Citation:
Yinglan Yu, Sajid Hussain, Jianping Qi, Lei Luo, Xuemei Zhang. Mechanisms and applications: Cargos transport to basolateral membranes in polarized epithelial cells[J]. Chinese Chemical Letters,
2024, 35(12): 109673.
doi:
10.1016/j.cclet.2024.109673
Mechanisms and applications: Cargos transport to basolateral membranes in polarized epithelial cells
English
Mechanisms and applications: Cargos transport to basolateral membranes in polarized epithelial cells
Received Date:
08 January 2024 Accepted Date:
22 February 2024 Revised Date:
20 February 2024 Available Online:
15 December 2024
Abstract:
In polarized cells, the differential distribution of proteins results in the formation of apical and basolateral membranes. The basolateral membrane contacts basal lamina and mediates cell-to-cell communication, which is crucial for maintaining homeostasis and enabling drug absorption. To establish and maintain the basolateral domain, intricate mechanisms are necessary to ensure the proper sorting and transportation of molecules. Sorting signals play a crucial role in regulating the distributions of basolateral proteins, determining their trafficking route and final residence. Newly synthesized proteins can be segregated into different carrier vesicles at either trans-Golgi network (TGN) or endosomes. Additionally, understanding basolateral transport in polarized epithelial cells is important for predicting diseases and delivering drugs. This review provides a summary of recent advancements in the mechanisms and applications of basolateral sorting and trafficking.
Biological barriers, which are responsible for protecting against foreign objects, are widely distributed throughout the body [1]. The epithelium, an essential component of these barriers, plays a crucial role in maintaining the balance between the organism’s internal environment and the external surroundings [2]. To fulfill this function, the epithelium consists of polarized cells with an asymmetric structure [3]. The epithelial membrane is typically divided into two parts: the apical surface and the basolateral surface, which are separated by intercellular junctional complexes known as tight junctions [4,5]. These polarized epithelial cells have distinct receptors, ion channels, and transporters in their apical and basolateral domains, enabling the directional transport of substances through endocytosis and exocytosis [6]. The apical surface, which faces the external lumen, is equipped with microvilli that aid in the movement of molecules between cells, while the basolateral surface, in contact with neighboring cells, contains spot desmosomes that facilitate cell-to-cell communication [7].
The development and maintenance of biochemical polarity are crucial for the normal physiological functions of epithelium, such as respiratory, digestive, and genito-urinary epithelium [8]. This requires differential cargo trafficking and sorting to proper membrane domains in polarized epithelial cells. The biogenesis of the apical membrane has been extensively studied [9-11]. In addition, complex sorting to the basolateral membrane for transcytosis in epithelial cells is necessary for the absorption of exogenous substances. This review will discuss the trafficking and sorting to the basolateral domains in polarized epithelial cells. Furthermore, this review will summarize the application of basolateral transport in disease prediction and drug delivery, providing new insights for the detection of related diseases and the design of mucosal drug delivery systems.
2.
Basolateral transporting mechanisms
To generate the asymmetric membrane of epithelium, it is necessary to transport the apical and basolateral proteins and lipids from their site of synthesis to their correct final destination. A longstanding objective in studying trafficking in polarized cells is to uncover the mechanisms involved in targeting newly synthesized lipids and proteins to the appropriate plasma membrane domains, as well as to understand the basolateral transport pathways in polarized cells.
2.1
Trafficking routes in polarized epithelial cells
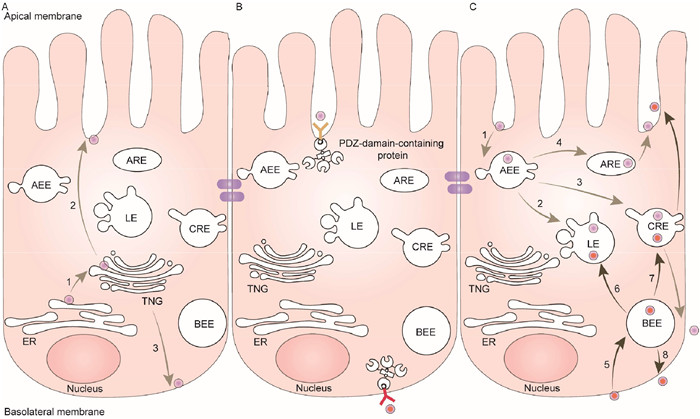
The Golgi apparatus, which is responsible for sorting newly synthesized proteins, is subdivided into three networks: cis-, medial- and trans-Golgi [12]. In epithelial cells, the trans-Golgi network (TGN) is considered the primary sorting station where proteins are assigned to specific membrane locations or endosomal compartments [13,14]. Additionally, polarized epithelial cells have distinct endosomal compartments that play a role in the trafficking and sorting of apical or basolateral cargos [15,16]. These compartments include apical early endosomes (AEE), basolateral early endosomes (BEE), apical recycling endosomes (ARE), and common recycling endosomes (CRE), all of which are involved in substance transport [17,18]. Cargos entering these organelles can be sorted to the apical surface, lysosomes, basolateral surface, or TGN. The distribution of components in the apical and basolateral domains in polarized epithelial cells involves three main processes (Fig. 1) [5,19].
Figure 1
Figure 1.
Major trafficking routes in polarized epithelial cells. (A) Biosynthetic routes to the apical and basolateral membranes, respectively. 1: Proteins are synthesized in the rough ER and transported to the TGN; 2: proteins are delivered to the apical surface; 3. proteins are delivered to the basolateral surface. (B) Proteins are selectively retained at the plasma membrane via interaction with PDZ-domain-containing proteins. (C) The trafficking of internalized components. 1: Apical endocytosed molecules are delivered to AEE; 2: apical endocytosed molecules are transported from AEE to late endosome (LE); 3: apical endocytosed molecules are transported from AEE to CRE and then released to the basolateral surface; 4: apical endocytosed molecules are transported from AEE to ARE and then recycled to the apical surface; 5: basolateral endocytosed molecules are delivered to BEE; 6: basolateral endocytosed molecules are transported from BEE to LE; 7: basolateral endocytosed molecules are transported from BEE to CRE and then released to the apical surface; 8: basolateral endocytosed molecules are recycled from BEE to the basolateral surface.
First, proteins are synthesized in the rough endoplasmic reticulum (ER) and transported to the TGN, where they are packaged into transport vesicles and subsequently delivered to either the apical or basolateral membrane [20,21]. During this transportation process, the cargo adaptor plays a vital role as it is recruited to the TGN and is responsible for facilitating the delivery of cargo. For instance, the cargo adaptor exomer complexes are essential for TGN-plasma membrane trafficking, while the adaptor protein (AP) complexes are required for Golgi-endosomes and Golgi-plasma membrane trafficking [22,23].
Second, certain cargos are selectively retained at the plasma membrane through interaction with the postsynaptic density-95, discs-large, zona occludens 1 (PDZ)-domain-containing proteins. PDZ domains, which are common structural domains consisting of approximately 80 amino acids, play a role in organizing receptors and their downstream effectors to transport cargo. They achieve this by specifically recognizing short C-terminal peptide motifs and internal sequences [24]. For instance, in C. elegans, the PDZ-domain-containing proteins LIN-2, LIN-7, and LIN-10 form a conserved heterotrimeric complex that is crucial for delivering proteins to their proper subcellular locations [25,26]. These C. elegans proteins LIN-2, LIN-7, and LIN-10, are essential for localizing the epidermal growth factor receptor (EGFR)-like tyrosine kinase receptor LET-23 to the basolateral membrane of vulval precursor epithelia. The absence of LIN-2, LIN-7, or LIN-10 results in the loss of LET-23-dependent signaling and a defect in vulval induction [27,28]. In polarized Madin-Darby canine kidney (MDCK) cells, galectin-3 has been shown to associate with the apical membrane component lactase-phlorizin hydrolase (LPH) in a glycoprotein-dependent manner [29,30]. Depletion of galectin-3 leads to mis-sorting of LPH to the basolateral surface. Furthermore, the presence of galectin-3 is necessary for the formation of clusters of glycoproteins in the apical domain. Interestingly, another member of the galectin family, galectin-4, interacts with apical domain sulfatides to sort proteins in intestinal HT29 cells [31,32]. Additionally, several other membrane proteins, including annexin 2, annexin 13b, myelin, and lymphocyte (VIP17/MAL), are believed to be involved in apical sorting [33-35].
Third, some internalized components are transported to early endosomes, from where they can be transferred to late endosomes, recycled back to the apical surface, or delivered to the opposite surface [19]. For instance, the transferrin receptor (TfnR) is a well-characterized recycling receptor in epithelial cells [36]. Upon specific targeting and binding to TfnR, the clustering of TfnR occurs with clathrin-coated pits. These coated pits then bud into the membrane and are transported to early endosomes. Finally, the endosome-derived vesicles recycle TfnR to the epithelial surface [37]. In epithelial cells, the polymeric immunoglobulin receptor (pIgR) uptakes its ligand polymeric immunoglobulin A (pIgA) at the basolateral membrane. The internalized molecules pass through a series of endosomal compartments, such as basolateral early endosomes and common endosomes. Within these compartments, some cargoes are sorted back to the basolateral side, some to late lysosomes, and others to the apical side [38].
2.2
Basolateral sorting mechanisms
Basolateral sorting of proteins is primarily conducted through the interaction of sorting signals with sorted adaptor complexes (Table 1) [3].
Table 1
Table 1.
Mechanisms for sorting to the basolateral membranes [3,23].
In polarized cells, the sorting of receptors and secreted proteins to the basolateral domain is primarily directed by sorting signals [39,40]. These sorting signals are assisted by endocytic signals, which play a role in polarized endocytosis and recycling. The most extensively studied sorting signals involved in basolateral sorting are tyrosine-based sorting motifs (YXXφ or NPXY), where X can be any amino acid and φ represents an amino acid with a bulky hydrophobic residue [41]. These sorting motifs are located in the cytosol-facing domains of proteins. The low-density lipoprotein receptor (LDLR) serves as a typical example. LDLR, a receptor localized in the basolateral domain of epithelial cells, contains two tyrosine-based sorting motifs in the proximal membrane for endocytosis, as well as a C-terminal motif in the cytoplasmic domain for Golgi-basolateral delivery. The proximal membrane motif, consisting of two tyrosine-based domains, is necessary for polarized endocytosis, while the C-terminal region, which includes a tyrosine-dependent domain, encodes a sorting signal for the basolateral membrane [36,42]. Additionally, the Nipah virus possesses two basolateral sorting signals, which include a di-tyrosine motif at position 28/29 in the G protein and tyrosine 525 in the F protein [43].
Dileucine-based sorting motif ([DE]XXXL[LI]) is another identified basolateral sorting signal [44]. This sequence is located in the cytosol-facing domains of many trans-membrane proteins in polarized epithelial cells [45]. In T cells, surface sorting of the Q-peptide from surface antigen protein CD4 requires two leucine residues [46]. Additionally, single leucine residue-based motifs have also been identified for basolateral sorting [7,47]. For example, the basolateral sorting of CD147 is mediated by signals consisting of a single leucine residue [48].
However, some basolateral cargos contain noncanonical sorting signals. For example, the basolateral sorting region that involves a 25-residue motif is located in the C-terminal tail of the P2Y(1) receptor. In this case, the basolateral sorting determinant is identified as the total number of charged residues [49]. The N-terminal domain within syntaxin 4 has also been identified, where a short stretch between residues 24 and 29 is found to be crucial for basolateral sorting [50]. Additionally, the sorting signals critical for the basolateral targeting of the receptor tyrosine kinase ERBB2 reside between Gln-692 and Thr-701 [51]. The basolateral sorting of polymeric immunoglobulin receptors depends on their 17-juxtamembrane cytoplasmic domains, with three residues (His656, Arg657, Val660) in this region identified as the basolateral targeting signal [52]. In the case of epidermal growth factor receptors, the basolateral sorting signals are L658L659, P667XXP670, and Arg662 residues [53]. Furthermore, the basolateral sorting signal of the TfnR is the noncanonical motif GNDS [54].
2.2.2
Basolateral sorting adaptor complexes
To ensure precise determination of the destination by basolateral sorting signals, the AP-clathrin complexes play an essential role in specific cargo trafficking. The AP consists of two large domains (100–160 kD), one medium domain (50 kD), and one small domain (20 kD). The appendage domains, formed by the C-terminal regions of the two large domains, bind accessory proteins such as AP180 and epsins. On the other hand, the N-termini, composed of medium and small domains, form the core or trunk domains that bind cargo molecules, phospholipids, and Arf proteins [55].
A total of five types of AP complexes have been identified, three of which (AP-1, AP-3, and AP-4) play a crucial role in protein sorting. AP-1 and AP-3 contain clathrin box motifs in their flexible linker regions and appendage domains, while AP-4 operates independently of clathrin. The medium domain of AP-1 has two isoforms, µ1A (AP-1A) and µ1B (AP-1B), both of which are essential for basolateral targeting. AP-3 also consists of two isoforms, AP-3A and AP-3B, with AP-1A and AP-3A being expressed in all cell types, AP-1B expressed only in epithelial cells, and AP-3B expressed exclusively in neuroendocrine and neuron tissues [47,56]. AP-1 facilitates the sorting of endosomes and basolateral surfaces, AP-3 mediates sorting in the TGN and/or endosomes, while AP-4 is involved in basolateral sorting in polarized epithelial cells [22].
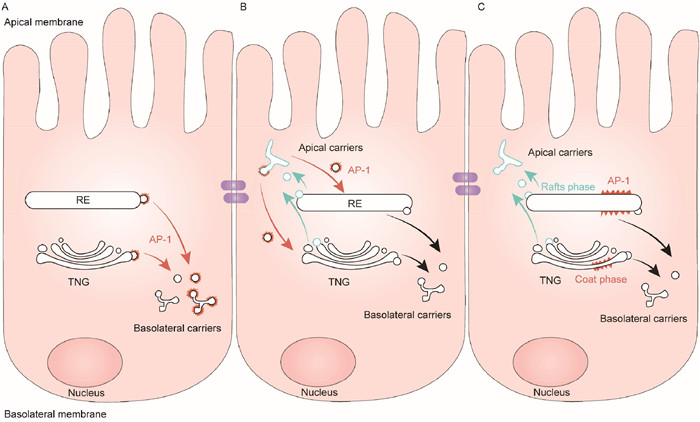
In polarized epithelial cells, such as Caco-2, HT29, DMCK, RL95–2, and Hec-1-A cells, AP-1 plays a crucial role in the trafficking of basolateral protein. Specifically, AP-1A is involved in protein sorting at the TGN and endosomes, while AP-1B regulates the recycling of proteins from recycling endosomes (REs) back to the basolateral membrane (Fig. 2) [47,57]. Both AP-1A and AP-1B contribute to the trafficking and sorting of polarized proteins with their roles being complementary. For example, the simultaneous knockdown of AP-1A and AP-1B leads to more severe defects in the basolateral sorting of TfnR and adenovirus receptors compared to individual knockdown [58]. Furthermore, AP-1B facilitates the export of the LDLR from the TGN in the absence of AP-1A [59]. In certain cells, AP-1B directly binds to basolateral sorting signals of cargo, which are then trafficked in an AP-1B-dependent manner. Notably, AP-1B exhibits approximately fivefold higher binding affinity to the noncanonical motifs of LDLR compared to AP-1A [60]. Efficient basolateral targeting of the poliovirus receptor and the interleukin-6 receptor β chain relies on AP-1B, while AP-1A is not involved [59]. Both AP-1A and AP-1B interact with tyrosine- or dileucine-based signals and determine their basolateral targeting [61,62]. Although there are some overlapping preferences in cargo recognition between AP-1A and AP-1B, AP-1A primarily localizes to the TGN, while AP-1B predominantly localizes to the REs [59,60,63]. Notably, AP-1B is specifically regulated by the phosphoinositide phosphatidylinositol 3,4,5-trisphosphate (PIPI3) and the small guanosine triphosphate (GTP)-binding protein Arf6 [64,65].
Figure 2
Figure 2.
The schemes depict models for the function of AP complexes in basolateral sorting at TGN and RE. (A) AP-1 sorts cargos into pleiomorphic transport carriers at the TGN/RE; then, these vesicles lose their coats and eventually deliver their cargos to the basolateral domains. (B) AP-1 removes basolateral cargos from apical carriers to return to the TGN/RE. (C) AP-1 segregates basolateral cargos into a "coat phase" of the TGN/RE, away from the “raft phase” that gives rise to apical carriers.
AP-4, the cargo adaptor for non-clathrin-coated vesicles at TGN, also plays a crucial role in basolateral polarity [66]. In MDCK cells, AP-4 binds to the cytosolic tails of CD-MPR and LDLR, directing them to the basolateral membrane. Interestingly, when the µ4 subunit of AP-4 is knocked down, these cargos are mislocalized to the apical membrane [66]. However, it is worth noting that knockout mice lacking the µ4 subunit of AP-4 do not display significant defects in polarized epithelial cells [67,68]. Instead, these mice exhibit mistargeting of certain neuronal membrane proteins, such as the dendritic Delta2 glutamate receptor and dendritic AMPA-type glutamate receptor [69]. Moreover, mutations in the gene encoding the µ4 subunit of AP-4 lead to a severe neurodevelopmental disorder in humans, characterized by intellectual disability, cerebral palsy, and microcephaly [70]. Further investigations have revealed that the µ4 subunit of AP-4 specifically binds to the tyrosine and phenylalanine residues in the cytosolic tails of the Delta2 glutamate receptor [71].
There are additional adaptors involved in basolateral trafficking and sorting. One example is the reduction of ankyrin (APs) expression, which leads to decreased sorting of E-cadherin from the TGN to the basolateral surface [72,73]. In yeast, the exomer serves as another adaptor that directs the traffic of chitin synthase Chs3p between the TGN and plasma membrane [74,75]. Another adaptor, sorting nexin SNX17, interacts with the IXNPXY motif in the LDLR-related protein LRP1 tail and regulates the recycling of LRP1 from endosomes to the basolateral membrane [76,77].
2.3
Cargo trafficking to the basolateral membrane
The release of transport cargos carrying basolateral proteins from the TGN or REs must be coupled to specify vesicle trafficking and fusion with target plasma membrane domains during the generation and maintenance of epithelial polarity. These processes involve microtubules, actin, and relevant protein complexes (Fig. 3) [78].
Figure 3
Figure 3.
Basolateral sorting and trafficking. Proteins in TGN are sorted into various vesicles through the interaction of sorting motifs and adaptor complexes. These vesicles are delivered to basolateral domains along cytoskeletal elements, which are regulated by various vesicle-tethering and SNARE complexes.
In polarized cells, the microtubule and actin cytoskeletons exhibit specialized organizations. Generally, microtubules are bundled along the apico-basal axis in epithelial cells, which represents the cell’s long axis. On the other hand, actin filaments are distributed underneath the entire plasma membrane [72,79]. Microtubules primarily regulate the efficiency of cargo trafficking to the plasma membrane in polarized epithelia, while actin meshworks are responsible for cargo formation, scission, fusion, and transportation [80-82]. During these processes, distinct microtubules and actin motors drive trafficking vesicles toward the basolateral surface [57]. For instance, plus-end microtubule kinesin motors participate in basolateral protein trafficking [83]. Additionally, myosin VI, an actin-based motor protein, appears to have a role in basolateral trafficking [84]. In MDCK cells, myosin VI is essential for sorting and trafficking newly synthesized basolateral surface proteins to their correct membrane domain [85].
2.3.2
GTPase-mediated export
The Arf, Rab, and Rho subfamily members of GTPases function as molecular switches that facilitate the formation and trafficking of vesicles in TGN [86,87]. These GTPases regulate vesicle budding, fission, and transportation via microtubules and actin [88]. The Rho and Rab GTPase families are specifically involved in basolateral trafficking [89,90]. For instance, Rab6 binds to kinesin and myosin motors to regulate the transport of vesicles from the TNG to the cell periphery [91]. Furthermore, vesicle-tethering complexes play a crucial role in exocytic pathways, enhancing the specificity and efficiency of cargo transport. One such complex is the exocyst complex, which consists of at least six proteins and is distributed between the plasma membrane and the trafficking cargo. In epithelial cells, the exocyst complex appears to facilitate the docking of a subset of basolateral vesicles at the plasma membrane [92]. The assembly of the tethering complex involves the Rab GTPase family. Specifically, Rab3 is located on synaptic vesicles, Rab8 is found on basolateral vesicles, and Rab10 is localized in the Golgi during early cell polarization [93,94]. In MDCK cells, Rab8 is involved in the exocytosis of cargo from REs to the basolateral surface [95]. Rab10, on the other hand, regulates transport from the Golgi to the basolateral membrane and may work in conjunction with Rab8 [96].
2.3.3
SNARE complexes-mediated export
Soluble N-ethylmaleimide sensitive factor attachment protein receptor (SNARE) complexes also play a role in basolateral trafficking, specifically in the transport of vesicles between the TNG and the basolateral membrane [97,98]. Recently, a new mechanism of basolateral trafficking has been described, involving a protein complex called factors for endosome recycling and Rab interactions (FERARI) [99]. FERARI is responsible for tethering, SNARE interactions, binding to sorting nexins (SNXs), scaffolding and protein-protein interactions, membrane tubule stabilization, and pinching. In epithelial cells, the recycling of TfnR and glucose transporter-1 (GLUT1) relies on the FERARI-mediated kiss-and-run mechanism. This process is facilitated by the FERARI tethering complex, which enhances the interaction between tubular SNX1-sorting endosomes and Rab11-positive recycling vesicles. Impairment of FERATI function leads to a significant reduction in the recycling of trafficking and sorting cargoes [100].
2.3.4
Polarized vesicle fusion at membrane domains
The polarized localization of membrane proteins can be influenced by the asymmetric stabilization of the plasma membrane domain. Proteins that are sorted in exocytic pathways can undergo further refinement through selective stabilization [101]. Ankyrin-spectrin, a cytoplasmic complex, binds to several membrane proteins such as cell-adhesion proteins, receptors, ion transporters, and channels [72]. In epithelia, ankyrin-spectrin complexes are found in different locations on the basolateral domain [98]. By binding to these scaffolds, membrane proteins involved in specific biological processes can increase their accumulation and residence time, thereby enhancing their function [102]. For instance, the ankyrin-spectrin complex binds to the T-tubule clustering of Na+/Ca2+-exchanger, Na+/K+-ATPase, and inositol 1,4,5-trisphosphate receptor (IP3R) [72].
2.3.5
Protein kinase-mediated export
Protein kinases and phosphatases play a role in the formation of secretory cargo at the TGN [103]. The function of membrane recruitment and cytosol release is mediated by the cyclic phosphorylation and dephosphorylation of AP-1. During the process of AP-1 recruitment to the membrane, protein phosphatase 2A regulates the dephosphorylation of the AP-1 µ1 subunit, which facilitates clathrin assembly. Once on the surface, the µ1 subunit of AP-1 is phosphorylated, leading to a conformational change and promoting binding to the sorting signal [104]. Protein kinase (PKD), consisting of PKD1, PKD2, and PKD3 isoforms, is a crucial regulator of TGN carrier fission [105]. In polarized cells, PKD1 and PKD2 are involved in the exit of proteins from the TGN, destined for the basolateral surface [106].
2.4
Basolateral trafficking vesicles
Several common proteins, including Na+/K+-ATPase, vesicular stomatitis virus-glycoprotein (VSV-G) protein, and E-cadherin, are sorted to the basolateral surface [106]. Na+/K+-ATPase can bind to APs called ankyrins and be transported from the TNG to the basolateral membrane [72]. VSV-G contains tyrosine-based sorting signals that are recognized by AP1 clathrin coat adaptor complexes and sorted through Rab8-positive CREs [94]. In the case of E-cadherin-containing vesicles, the exocyst recruits PARD3 to the plasma membrane through its interaction with phosphatidylinositol 4,5-bisphosphate PI(4,5)P2, thereby serving as the plasma membrane trafficking receptor [107].
Exosomes are small extracellular vesicles that are released by cells [108-110]. These cells release two distinct types of exosomes: apical exosomes from their apical membrane and basolateral exosomes from their basolateral membrane. According to Matsui et al., MDCK cells release basolateral exosomes through a sphingomyelinase-dependent ceramide production machinery [111]. Their subsequent research reveals that the release of basolateral exosomes is regulated by Rab39. Specifically, Rab39 interacts with its effector uveal autoantigen with coiled-coil domains and ankyrin repeats (UACA), and recruits the component of the BLOC-1-related complex (BORC) lyspersin. The Rab39-UACA-BORC complex plays a role in regulating basolateral exosome trafficking and release [112].
3.
Applications of basolateral trafficking
Proper basolateral sorting is crucial for maintaining the homeostasis of polar epithelial cells. In disease pathology, various defects in trafficking can occur, such as inverted polarity, intracellular retention, and disrupted recycling. Additionally, basolateral sorting signals can facilitate unidirectional transcytosis from the apical to the basolateral side. Hence, basolateral trafficking plays a significant role in predicting diseases and designing drug delivery systems (Fig. 4).
Figure 4
Figure 4.
The applications of basolateral trafficking in disease prediction and drug delivery.
The establishment and maintenance of basolateral sorting and trafficking are crucial for maintaining homeostasis. Perturbations in basolateral trafficking have been observed in various diseases [113]. For example, in Liddle syndrome, mutations lead to a downregulation of sodium channels in epithelial cells [114]. Similarly, mutations in proximal tubule cells can cause Dent disease by redistributing H+-ATPase to the basolateral surface [115,116]. Basolateral exosomes derived from proximal tubular epithelial cells play a significant role in modulating tubulointerstitial inflammatory responses seen in immune-mediated kidney diseases [117]. Recently, it has been reported that basolateral inflammatory exosomes are increased in the circulation of patients with chronic kidney disease, suggesting their potential diagnostic utility in the future [118]. Epithelial polarity changes occur when neoplastic cells arise, causing tumor necrosis factor receptors to localize on the basolateral surface. This localization allows it to bind ligands and subsequently trigger apoptotic signaling [119]. Additionally, aberrant basolateral trafficking is observed in infected mucosal epithelium and alcohol use disorder [120-122]. The variation in epithelial compartmentalization of ligands and receptors can be used as a general marker to generally monitor tissue homeostasis.
3.2
Drug delivery
The basolateral sorting and trafficking mechanism can also be utilized for drug delivery [123-128]. For example, nanoparticles modified with butyrate (Bu-NPs) can bind to monocarboxylate transporter-1 (MCT-1) in intestinal epithelial cells. Leptin can enhance the insertion of MCT-1 into the basolateral membrane of epithelial cells, thereby improving the transport of Bu-NPs [129,130]. Similarly, nanoparticles modified with fructose undergo apical endocytosis and basolateral exocytosis through glucose transporter type 2 (GLUT2). When co-administered with a sweetener, acesulfame potassium, it can increase the apical distribution of GLUT2 in intestinal epithelial cells, facilitating the unidirectional transcytosis of fructose-functionalized nanoparticles [131]. Wang et al. developed an oral insulin delivery system modified with a Golgi apparatus targeting peptide, which can migrate to the basolateral membrane and traverse the intestinal epithelium [132]. Zhang et al. demonstrated that micelles modified with basolateral membrane sorting signal mainly accumulate in vesicles near the basolateral surface, while micelles modified with TGN sorting signal peptide exhibit superior colocalization with TGN, transcytosis ability, and in vivo absorption [133]. In the design of active targeting nanoparticles to overcome the polarized epithelial barrier, it is important to consider receptor characteristics and ligand density. Firstly, it is preferable for receptors to be transferred through CRE or the Golgi complex for efficient transcytosis. Secondly, proper ligand density is crucial for achieving the best transcytosis. Thirdly, the delivery of active-targeting nanoparticles with the appropriate ligand density may follow the receptor’s itinerary, while nanocarriers with high ligand density may alter the transport pathway [134]. Additionally, the use of tailored polymers and appropriate rigidity can improve ligand targeting and enhance transcytosis [135-141]. Further investigations have suggested that the polarity of epithelial cells plays a crucial role in the transportation of active-targeting nanocarriers, and this transportation process may also impact the polarity by altering the expressions of vesicle-related proteins [142].
Unidirectional transport can be achieved by utilizing the different microenvironments of the apical and basolateral sides of polarized epithelium. For example, the apical side of the intestinal epithelium is acidic (pH 6.0–6.5), while the basolateral side is neutral (pH 7.4). Intestinal epithelial cells express high levels of the neonatal Fc receptor (FcRn), which can selectively bind its ligands at acidic pH but not at neutral pH. Consequently, substances such as peptides, immunoglobulin G (IgG), or albumin modified nanoparticles can effectively bind to FcRn at the apical surface of epithelial cells, facilitating their internalization. These substances can then be released into the extracellular space at the basolateral side, resulting in apical-to-basolateral unidirectional transport [143-145]. Furthermore, milk-derived exosomes are also capable of undergoing basolateral exocytosis and traversing the intestinal epithelium while adapting to the pH variations within the intestinal environment [146]. Yang et al. designed ligand-switchable nanoparticles that were modified with a pH-triggered stretchable peptide. After oral administration, the peptide adopts a stretched conformation in acidic environments, which mediates efficient transcytosis in the intestinal epithelium. Upon entering systemic circulation, the peptide folds at physiological pH, allowing for basolateral transport [147]. Zheng et al. fabricated zwitterionic hydrogel nanoparticles for enhanced oral insulin delivery and found that these nanoparticles are prone to undergo basolateral exocytosis. Further investigation revealed that peptide transporter 1 (PEPT1) facilitates the apical internalization of zwitterionic nanoparticles due to their higher binding affinity at pH 6.5, while basolateral peptide transporter (BPT) mediates basolateral exocytosis at pH 7.4, resulting in efficient apical-to-basolateral transport [148].
4.
Conclusions
Basolateral sorting and trafficking in polarized cells are crucial for establishing and maintaining cell polarity, which is essential for homeostasis. Polarized cells utilize intricate sorting and trafficking systems to regulate the distribution of proteins to their basolateral membrane. The key factors controlling basolateral membrane sorting include the cell compartments, sorting signals, and regulators that have been identified. The biosynthesis and separation of cargo have been observed to involve the endoplasmic reticulum, Golgi, and endosomes. However, the underlying mechanisms of these complex processes are still not fully understood. Further research is needed to comprehensively understand trafficking routes, sorting signals, sorting mechanisms, as well as the role and function of Golgi, endosomes, and cytoskeletal elements. Future studies will uncover and define basolateral sorting signals and mechanisms. Moreover, additional research is necessary to explore how the basolateral trafficking pathways adapt to the physiology of different tissues. Considering the significance of polarity in normal tissue function, these investigations have important implications for understanding the pathogenesis, prediction, and therapy of diseases. Additionally, basolateral transport can facilitate the transcytosis of drugs and provide new insights for the design of mucosal drug delivery systems.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
This study is supported by the China Postdoctoral Science Foundation (No. 2023M740731), Fundamental Research Funds for the Central Universities (No. SWU-KQ22024), National Key Research and Development Program of China (No. 2021YFD1800900), National Natural Science Foundation of China (No. 82073790), the Chongqing Science and Technology Commission (No. CSTB2022TIAD-LUX0001).
[1]
K.S. Doran, A. Banerjee, O. Disson O, M. Lecuit, CSH Perspect. Med. 3 (2013) a010090. doi: 10.1101/cshperspect.a010090
[2]
H. Malhaire, J.C. Gimel, E. Roger, J. Benoît, F. Lagarce, Adv. Drug Deliv. Rev. 106 (2016) 320–336. doi: 10.1016/j.addr.2016.03.011
M.P. Lisanti, M. Sargiacomo, L. Graeve, A.R. Saltiel, E. Rodriguez-Boulan, Proc. Natl. Acad. Sci. U. S. A. 85 (1988) 9557–9561. doi: 10.1073/pnas.85.24.9557
[12]
J. Klumperman, CSH Perspect. Biol. 3 (2011) 839–848.
Figure 1
Major trafficking routes in polarized epithelial cells. (A) Biosynthetic routes to the apical and basolateral membranes, respectively. 1: Proteins are synthesized in the rough ER and transported to the TGN; 2: proteins are delivered to the apical surface; 3. proteins are delivered to the basolateral surface. (B) Proteins are selectively retained at the plasma membrane via interaction with PDZ-domain-containing proteins. (C) The trafficking of internalized components. 1: Apical endocytosed molecules are delivered to AEE; 2: apical endocytosed molecules are transported from AEE to late endosome (LE); 3: apical endocytosed molecules are transported from AEE to CRE and then released to the basolateral surface; 4: apical endocytosed molecules are transported from AEE to ARE and then recycled to the apical surface; 5: basolateral endocytosed molecules are delivered to BEE; 6: basolateral endocytosed molecules are transported from BEE to LE; 7: basolateral endocytosed molecules are transported from BEE to CRE and then released to the apical surface; 8: basolateral endocytosed molecules are recycled from BEE to the basolateral surface.
Figure 2
The schemes depict models for the function of AP complexes in basolateral sorting at TGN and RE. (A) AP-1 sorts cargos into pleiomorphic transport carriers at the TGN/RE; then, these vesicles lose their coats and eventually deliver their cargos to the basolateral domains. (B) AP-1 removes basolateral cargos from apical carriers to return to the TGN/RE. (C) AP-1 segregates basolateral cargos into a "coat phase" of the TGN/RE, away from the “raft phase” that gives rise to apical carriers.
Figure 3
Basolateral sorting and trafficking. Proteins in TGN are sorted into various vesicles through the interaction of sorting motifs and adaptor complexes. These vesicles are delivered to basolateral domains along cytoskeletal elements, which are regulated by various vesicle-tethering and SNARE complexes.

 DownLoad:
DownLoad:




 下载:
下载:





 下载:
下载:
