations [62,63]. Achieving this goal requires single-cell RNA sequencing (scRNA-seq), which involves single-cell capture, cell lysis, mRNA extraction, reverse transcription, DNA amplification, and sequencing [62,63]. Therefore, traditional scRNA-seq methods entail highly complex workflows and necessitate manual manipulation of numerous plates.
To simplify this procedure, a microwell-based single-cell microfluidic platform was reported for semi-automatic full-length scRNA-seq of HPV-infected cells [64]. However, sequential addition of different reagents to each well is still required. In contrast, microvalves- and droplets-based single-cell platforms, with their superior capabilities in automation, integration and high-throughput, provides valuable resource to drive the commercialization and ubiquity for scRNA-seq. The widely used Fluidigm C1 and 10x Genomics are developed upon high-density microvalves chips, called integrated fluidic circuits (IFCs), and open-source droplet sequencing (Drop-Seq) pipeline, respectively [65-67]. Using above commercial equipment, numerous transcripts profiling studies were conducted, successfully revealing the heterogeneity in virus gene expression during the infection of viruses, including IAV [68,69], cytomegalovirus (CMV) [70,71], ZIKA [72], HIV [73] and HPV [74]. Moreover, these studies have helped to identify cells subpopulations by virus entry receptors [75], biomarkers of virus permissiveness [76], and initiating antiviral program [77]. However, current Drop-Seq and 10x Genomics technologies could only quantify A-tailed mRNA transcripts. Given that, some studies have reported improved droplet microchips, enabling multiplexed amplicon sequencing and transcriptome profiling [78], directly measurement of unspliced and multiply spliced RNA [79], and large-scale quantitative analysis of viral genotypes using customized barcoded primers [80]. To correlate single-cell viral infection phenotypes with corresponding transcripts analyses, Lin et al. developed a microvalves-based device embedded with microstructure traps to perform automatically multi-parameter and ultra-sensitive protein/mRNA measurements in single cells (Fig. 3d) [81]. In addition, Cortés-Llanos et al. recently reported a microwell array to monitor the T-lymphocyte responses to LRAs treatment, and perform scRNA-seq on released single cells via magnetic cell manipulation [82]. Collectively, microfluidics serves as an efficient scRNA-seq tool, promoting the elucidation of cellular and molecular mechanisms underlying viral diseases.
4.1.3
High-throughput screening
Droplet microfluidics, with its superior high-throughput and parallelization capabilities, is an ideal tool for inquiring virus variants and evolution, as well as screening effective neutralizing antibodies (nAbs) and antivirals. Fischer et al. utilized a water-in-oil droplets chip to isolate viral variants that escaped from antibody evolutionary stressors [83], while Weitz and co-workers modeled the fitness landscapes of murine norovirus (MNV) with and without nAbs selection pressure by insulating and passaging small viral populations within droplets for 5 times [84]. Besides, viral infectivity assessments, as the core element for antivirals and nAb screenings, usually take days or weeks by traditional plaque assays. To expedite infectivity assessment, Weitz’s group developed a droplet-based, rapid, and culture-free infectivity assay that enabled infection, incubation and quantification of infectious virions in one day [85]. Additionally, given the potential for irrelevant hits when using traditional antigen-antibody binding assays for antibody identification, several high-resolution droplet screening platforms have been developed. These platforms focused on antibody function and efficacy to identify nAbs repertoires produced by hybridoma cells during murine hepatitis virus (MHV) infection [86], and enable effective nAbs retrieval and Ab-secreting cell enrichment against Chikungunya virus (CHIKV) (Fig. 3e) [87]. These droplet-based microchips facilitate the high-throughput screening of nAbs and antiviral agents against an extensive range of viruses and biohazards, as well as the identification and isolation of drug-resistant viral strains.
4.2
Multi-cell culture chips for viral life cycle monitoring
Conventional CPE-based plaque assays are primary methods to understand viral infection and transmission events in vitro, despite that the workflows are usually reagent-, time- and labor-intensive. In plaque assays, viruses act on permissive cells in a free-diffusion manner, resulting in uncertainty about plaque sites, which is unfavorable for long-term viral events monitor. Moreover, the two-dimensional and static cell culture conditions also limit the cognition of viral events under physiological environment. Microfluidic multi-cell culture platforms are able to fulfill the above-mentioned research limitations, and have made remarkable achievements in controllable cell patterns, simulation of extracellular microenvironment particularly biofluids (Fig. 2c), and long-term dynamic monitoring. An overall summary of research on virus-host interactions using multi-cell culture microfluidic chips was provided in Table S2 (Supporting information), organized by applications and cell culture methods. Deeper insights into viral infection, dissemination, and virus-induced cellular events on self-designed cell culture or co-culture platforms were representatively showcased in Fig. 4.
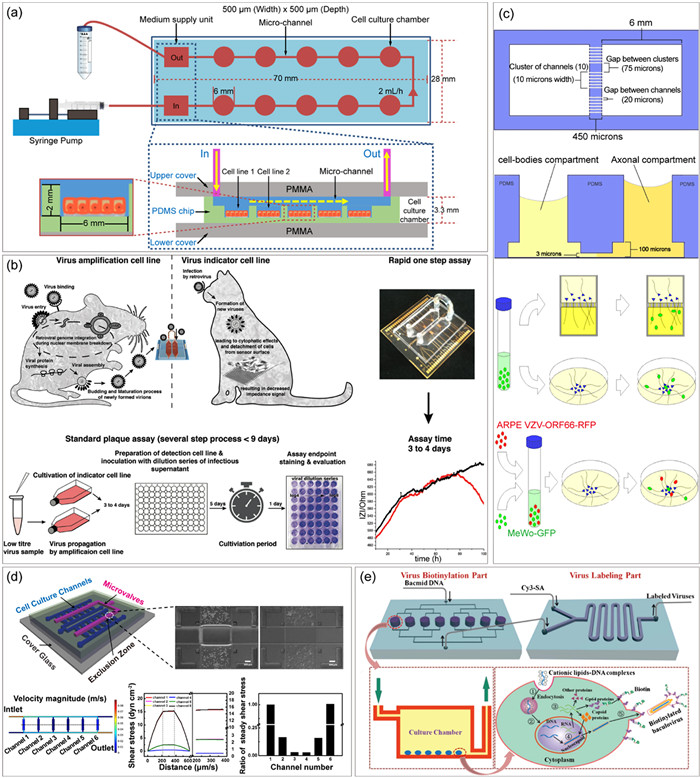
Figure 4
Figure 4.
Multi-cell culture microfluidic platforms for viral events analysis. (a) A customized device supporting synchronously culture of different cell lines for rapid susceptible cells screening. Copied with permission [22]. Copyright 2022, The Authors, published by Elsevier B.V. on behalf of KeAi Communications Co. Ltd., under the CC BY-NC-ND license. (b) An impedance-based microfluidic chip for rapid infectivity assay. Reproduced with permission [23]. Copyright 2021, Royal Society of Chemistry, under the CC BY license. (c) A co-culture microchip designed to monitor viral and cellular proteins transfer between infected non-neuronal cells and human axons. Reproduced with permission [102]. Copyright 2015, Grigoryan et al., under the CC BY License. (d) A multi-shear microfluidic platform for exploring the effect of shear stress on vaccina virus-induced cell migration. Reproduced with permission [41]. Copyright 2017, Royal Society of Chemistry. (e) Site-specific labeling of baculovirus on an integrated microfluidic system. Copied with permission [110]. Copyright 2013, Royal Society of Chemistry.
Initial efforts regarding viral events on microfluidic platforms are to reduce experimental workloads by simple microfluids manipulation and substrate patterning. For instance, viral infections were performed under different multiplicity of infections (MOIs) on a single three-stream microdevice [88]. And the purification of specific viruses from a large virus population was also realized by employing size-controlled cell patterns [89]. Certainly, selection of suitable and susceptible cell lines is the basis for viral infectivity assessment. Given that traditional plate-based methods are often complex and laborious, Su et al. developed a microfluidic chip with 10 chambers to culture multiple cell lines in parallel, and achieved rapid screening of susceptible host cells for enterovirus 71 (EV71) and H1N1 (Fig. 4a) [22]. This work also proposed a rapid identification method for emerging viruses.
Benefiting from the excellent performance of the microfluidic platforms in live imaging, real-time, in situ and long-term monitoring of infection processes of various viruses, including retrovirus [90], hepatitis B virus (HBV) [91], pseudorabies virus (PRV) [25], and adenovirus [92] were successfully achieved. Among these works, Elvassore’s group found that shifting virus delivery mode, from diffusive-limited to convective-dominated mode, could enhance infection efficiency of adenoviral, even at low MOIs [92]. Similarly, Purtscher et al. observed an elevate infection level of PG-4 reporter cells under continuous fluid flow, using an electrical sensor array-based microfluidic platform (Fig. 4b) [23]. Meanwhile, this platform significantly shortened experimental period and served as an efficient alternative to standard plaque assays involving cell-based viral amplification, virion release and CPE detection. Since viruses commonly serve as in vitro gene transfer vectors, findings above have inspired scholars to explore optimized transfection conditions utilizing microchips. Elvassore’s group, for instance, realized efficient viral transduction at low viral doses, by repeated pulsed cell-virus contact [93], while Ahmadi et al. demonstrated a nearly 9-folds increase in HIV-1 lentiviral vector transduction on microchip compared to that in 96-well plates [94].
Moreover, multicellular microchips supporting online monitoring and rapid readouts, facilitated the assessment of factors affecting infection. For example, large-scale screening of interfering RNAs for VSV and Rift Valley fever virus (RVFV) was performed on a PDMS-based dot-matrix device [95]. And the evaluation of carrageenan antiviral drugs against dengue virus (DENV) was achieved by real-time on-chip cellular oxygen consumption rate monitoring [96]. Beyond adherent cells, microfluidics also enables long-term viral infection tracking and imaging in suspension cells. Using a microwell-based device with restricting finger-like channels, Razooky et al. characterized single cell gene-expression kinetics during infection by four different HIV-1 variants [97].
4.2.2
Viral spread
Cell-to-cell spread is recognized as a highly efficient dissemination route for enveloped viruses. In traditional plaque assays, semi-solid medium is applied over a confluent cell monolayer to get rid of virion free-diffusions, therefore intercellular transmission rates can be determined according to the size change of the plaques. Compared to this single and inflexible characterization, microfluidics-based cell patterning allows for the observation of viral dissemination from various perspectives. For example, linearized VSV propagation was observed with linearized cell microarrays [12], and direction-limited PRV spread was captured along the single directional cellular networks based on PEG pattern [98]. Moreover, the impact of external factors on virus transmission behavior has been investigated by simulating extracellular microenvironment on microfluidic chips. Yin’s group discovered that fluid flow could enhance viral spread in microchannels [99], and Zhang’s group demonstrated an anisotropic cell-to-cell spread of VACV on a microgroove topographic PDMS substrate [24].
Thanks to cell co-culture microchips, virion transmission between different kinds of cells could be effectively visualized in more detail. Liu et al. designed a co-culture system featuring two chambers connected with microchannels to study neuron-to-cell spread and neuronal axonal transport of PRV [100]. Results demonstrated that immature nucleocapsids lacking tegument and envelope go through retrograde transport, as opposed to the anterograde transport of mature PRV particles during viral egress in axons. A similar retrograde axonal transport of labeled alpha-herpesvirus varicella zoster virus (VZV) capsid was visualized by Goldstein’s group using compartmentalized microfluidic chambers [101]. Based on the same platform, Goldstein’s group also demonstrated the direct transfer of viral and cellular proteins from VZV-infected non-neural cells to human embryonic stem cells (hESC)-derived neuronal axons, suggesting the fusion of neurons into syncytia of VZV-infected cells (Fig. 4c) [102]. Recently, based on an epithelial cell-neurons co-culture chip, Sun et al. found that HSV-1 glycoproteins could alter the protein composition of extracellular vesicles, promote neurite outgrowth and facilitate infection of peripheral neurons [103]. Briefly, cell co-culture microchips, when coupled with real-time imaging, enable the capture of immunocytochemical evidence to clarify the mode and mechanisms of intercellular viral transmission.
4.2.3
Virus-induced cellular events
Virus invasion can cause not only basic infection and dissemination but also morphological and behavioral changes in cells, such as the formation of virus-induced specialized subcellular structures and cell migration. Multi-cell culture chips offer valuable tools for uncovering potential correlations involved in virus-induced cellular events. Tunneling nanotubes (TNTs) are considered as vital physical connection between cells for intercellular communication [104]. On a microchip with high-throughput side-chambers, Zhang’s group monitored distinct patterns of VACV-induced TNTs formation, showing the effect of virus titer on TNTs formation [105]. It is well-known that some viruses, such as West Nile virus (WNV), human cytomegalovirus (HCMV), Epstein-Barr virus (EBV) and VACV, can trigger cell migration after infecting cells derived from specific tissues [106-109]. This phenomenon is thought to be closely related to immune activation, cardiovascular disease, tumor metastasis, viral diffusion, etc. [106-109]. Zhang’s group developed various microfluidic platforms which integrated steady multi-shear stress (Fig. 4d) [41] and microgroove topographic substrate [42] for VACV-induced cell migration assays. The results showed an accelerated directional cell migration and differential localization of subcellular organelles in response to different external cues. Notably, Zhang’s group also reported a microfluidic-based labeling method for enveloped viruses (Fig. 4e) [110]. Biotinylated viruses were obtained during the viral life cycle and conjugated with fluorescently labeled avidin to achieve baculovirus labeling. Collectively, multi-cell microfluidic chips are capable of adapting virus incubation methods, modifying traditional plaque assays, and optimizing long-term viral events monitoring, which open a window for deeply understanding the interactions between the virus, host cells and microenvironment.
4.3
Organ chips for modeling viral infectious diseases
Viruses invading and transmission among hosts may result in viral infectious diseases, as we have experienced the influenza and COVID-19 pandemics. Organ chips, also known as microfluidic microphysiological systems, aims to mimic organ-level physiological features and functions, by restoring the tissue-tissue interfaces, extracellular biomechanical and biochemical cues in vitro [7,29]. These bionic microfluidic chips as schematically shown in Fig. 2c, serve as a cost-effective alternative to animal models, facilitating the exploration of viral infectious diseases, especially in understanding the organism responses to virus invasion and the development of antiviral strategies. Currently, lung-, liver-, gut-, kidney-, BBB-, microvascular system- and lymphoid follicle (LF)-on-a-chip models have been built and employed to investigate viral pathogenesis and evaluate antiviral efficacies for respiratory viruses, hepatitis viruses, and so on. Representative viral diseases models constructed using organ chips mentioned above were exhibited in Figs. 5 and 6, while a comprehensive overview of the use of organ chips in studying virus-host interactions was presented in Table S3 (Supporting information), categorized by organ types and target viruses. Notably, the SARS-CoV-2 and IAV have received the most research attentions [27,111-127].
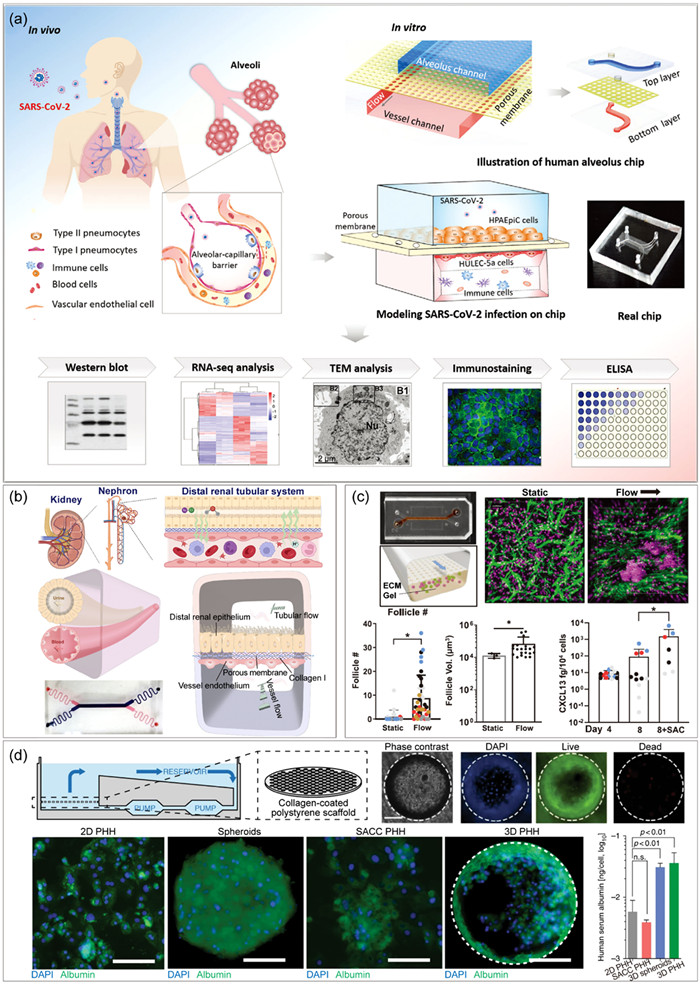
Figure 5
Figure 5.
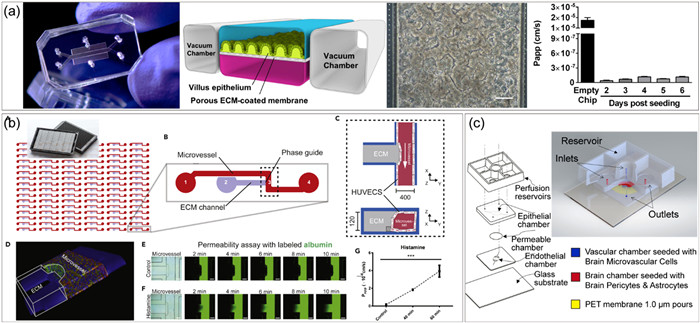
Organ-on-a-chip for viral infectious diseases modeling. (a) Microengineered human alveolus chip for modeling SARS-CoV-2-induced lung injury. Copied with permission [27]. Copyright 2020, The Authors, published by Wiley-VCH GmbH, under the CC BY license. (b) Distal renal tubular system-on-a-chip for exploring the pathogenesis of influenza A virus-induced kidney injury. Reproduced with permission [125]. Copyright 2023, Royal Society of Chemistry. (c) A human lymphoid follicles chip for exploring seasonal influenza vaccination responses. Reproduced with permission [126]. Copyright 2022, The Authors, published by Wiley-VCH GmbH, under the CC BY license. (d) 3D microfluidic liver cultures for hepatitis B virus infection research. Reproduced with permission [135]. Copyright 2018, The Authors, under the CC BY license.
Figure 6.
Organ-on-a-chip for viral infectious diseases modeling. (a) Human gut-on-a-chip for modeling enterovirus infection. Reproduced with permission [138]. Copyright 2017, under the CC0 license. (b) A human microvessel-on-a-chip for reproducing Ebola hemorrhagic shock syndrome. Copied with permission [139]. Copyright 2019, The Authors, under the CC BY-NC-ND license. (c) A human blood brain barrier chip for evaluation antivirals against encephalitis virus infection. Reproduced with permission [140]. Copyright 2022, The Authors, under the CC BY license.
Respiratory viruses invade the human body, and the lungs are considered to be the initial target organs. Currently developed in vitro lung-on-a-chip models include airway-on-a-chip and alveolar-on-a-chip, both characterized by the formation of air-liquid interface with the regionalized co-culture of epithelial lung cells and endothelial microvascular lung cells [7,18]. Briefly, a porous membrane is used to mimic airway or alveolar septum. The lower microchannel is seeded with vascular endothelial cells and applied with continuously perfusion, simulating capillary and fluid shear stress. The difference lies in the upper microchannel where alveolar epithelial cells or human primary airway epithelial cells are exposed to air or oxygen. Together with periodic stretching of porous membrane controlled by vacuum pumps connected to the 2 chambers on the both sides of main channel, the typical extracellular physical environment of alveolar or airway are reproduced.
Based on this technique breakthrough, numerous studies have been conducted on lung-on-chip models, trying to explore the respiratory diseases caused by viral infections [27,111-120,128,129]. Nawroth et al. described a human rhinovirus 16 (HRV16)-induced asthma exacerbation model on an airway chip, and found that IL-13 may impair host immune response to HRV16 infection [129]. Ingber’s group developed a human lung airway chip model, which helped to rebuild the evolution of IAV through mutation and gene reassortment during host-to-host transmission [111]. They also developed a human alveolus chip, and unveiled the impact of mechanical breathing on innate immune responses to IAV infection [112]. A bacterial-IAV co-infection immune-responsive alveolus chip model was reported by Deinhardt-Emmer et al., showing a significant endothelial cell damage associated with barrier function loss upon infection, implying worsening respiratory illness [113]. Furthermore, in response to the SARS-CoV-2 pandemic, many researchers developed SARS-CoV-2-induced lung injury microchip models to elucidate viral pathogenesis, as well as effectiveness evaluation of antibodies and antiviral drugs [27,114-117]. Qin’s group revealed crucial roles of immune cells in alveolar barrier injury and exacerbated inflammation during SARS-CoV-2 infection (Fig. 5a) [27], while Thacker et al. demonstrated the occurrence of SARS-CoV-2-mediated endothelial cell damage independently from cytokine storm [114]. Recently, a gravity-perfused airway-chip especially designed for biosafety level 3 (BSL-3) studies was developed, and successfully recapitulated tissue response and immune response to SARS-CoV-2 infection [115]. Additionally, the differences of IAV and coronavirus in pathogenicity and therapeutics were synchronously compared on lung chips [118-120]. In this research aspect, Ingber’s group realized the rapid identification of drug candidates against IAV and pseudotyped SARS-CoV-2 infection on airway chips [119], while Yadav et al. found distinct immune responses to SARS-CoV-2 and IAV on 2 different types of lung chips [120].
Beyond lungs, clinical cases indicate that infection and injuries to multiple secondary organs greatly increase morbidity and mortality from respiratory viruses. Utilizing various organ chips, researchers demonstrated that SARS-CoV-2 infection disrupted intestinal and vascular barrier function [121,122], caused BBB dysfunction and neuroinflammation [123], and led to hepatic dysfunctions [124]. Similar compromised barrier function was also observed in vascular endothelium and intestinal epithelium during infection by human coronavirus NL63 (HCoV-NL63) [122,130], a model virus that shares the same angiotensin-converting enzyme 2 (ACE2) receptor with SARS-CoV-2. Notably, Zhang’s group recently developed a distal renal tubular system-on-a-chip to reveal the pathogenesis of IAV-induced acute kidney injury (Fig. 5b) [125]. This work demonstrated that virus entered the kidney by cell-free transmission without disrupting the vascular barrier. Additionally, a series of subcellular responses to viral infection, including disrupting intercellular tight junction, epithelial microvilli, transporter polar distribution and inorganic ion reabsorption, were consistent with their previous work about PRV infection in kidney tubes [131]. Besides, lymphoid chips were developed, showing great potential in human seasonal influenza vaccine (Fig. 5c) and SARS-CoV-2 vaccine test [126,127]. In summary, organ chips offer valuable and resource-rich platforms for human viral respiratory and extra-respiratory disease investigation, pathogenesis exploration, and antiviral drug identification.
4.3.2
Hepatitis viruses
A chronic infection of hepatitis virus is closely linked to the development of severe liver diseases. Hepatitis B virus (HBV) and hepatitis C virus (HCV) are two major health concerns globally, infecting 260 and 180 million people respectively [132]. As a complex physiological system, liver exerts multiple functions and consist of multiple inter-connected parenchymal, non-parenchymal, and immune cell sub-types. However, conventional liver infection model only includes human primary hepatocytes (PHHs), which would rapidly de-differentiate under traditional culture after isolating from liver. Furthermore, due to the narrow host range of hepatitis viruses, available animal models are extremely limited. The developed liver chip platforms enable the study of viral hepatitis diseases in a more physiological relevant environment. Noh’s group firstly reported a complete in vitro HBV replication cycle on a rat liver sinusoid chip [133]. After that, they applied this model to PHH culture, obtained stable morphology and viability for up to 26 days, and successfully detected cell-secreted HBV DNA and HBV core antigen [134]. Another PHHs-derived liver model with recirculation of nutrients and oxygen was reported by Dorner’s group, which could last for at least 40 days and reproduce full HBV life cycle (Fig. 5d) [135]. Furthermore, co-culture with non-parenchymal Kupffer cells enabled identification of cellular origin of immune effectors, while the simulated sinusoid microarchitecture system permitted to explore the roles of different liver-resident cell subtypes in anti-HBV response [136]. Moreover, Natarajan et al. recently developed an adult stem cells-induced liver model, with the co-culture of CD8+ T cells, and achieved study of human adaptive immune responses to HCV at a molecular level [137]. All these liver chips above reconstitute a more complex liver microenvironment and architecture in vitro, which is crucial for understanding viral hepatitis pathogenesis, immune evasion mechanisms and therapeutic drugs effectiveness.
4.3.3
Other viruses
Except for the extensively studied respiratory and hepatitis viruses, diversified organ chips make it possible to gain insight into the infection events of various human viruses, including enterovirus [138], herpesvirus [131], ebolavirus [139] and alphavirus [140]. The human gastrointestinal tract is structurally dynamic, complex, and susceptible to bacterial, parasitic, and viral infections. However, animal models suitable for enterovirus infection research are limited due to the difference in virus receptor expression between experimental animals and humans. To solve this problem, Ingber’s group developed a gut chip mimicking the physiological flow and peristalsis-like motions of human intestine, and introduced coxsackievirus B serotype 1 (CVB1) as well to build the viral infection gut model (Fig. 6a) [138]. A polarized release mode of cytokines and infectious virus production were reproduced. Beyond gut, kidney contains specialized structures and physiological flows, which are difficult to mimic in conventional static culture system. Zhang’s group reported a kidney chip that reproduced the interstitial fluid environment in distal renal tubules, by integrating a microchannel, a porous membrane and a static liquid well, and explored the pathogenesis of PRV-induced kidney dysfunctions for the first time [131]. Besides, Junaid et al. successfully reestablished vascular barrier function under continuous perfusion using a phase-guided microvascular chip system (Fig. 6b) [139]. This platform has demonstrated the loss of vascular integrity induced by EBOV, and provided a promising model for viral hemorrhagic syndromes research. Recently, Boghdeh et al. partially recapitulated human BBB function on a gravity-flow neurovascular unit and evaluated proinflammatory responses and drug treatment effects following Venezuelan equine encephalitis virus (VEEV) infection (Fig. 6c) [140].
5.
Challenges and future prospective
Compared to the rapid popularity of microfluidics in virus POCT, the utilization of microchips in virus-host interactions research started relatively late and remains relatively rare. Diverse cellular microfluidic chips offer options for improving conventional virological approaches and obtaining innovational discoveries in viral events, while they also encounter obstacles in extensive applications. Despite the flexible and integration-friendly nature of microfluidic platform to meet diverse analytical needs, it also requires various peripheral settings, including optics, pneumatics, electronics, thermals, magnetics, acoustics and other equipment to drive the entire system and perform characterization [13]. Typically, an online cell culture system equipped with a CO2 cylinder, confocal fluorescence microscopy, pneumatic valve control system, and perfusion pumps would commonly be used in microchips-based virus-host interactions research, which creates a dilemma of portable microchip being surrounded by a large array of equipment [41,42]. Besides, the manufacture of chips also requires specialized environments and equipment, such as cleanrooms and photolithography machines. Therefore, their production and usage tend to be relatively high-cost. Furthermore, research on certain highly pathogenic viruses must be performed in laboratories classified as BSL-3 or higher, yet most of the above-mentioned peripherals cannot readily meet the requirements [115]. Moreover, special personnels from multiple professional backgrounds are needed for experiment performing. Additionally, there is a lack of universal gold standards for microfluidic-based virus-host studies. These limitations make it difficult for many labs to acquire the necessary infrastructure and resources, thereby hindering the wide-scale adoption and translation of scientific research findings into practical clinical applications. As a result, although organ chips are designed to closely emulate physiological conditions and proved to hold great promise in drug screening, they have not yet reached the stage where they can fully replace living animals in preclinical testing.
Facing these challenges, developing standardized, automated, commercialized, and affordable “virus-cell” microfluidic systems may be a viable path to promote the widespread adoption of microfluidics in the field of viral cell biology. Automated cell handling systems are highly preferred for single cell manipulation, patterned cell seeding, cell culture, co-culture, as well as extracellular environment simulation for organ chip platforms. Automated virus handling systems are also crucial for flexible adjustment of virus infection MOIs and time points, as well as the manipulation of single virions. Furthermore, specially designed minimized integrable on-line characterization instruments are also demanded for on-line in-situ virus-host interaction observation and data collection. Ideal modules include imaging units, DNA and RNA sequencing units, and the spectrometry units for proteomics and metabolomics study. In addition, integrable organ and tissue relevant function evaluation modules are also necessary for the virus-host interaction study based on organ chips. These modules enable real-time tracking of organ function changes during virus infections. Last but not the least, biosafety issues should always be considered in module developing for virus-related research. The entire integrated system should be sealed, and cultured in specially modified incubators compatible to the whole system, to get rid of contamination and ensure safety.
A deeper integration of microfluidics with other cutting-edge technologies will further propel virus-host interaction research into a new era. Among them, artificial intelligence (AI), fueled by the exponential growth of the computational field in recent years, will undoubtedly play the decisive role. The large amount of raw data obtained from high-throughput microfluidic systems can be processed and integrated using AI. Conventional confocal images, for example, could be transformed to super-resolution images with deep learning training models, providing virus-host interactions information in more details [141]. AI-based bioinformatics integrate data sets obtained from highly automated and integrated microfluidic systems, including imaging data, function-relevant data, genomic data, transcriptomic data, proteomic data, and metabolomic data [142]. This integration helps to yield deeper insights into virus pathogenicity, and provide suggestions for infection prediction that may not be immediately apparent to human analysts. Additionally, with the real-time data processing using big data models, AI can provide real-time characterization recommendations for the next steps and automate adjustments to ongoing experimental parameters. Furthermore, predictive modeling can be provided, guiding high-throughput drug screening and precision medicine.
6.
Conclusions
In this review, we comprehensively summarized the research of microfluidic technology in virus-host interactions, and highlight its significant contributions to the field of viral cell biology over the past two decades. Microfluidics, with its ability in the flexible manipulation of biological entities and the accurate simulation of the extracellular microenvironment, has created diverse innovative platforms for addressing cell biology concerns. These platforms, which include single-cell chips, multi-cell culture chips and organ chips, provide unprecedented opportunities to explore the heterogeneity of both hosts and viruses at single-cell resolution, efficiently monitor viral infection processes within complex physiological microenvironment, and remodel human viral infectious diseases in vitro. Findings obtained on microfluidic platforms, which are often inaccessible using traditionally virological methods, have yielded valuable insight into the pathogenesis of viral diseases and the development of reliable antiviral treatment strategies. As is well known, a brilliant application of microfluidic technology lies in scRNA-seq. If we aspire to replicate such successes in the field of viral cell biology, seeking the core strengths, breaking through the limitations, and establishing a positive transformation from academic exploration to commercialization should be crucial steps. Positioned at the core of multidisciplinary interactions, we have compelling reasons to believe that collaborative efforts from chemists, virologists, computational biologists, and other experts, will pave the way for microfluidics to establish a new paradigm for viral cell biology.
This work was supported by the National Natural Science Foundation of China (Nos. 22274118, 22074107, and 81870359), the Natural Science Foundation of the Jiangsu Higher Education Institutions of China (No. 22KJB320020), and the Health@InnoHK funding (C2i) from Innovation & Technology Commission (ITC) of Hong Kong SAR Government.
Supplementary materials
Supplementary material associated with this article can be found, in the online version, at doi:10.1016/j.cclet.2024.110302.
[1]
V. Hill, G. Githinji, C.B.F. Vogels, et al., Cell Host. Microbe 31 (2023) 861–873. doi: 10.1016/j.chom.2023.03.003
M.R. Karim, T. Islam, M. Shajalal, et al., Brief. Bioinform. 24 (2023) 1–22.
Figure 1
Statistical analysis of virus-host interaction publications based on microfluidic platforms (as of April 2024). Note: This statistical analysis excludes studies that only use commercial single-cell RNA sequencing instruments without involving the fabrication or improvement of single-cell microchip.
Figure 2
Conventional methods and microfluidic technology in viral cell biology research. (a) Traditional cell infection models were constructed by culturing transformed cell lines, primary cells, or stem cells in Petri dishes or well plates, and their advantages and disadvantages were listed. (b) Animal infection models were established on a variety of animals ranging from small to large, and their superiority and inherent shortcomings were described. (c) Taking the technical development of cell microfluidic chips as a clue, the advantages and main applications of single-cell microchip, multi-cell culture chip and organ chip in addressing virus-host interactions were elaborated.
Figure 3
Single-cell microfluidic platforms for virus-host interactions analysis. (a) Microstructure trap-based single-cell capture array for investigating acid-dependent membrane fusion of vaccinia virions. Reproduced with permission [51]. Copyright 2013, Biophysical Society. (b) A microwell array device for quantitative profiling of innate immune activation by viral infection in single cells. Reproduced with permission [56]. Copyright 2017, Royal Society of Chemistry. (c) Single-cell analysis of the activity of antiviral agents on a multilayer device equipped with microvalves and microstructure traps. Reproduced with permission [57]. Copyright 2019, The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science, under the CC BY-NC license. (d) An automated valve-based microfluidic system with cell-trapping microstructures for multi-parameter and ultra-sensitive protein/mRNA measurements. Reproduced with permission [81]. Copyright 2019, The Authors, under the CC BY license. (e) High-thought screening and enrichment of cells secreting virus neutralizing antibodies on a droplet-based microfluidic system. Reproduced with permission [87]. Copyright 2022, Royal Society of Chemistry, under the CC BY-NC license.
Figure 4
Multi-cell culture microfluidic platforms for viral events analysis. (a) A customized device supporting synchronously culture of different cell lines for rapid susceptible cells screening. Copied with permission [22]. Copyright 2022, The Authors, published by Elsevier B.V. on behalf of KeAi Communications Co. Ltd., under the CC BY-NC-ND license. (b) An impedance-based microfluidic chip for rapid infectivity assay. Reproduced with permission [23]. Copyright 2021, Royal Society of Chemistry, under the CC BY license. (c) A co-culture microchip designed to monitor viral and cellular proteins transfer between infected non-neuronal cells and human axons. Reproduced with permission [102]. Copyright 2015, Grigoryan et al., under the CC BY License. (d) A multi-shear microfluidic platform for exploring the effect of shear stress on vaccina virus-induced cell migration. Reproduced with permission [41]. Copyright 2017, Royal Society of Chemistry. (e) Site-specific labeling of baculovirus on an integrated microfluidic system. Copied with permission [110]. Copyright 2013, Royal Society of Chemistry.
Figure 5
Organ-on-a-chip for viral infectious diseases modeling. (a) Microengineered human alveolus chip for modeling SARS-CoV-2-induced lung injury. Copied with permission [27]. Copyright 2020, The Authors, published by Wiley-VCH GmbH, under the CC BY license. (b) Distal renal tubular system-on-a-chip for exploring the pathogenesis of influenza A virus-induced kidney injury. Reproduced with permission [125]. Copyright 2023, Royal Society of Chemistry. (c) A human lymphoid follicles chip for exploring seasonal influenza vaccination responses. Reproduced with permission [126]. Copyright 2022, The Authors, published by Wiley-VCH GmbH, under the CC BY license. (d) 3D microfluidic liver cultures for hepatitis B virus infection research. Reproduced with permission [135]. Copyright 2018, The Authors, under the CC BY license.
Figure 6
Organ-on-a-chip for viral infectious diseases modeling. (a) Human gut-on-a-chip for modeling enterovirus infection. Reproduced with permission [138]. Copyright 2017, under the CC0 license. (b) A human microvessel-on-a-chip for reproducing Ebola hemorrhagic shock syndrome. Copied with permission [139]. Copyright 2019, The Authors, under the CC BY-NC-ND license. (c) A human blood brain barrier chip for evaluation antivirals against encephalitis virus infection. Reproduced with permission [140]. Copyright 2022, The Authors, under the CC BY license.

 DownLoad:
DownLoad:


 下载:
下载:







 下载:
下载:
