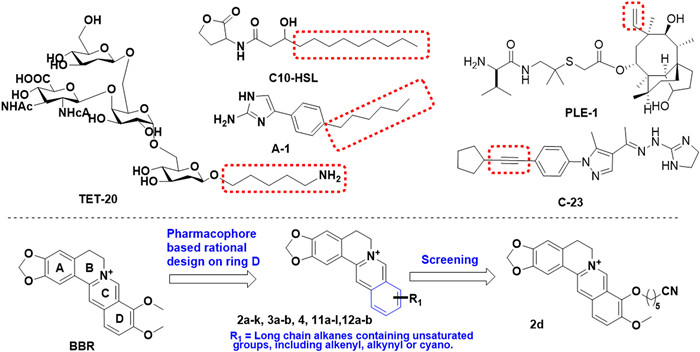
Scheme 1.
Chemical structures of antibiotic adjuvants against A. baumannii, and structure modification strategy on BBR.
Novel berberine derivatives as adjuvants in the battle against Acinetobacter baumannii: A promising strategy for combating multi-drug resistance
Liping Zhao , Xixi Guo , Zhimeng Zhang , Xi Lu , Qingxuan Zeng , Tianyun Fan , Xintong Zhang , Fenbei Chen , Mengyi Xu , Min Yuan , Zhenjun Li , Jiandong Jiang , Jing Pang , Xuefu You , Yanxiang Wang , Danqing Song

Acinetobacter baumannii (A. baumannii) is a Gram-negative opportunistic nosocomial pathogen, which is responsible for a variety of hospital-acquired and community-acquired infectious diseases, such as pneumonia, urinary tract infections, and surgical site infections, particularly among patients in intensive care units. Drug resistant A. baumannii, especially carbapenem-resistant A. baumannii (CRAB) has been ranked as the most critical pathogens by World Health Organization (WHO) due to the lack of effective and safe treatment options [1-5]. The development of new antibacterial or antibiotic adjuvants is critically needed. Given the lengthy development cycle for novel antibiotics, exploring novel antibacterial adjuvants to enhance the efficacy of first-line antibiotics is considered as an effective solution for bridging the widening gap between clinical demands and drug innovation. Antibiotic adjuvants, showing little or no antimicrobial activity alone generally [6-10], could enhance antibiotic activity either by blocking resistance or by boosting the host response to infection. Several adjuvants are already in clinical use, such as avibactam, which inhibits the β-lactamases but exhibits no antimicrobial activity alone.
Aztreonam (ATM) is the only monobactam in clinical use, and has been a commonly prescribed antibiotic in clinical practice since 1986, primarily for the treatment of Gram-negative bacterial infections. However, ATM exhibits limited efficacy against A. baumannii, mainly due to β-lactamase production and efflux pump overexpression. Berberine (BBR), a traditional Chinese medicine isoquinoline alkaloid, has been used clinically as an antidiarrheal agent for more than 2000 years [11-14], with low general toxicity, genetic toxicity, and mutagenicity in vitro and in vivo. A moderate synergistic effect between BBR and ATM against A. baumannii was demonstrated, which provided further insights for the future development of BBR or its derivatives as an antibiotic adjuvant [15,16]. As of present, there is no literature available regarding the synergistic antibacterial activity of BBR derivatives with antibiotics against A. baumannii.
Based on a rational design strategy, we initiated an in-depth structural analysis of the most recently discovered agents exhibiting anti-A. baumannii properties or synergistic effects. These synergistic enhancers share a common alkyl chain pharmacophore (Scheme 1), including C10-HSL (a quorum sensing inhibitor of A. baumannii) [17], compound A-1 (aminimizole-based small molecule adjuvant suppresses A. baumannii resistance by down-regulating the PmrAB system) [18], and an oligosaccharide TET-20 (a candidate vaccine adjuvant against A. baumannii) [19]. Moreover, recently identified novel candidates exhibiting anti-A. baumannii properties, such as the pleuromutilin analogue PLE-1 [20] and the benzopyrazole compound C-23 [21], feature unsaturated moieties such as alkenyl and alkynyl groups. At present, the reported structural modifications of BBR derivatives with antibacterial activity mainly focus on the ring C or D, and derivatives resulting from C-ring modifications (positions 7, 8, and 13) of BBR typically exhibit enhanced activity against Gram-positive bacteria [22,23]. Conversely, modifications targeting the D-ring (positions 9 and 12) may yield more promising candidates with activity against Gram-negative bacteria [24,25]. Therefore, the structural modifications in this study predominantly focused on the D-ring. These findings serve as a foundation and guidance for subsequent modifications of BBR derivatives.
Herein, based on the summary of pharmacophore which is beneficial to improve anti-A. baumannii synergistic activity, thirty novel BBR derivatives with unsaturated moieties containing alkyl chain pharmacophore on ring D were rationally designed and synthesized (Scheme 1). The synthetic route was described in Schemes S1–S3 (Supporting information). The antimicrobial activities of all the newly synthesized compounds used alone and in combination with ATM against A. baumannii ATCC 19606, a reference strain commonly used in antibacterial activity evaluation, were evaluated via the broth microdilution method and checkerboard analysis, respectively, taking BBR and levofloxacin as the positive controls. The minimum inhibitory concentration (MIC) value of levofloxacin against the ATCC 19606 strain is 0.25 µg/mL (Table S1 in Supporting information), indicating the reliability of the experimental results. The combined effect of all target compounds and ATM were indicated by fractional inhibitory concentration index (FICI). The combination was defined to be synergistic when the FICI was ≤0.5; indifferent when 0.5 < FICI < 4; antagonistic when FICI ≥ 4. The MICs and FICIs of the analogues were listed in Table S1 (Supporting information).
The results showed that BBR exhibited a weak inhibitory effect on A. baumannii ATCC 19606 when used alone, with an MIC of 1024 µg/mL. In the case of combination, a synergistic effect was observed, with an FICI of 0.281. The addition of 32 µg/mL BBR led to a significant decrease in the MIC value of ATM from 32 µg/mL to 8 µg/mL. Structure–activity relationship (SAR) study of BBR’s derivatives was first focused on the 9-substituents of BBR, and compounds with ether linker (2a–k) at position 9 were prepared and screened. Out of these compounds, 2a and 2b, which feature 1-butynyl and 1-octynyl substituents, respectively, demonstrated superior synergistic effects than BBR, with FICIs of 0.258 and 0.254. When the alkynyl group was replaced by a cyano group, compounds 2c–e were synthesized and evaluated. Compound 2d exhibited a promising synergistic potency with an FICI value of 0.129. The MIC of ATM against A. baumannii was reduced to 4 µg/mL (1/8 MIC) with the addition of only 4 µg/mL 2d, which was significantly lower than that of BBR (32 µg/mL). Next, compound 2f in conjunction with an internal alkyne showed comparable activity, compared with BBR. Additionally, compounds 2g and 2h, which contained an alkenyl group, displayed improved synergetic effects with the FICI values around 0.25 when compared to the leading compound. Meanwhile, 2i–k with polyethylene glycol linker showed comparable activities to those alkane linker derivatives. An indifferent effect (FICI > 0.5) was observed in the combination of ATM and 3a or 3b with a reduced tetrahydroberberine skeleton, suggesting the π-conjugated structure might play a critical role in improving synergistic antimicrobial potency of BBR analogues. Next, 9-amine BBR analogue 4 was generated and only exhibited moderate activity. These findings highlight the favorable effects of appropriate modification at 9-position in optimizing the synergic activities.
When the same substituent groups were moved to position 10, compounds 11a–l, 12a and 12b were generated, and they exhibited comparable or decreased activities compared to their counterparts. Among these compounds, 11h and 11i, featured alkenyl substituents at position 10, displayed better synergetic effects with the FICIs of 0.254 and 0.252. Similarly, declined synergetic potency of 12a and 12b again confirmed the vital role of the conjugated system in improving antimicrobial activity. Finally, a modification was made at the 3-position by selectively opening the methylenedioxy ring to produce compounds 14a and 14b. Both compounds exhibited a significant decrease in synergetic activity, indicating that the methylenedioxy moiety in BBR analogues might be essential for synergistic effects. Consequently, six BBR analogues with the best synergistic antibacterial effects, namely 2b, 2d, 2g, 2h, 11h and 11i were selected as the representative compounds for further investigation.
Six highly active compounds 2b, 2d, 2g, 2h, 11h and 11i were first evaluated for their safety profiles. The MTT assay was used to assess the in vitro cytotoxic effects of these compounds on human large cell lung carcinoma cells H460, human hepatocarcinoma cells HepG2, and human acute monocytic leukemia cells THP1. As shown in Table S2 (Supporting information), compound 2d showed the highest biosafety, with 50% cytotoxicity concentrations (CC50) values of 107.8, 87.75 and 82.45 µmol/L for H460, HepG2 and THP1, respectively. Other compounds containing alkynyl and alkenyl group, including 2b, 2g, 2h, 11h and 11i, displayed relatively higher cytotoxicity. Therefore, compound 2d was chosen as the representative compound for the further investigation.
To preliminarily investigate the pharmacokinetic properties of 2d, the stability assay was performed in human plasma (Bioreclamation IVT, Cat. No. HUMANPLK2P2N, Batch No. HMN761043) in vitro using propantheline bromide, which contains an ester bond, as the control [26]. Compound 2d and propantheline were incubated with human plasma, and samples were collected at 0, 10, 30, 60, and 120 min, respectively. As shown in Table S3 (Supporting information), propantheline was rapidly hydrolyzed in blood within 1 h, as expected. In contrast, compound 2d demonstrated satisfactory stability in plasma, remaining stable for up to 2 h.
To explore whether 2d could enhance the activity of ATM against other A. baumannii strains, one standard strain (ATCC 17978) and thirteen randomly selected clinical isolates, including six carbapenem-susceptible strains and seven carbapenem-resistant A. baumannii strains were used to conduct the checkerboard assay. As shown in Table S4 (Supporting information), the combination of 2d and ATM exhibited a synergistic effect against 11 out of 14 tested A. baumannii strains, with a median FICI of 0.313. The MIC values of ATM against ATCC 17978, 20-6, 21-22 and 21-24 were reduced to 2–4 µg/mL when used in combination with different concentrations of 2d. The CRAB isolates demonstrated higher resistance to ATM (MICs at 64–128 µg/mL) compared to A. baumannii ATCC 19606. Synergistic effects were also observed in the combination of 2d and ATM for all the tested CRAB strains, with FICI values ranging from 0.281 to 0.313. These results suggested that the addition of 2d increased the susceptibility of CRAB strains to ATM. We further evaluated the synergistic effects of the 2d/ATM combination against three strains of extended-spectrum β-lactamases (ESBLs)-producing A. baumannii, including 22-LP37, 22-LP41 and 22-LP42 strains (Table S5 in Supporting information). The FICIs ranged from 0.516 to 0.531, indicating no apparent synergies, which can be explained by the hydrolysis of ATM by ESBLs. Therefore, we continued to explore whether the introducing of β-lactamase inhibitor could restore the synergy of 2d to ATM against ESBL-producing A. baumannii.
Avibactam (AVI) is a non-β-lactam β-lactamase inhibitor that can inhibit almost all serine β-lactamases (SBLs) but not metallo-β-lactamases (MBLs). Due to its stability against MBLs, the ATM-AVI combination may therefore be a viable choice against carbapenem-resistant Enterobacteriaceae (CRE) producing SBLs, MBLs, and both. Large-scale in vitro susceptibility studies have provided substantial evidence for this strategy, and this combination is actively undergoing clinical development for the treatment of serious infections caused by CRE. However, it has been noted that this approach still exhibits limited potencies against A. baumannii. Therefore, the in vitro synergistic antibacterial activity of 2d was evaluated with ATM/AVI against the above mentioned ESBL-producing A. baumannii (22-LP37, 22-LP41 and 22-LP42). The results shown in Fig. 1A and Table S6 (Supporting information) indicated that the combination of 2d (16–32 µg/mL) with ATM/AVI (2:1, concentration ratio) consistently resulted in significantly synergistic effect, with the triple-combination FICIs of 0.145, 0.270 and 0.297 in the three clinical isolates. Considering that ATM is not easily hydrolyzed by MBLs, 2d/ATM/AVI triple-combination is anticipated to be stable to all classes of β-lactamase, and thus, may provide a promising antibacterial effect against β-lactamase producing A. baumannii.
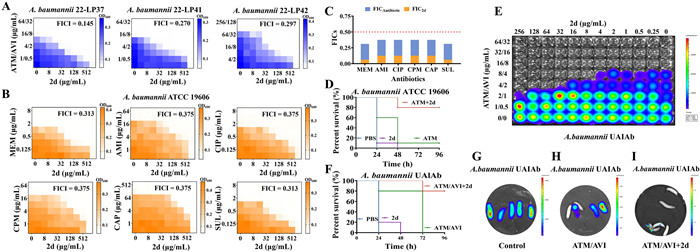
To further explore the synergistic effects of compound 2d in combination with other antibacterial agents, this study investigated the antimicrobial activity of 2d in combination with several representative antibacterial of different classes commonly used for clinical treatment of A. baumannii infections, including meropenem (MEM), amikacin (AMI), ciprofloxacin (CIP), cefepime (CPM) and chloramphenicol (CAP). Additionally, a β-lactamase inhibitor sulbactam (SUL), which has demonstrated clinical efficacy against A. baumannii, was also selected. As shown in Figs. 1B and C, compound 2d increased the susceptibility of A. baumannii to all evaluated antibacterial. The addition of 2d at concentrations of 64–128 µg/mL significantly lowered the inhibitory concentrations of MEM, AMI, CIP, CPM, CAP and SUL to 1/4 of their respective MICs (Table S6). The degree of combination responses was further analyzed and visualized by the online SynergyFinder software using zero interaction potency (ZIP) calculation method [27]. As depicted in Fig. S1 (Supporting information), the ZIP scores of 2d with various antibacterial agents, including ATM, MEM, AMI, CIP, CPM, CAP and SUL were 20.011, 15.605, 10.143, 13.891, 12.343, 8.496 and 12.602, respectively (ZIP score: >0 indicated synergy; >10 indicated strong synergy). These findings showed that compound 2d demonstrated highly synergistic effects when combined with the tested antibacterial against A. baumannii.
Galleria mellonella (G. mellonella) larvae have emerged as a reliable and commercially available model for studying the in vivo efficacy of antimicrobial agents against pathogen infections [28]. To investigate the protective effect of the 2d/ATM combination against A. baumannii infection, in vivo experiments were conducted using G. mellonella larvae. The larvae were challenged a lethal dose of 1 × 106 colony-forming units (CFU) of A. baumannii ATCC 19606, and treated with a single injection of 10 µL of phosphate buffer saline (PBS), 8.3 mg/kg 2d, 66.7 mg/kg ATM, or 2d and ATM (8.3 and 66.7 mg/kg) 0.5 h post-infection. As depicted in Fig. 1D, most larvae treated with PBS, 2d or ATM alone succumbed to the infection within 48 h of A. baumannii inoculation. Notably, the monotherapy of either compound failed to provide protection against the lethal infection. However, the combination of 2d and ATM showed a significant increase in the survival rate, with 80% of the larvae surviving up to 96 h post-infection. The survival rate was significantly higher in the combination group compared to the monotherapy groups (P < 0.01). Thus, the 2d and ATM combination exhibited potent protective efficacy against lethal A. baumannii infection.
To facilitate and visualize the evaluation of anti-A. baumannii adjuvant candidates, a stable autoluminescent ESBL-producing A. baumannii strain UAlAb was employed for potency study both in vitro and in vivo. As shown in Fig. 1E, the triple-combination of 2d (32 µg/mL) with ATM and AVI (4 and 2 µg/mL) exhibited significantly synergistic effect against A. baumannii UAlAb in checkerboard assay as demonstrated by bioluminescent monitoring. In the G. mellonella larvae model, the control group of infected larvae displayed the strongest luminescence at 24 h post-infection and eventually all died as displayed in Figs. 1F and G. The ATM/AVI group showed partial protection, with 3 out of 5 larvae developing varying degrees of luminescence at 24 h post-infection but ultimately all succumbing to the infection by 72 h (Figs. 1F and H). Meanwhile, the 2d/AVI/ATM triple-combination group showed minimal luminescence at 24 h post-infection (Fig. 1I), and had a significantly better protective efficacy, with 4 out of 5 larvae (80%) surviving at 96 h, compared to the ATM/AVI group (Figs. 1F and I). Therefore, the luminescence intensity of infected larvae at 24 h post-infection can serve as a valuable indicator of the protective efficacy.
To gain insight into the synergic mechanism of 2d, transcriptome analysis of A. baumannii ATCC 19606 exposure to ATM alone and in combination with 2d was performed. After co-cultured with ATM (4 µg/mL) for 2 h, the bacteria were treated with or without 2d at the concentration of 4 µg/mL for 4 h. The comparison of treatment with 2d/ATM combination to ATM alone revealed an up-regulation of 983 and down-regulation of 918 differentially expressed genes (Figs. 2A and B). Notably, the results revealed that the transcriptional levels of four efflux-pump coding genes, including adeA, adeB, emrA and emrB, were significantly up-regulated (Fig. 2C). This suggested that the possible suppression of the efflux-pump system by 2d led to a compensatory transcriptional up-regulation of the corresponding efflux-pump related genes. Next, the differential gene expression analysis was verified using quantitative PCR (RT-qPCR) assay. As depicted in Fig. 2D, the expression changes of adeB and adeA were more significant than those of emrA and emrB in A. baumannii ATCC 19606 between ATM/2d treatment group and ATM group. Antibiotic efflux is one of the major resistant mechanisms, whereby bacteria pump out the antibiotics from their cellular interior to the external environment using special transporter proteins called efflux pumps. It has gained significant attention in the discovery and optimization of synthetic and naturally derived efflux pump inhibitors (EPIs) in the past two decades. EPIs can provide an available option to restore the pharmacological activity and clinical efficacy of some first-line antibiotics.
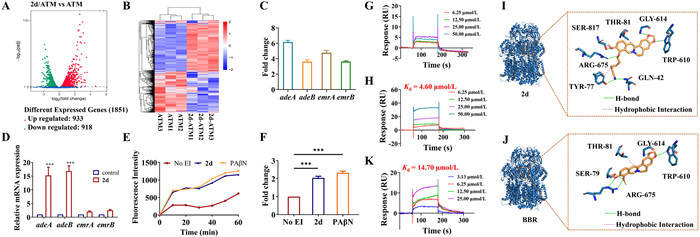
The capacity of 2d to impede the efflux of ethidium bromide, a recognized substrate of the efflux pump, from A. baumannii was subjected to further assessment utilizing a fluorescence-based assay. The fluorescence intensity of ethidium bromide experiences enhancement upon its interaction with bacterial DNA. As depicted in Figs. 2E and F, a notable increase in fluorescence was observed in the presence of 2d or PAβN, a widely recognized inhibitor of efflux pumps. This increase in fluorescence signifies a pronounced inhibition on the efflux of ethidium bromide, in comparison to the untreated control group. When A. baumannii was treated with 2d or PAβN, there emerged an elevation of approximately 2.04- and 2.32-fold in the fluorescence intensity of ethidium bromide, respectively, compared with the untreated control following a 60-min incubation. These findings collectively suggest that 2d likely hindered the efflux of ethidium bromide by rendering the bacterial efflux pumps inactive.
As one of the best studied efflux pumps, the resistance-nodulation-division (RND) family-type pump AdeABC was reported to be expressed in an estimated 80% of clinically isolated A. baumannii strains and has a broad substrate range. Similar to other RND-type pumps, AdeABC has a three-component structure: membrane fusion protein (AdeA), inner membrane efflux transporter (AdeB), and outer membrane protein (AdeC) [29]. Also, confocal analysis demonstrated that 2d could enter A. baumannii (Fig. S2 in Supporting information). Thus, according to the RT-qPCR results, recombinant AdeA and AdeB proteins were expressed and purified. The affinity with 2d was then evaluated through surface plasmon resonance (SPR) assay. As depicted in Figs. 2G and H, 2d could directly bind to immobilized AdeB fragment protein in a concentration-dependent manner, with a Kd value of 4.60 µmol/L, while 2d had no interaction with the AdeA protein, suggesting a possible interaction between 2d and AdeB.
The importance of the proximal multidrug binding site for AdeB activity has been established, as evidenced by the conservation of residues in the F and G loops among other multidrug pumps, such as AcrB and MtrD. To investigate the interaction between AdeB and 2d, molecular dockings were carried out, with BBR serving as a control. The G-loop residues of AdeB, which are 609GWGFSGA615, were chosen as the docking sites. As shown in Figs. 2I and J, compound 2d gave the affinity score of −8.3 kcal/mol, while BBR gave the score of −8.1 kcal/mol (affinity score < −7.0 kcal/mol was considered a strong interaction between protein and ligand; −7.0 kcal/mol < affinity score < −5.0 kcal/mol was considered good interaction; affinity score > −5.0 kcal/mol indicated a certain interaction) [30]. The molecular docking results indicate that both compounds fit well in the G-loop, forming three hydrogen bonds with the TRP610, GLY614 and ARG675. In addition, compound 2d also formed three more hydrogen bonds interactions with the GLN42, TYR77 and SER817, as well as hydrophobic interactions with THR81 and TYR77, which might contribute to its higher affinity. Molecular dynamics simulation analysis of 2d further demonstrated that the binding status was stable, and again verified the possible hydrogen bonds interactions between 2d and AdeB (Fig. S3 in Supporting information).
Additionally, the affinity between BBR and AdeB was also evaluated using SPR assay, and the Kd value was 14.73 µmol/L (Fig. 2K), while 2d gave the Kd value of 4.60 µmol/L (Fig. 2H). Our results verified that 2d had a stronger interaction affinity with AdeB, compared with that of BBR, indicating that 2d may be a potent inhibitor of the multidrug transporter AdeB. By preventing the efflux effect of structurally diverse antibiotics, 2d could potentially achieve a synergistic antimicrobial effect.
On account of the observed interaction between 2d and AdeB, the genotype of both the standard and clinically isolated strains utilized in the synergistic study was investigated by PCR. The strains demonstrated synergistic effects between 2d and ATM were found to harbor the adeB gene (Fig. 3A). In contrast, the other three strains (21-2, 21-5, 21-16) displayed no synergy in the checkerboard assay were lack of adeB gene. These results again substantiate the ability of compound 2d to augment the antibacterial efficacy of ATM against strains that produce AdeB.
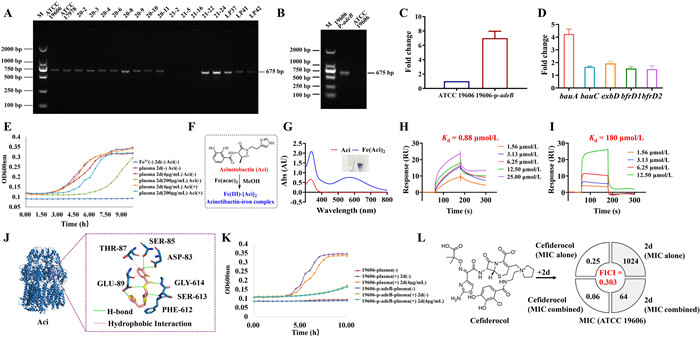
To further elucidate whether the action of 2d on drug efflux is specifically targeted at AdeB, an adeB-overexpressing strain, 19606-p-adeB, was constructed. Validation of adeB gene overexpression was ascertained by comparing the PCR amplification products of the extracted plasmids (Fig. 3B) and the gene transcription levels (Fig. 3C) of both the parental strain (ATCC 19606) and the mutant counterpart (19606-p-adeB). Given that the adeB gene in ATCC 19606 is solely located in the chromosome, no 675 bp band was amplified from the extracted plasmid of the wild-type strain. Conversely, amplified PCR products of adeB from the plasmid and a 7.00 ± 0.97-fold increase in mRNA expression level were observed in the overexpressing strain. Subsequently, a checkerboard assay was conducted utilizing the 19606-p-adeB strain, compared with the parental strain. As depicted in Table S7 (Supporting information), the MIC value of ATM increased from 32 µg/mL to 64 µg/mL in the 19606-p-adeB strain. Additionally, the presence of 2d at 4 µg/mL, which exhibited a significant synergistic effect with ATM in the wild-type strain, did not lead to a significant decrease in the MIC of ATM against the 19606-p-adeB strain, maintaining a value of 64 µg/mL. Therefore, no synergy was discernible between ATM and 2d in the 19606-p-adeB strain at this 2d concentration, as indicated by an FICI value of 1.003. When the concentration of 2d was increased to 32 µg/mL and above, the synergy between 2d and ATM was restored, with the MIC of ATM reduced to 16 µg/mL and an FICI value of 0.282. This observation implies that the overproduction of AdeB makes 2d unable to completely occupy all active sites of the AdeB protein, thus preventing the efflux of ATM.
In addition to the efflux pump system, transcriptome analysis showed significant up-regulation of genes related to iron acquisition (Fig. 3D), indicating that the administration of 2d may inhibit iron acquisition in A. baumannii. Specifically, the transcriptional level of acinetobactin (Aci) receptor BauA-coding gene bauA was significantly up-regulated, suggesting that the recycling of Aci in A. baumannii might be suppressed. Iron is a trace element that is essential for nearly all bacterial pathogens. However, the level of free iron at the host-pathogen interface is magnitude below what is required to support the microbial growth. To counteract iron starvation, bacteria have developed several iron acquisition strategies, including the secretion of iron-binding small molecules known as “siderophores” to steal iron from transferrin and lactoferrin. Aci is often identified as the major siderophore produced by A. baumannii, but it is still unclear how Aci is pumped out extracellularly.
As depicted in Fig. 3E, the iron deficiency state (indicated by the dark blue curve) of bacteria can be reversed through adding plasma (purple curve). However, the growth of bacteria in plasma can be then inhibited in a dose-dependent manner upon the addition of 2d (4 µg/mL in red and 200 µg/mL in green curve). In this case, the growth of bacteria could be restored upon the complementary addition of Aci (orange and cyan curve). These results indicate that after the administration of 2d, the amount of Aci excreted by bacteria is insufficient, leading to growth inhibition. The addition of exogenous Aci can reverse the growth inhibition of bacteria caused by 2d.
Based on a previous report, the expression of AdeB was increased more than four-fold under low iron conditions (20 µmol/L). We speculate that Aci is exported to the extracellular space through AdeABC pump for capturing Fe(Ⅲ) from the host. The complex was then taken back into the pathogens by the receptor protein (BauA). After the addition of 2d, it inactivated the AdeABC efflux pump system, and then suppressed the export of Aci and the subsequent iron uptake of A. baumannii. As shown in Figs. 3F and G, Aci could formed a stable and colored 2:1 complex with Fe(Ⅲ). SPR experiment also showed that Aci displayed a strong affinity binding with immobilized AdeB fragment protein with a Kd value of 0.88 µmol/L, while Fe(Ⅲ)-[Aci]2 gave a Kd value of 180 µmol/L, which further confirmed that AdeABC might be the efflux channel of Aci (Figs. 3H and I). The molecular docking analysis between Aci and AdeB protein was then performed. As shown in Fig. 3J, Aci could fit well in the active pocket of AdeB, where six hydrogen bonds and three hydrophobic interactions contributed together to their strong interactions, with an affinity score of −7.7 kcal/mol. It is worth noting that the addition of 2d showed no obvious inhibitory effect on the growth of 19606-p-adeB strain (green curve and cyan curve, Fig. 3K), indicating that the overexpression of AdeB might compensate for the restriction of iron acquisition and bacterial growth caused by 2d. Compared with the parental strain, the relatively slow growth rate of the 19606-p-adeB strain can be explained by the fitness cost effect after receiving the adeB overexpressing plasmid.
Cefiderocol is a novel siderophore cephalosporin conjugated with a catechol moiety, with a promising in vitro and in vivo activity against A. baumannii [31]. It was approved by the U.S. Food and Drug Administration (FDA) in 2019 to treat hospital-acquired pneumonia and ventilator-associated pneumonia caused by A. baumannii. Cefiderocol binds to Fe(Ⅲ) and is actively transported into bacteria through bacterial iron transporter and outer membrane. A study conducted with Hua et al. in 2023 showed that a combination of cefiderocol and CCCP, as an efflux pump inhibitor, has synergistic effect on infections caused by A. baumannii isolates [32]. We further verified the synergistic effect between 2d and cefiderocol against A. baumannii ATCC 19606 (Fig. 3L). The MIC of cefiderocol against A. baumannii was reduced from 0.25 µg/mL to 0.06 µg/mL with the addition of 64 µg/mL 2d. The synergic effect of 2d to cefiderocol might attribute to its suppressive effects on both iron uptake and efflux pump, leading to the increasing concentration of cefiderocol inside the bacteria. These data suggested that 2d could facilitate antibiotic efficacy by competing against antibiotics and Aci for the binding sites of AdeB, thus increasing the antibiotic uptake and the susceptibility against A. baumannii (Fig. 4).
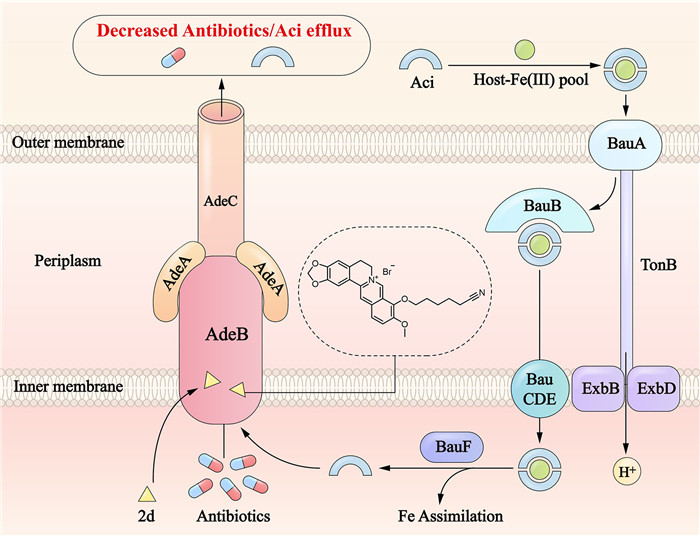
To summarize, this study designed, synthesized, and evaluated thirty novel BBR derivatives for their synergistic antibacterial activity against A. baumannii. Among these derivatives, compound 2d demonstrated the most promising synergetic potency, decreasing the MIC value of ATM to no more than 1/4 of its MIC when used alone, which was superior to that of BBR. Instead of killing the bacteria itself, compound 2d could exert its synergistic effect on the effectiveness of ATM by the reducing the efflux of ATM and enhancing ATM’s concentration in bacteria. Compound 2d also demonstrated in vitro synergistic effects against CRAB strains and ESBLs producing strains in dual- and triple- combinations, respectively. In G. mellonella larvae model, 2d/ATM combination could significantly increase the survival rate of the larvae compared to either component alone. Mechanism study disclosed that 2d might inhibit the efflux pump and iron acquisition of A. baumannii by binding to AdeB, thereby achieving synergistic antimicrobial efficacy (Fig. 4). Our study provides instructive guidance for the further development of BBR derivatives as anti-A. baumannii antibiotic adjuvants with a dual-function antibacterial mechanism.
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
We appreciate Dr. Tianyu Zhang (Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences) for kindly providing the autoluminescent Acinetobacter baumannii strain UAlAb. This research project was supported by grants from National Natural Science Foundation of China (Nos. 32141003, 82104013), and CAMS Initiative for Innovative Medicine (Nos. 2021-1-I2M-070, 2021-1-I2M-039, China).
Supplementary material associated with this article can be found, in the online version, at doi:
E.S. Milani, A. Hasani, M. Varschochi, et al., J. Hosp. Infect. 117 (2021) 135–146. doi: 10.1016/j.jhin.2021.09.010
T. Demirdal, U.S. Sari, S.A. Nemli, Ann. Clin. Microbiol. Antimicrob. 15 (2016) 11–17. doi: 10.1186/s12941-016-0123-7
O. Ayobami, N. Willrich, T. Harder, et al., Emerg. Microbes Infect. 8 (2019) 1747–1759. doi: 10.1080/22221751.2019.1698273
S. Domingues, N. Rosário, Â. Cândido, et al., Microorganisms 7 (2019) 30–37. doi: 10.3390/microorganisms7020030
W.P. Xu, Z. Ma, G. Dhanda, et al., Chin. Chem. Lett. 34 (2023) 107847. doi: 10.1016/j.cclet.2022.107847
C. Willyard, Nature 543 (2017) 15. doi: 10.1038/nature.2017.21550
G.D. Wright, Trends Microbiol. 24 (2016) 928. doi: 10.1016/j.tim.2016.07.008
E.E. Gill, O.L. Franco, E.W. Hancock Robert, Chem. Biol. Drug Des. 85 (2014) 56–78.
L. Kalan, G.D. Wright, Expert. Rev. Mol. Med. 13 (2011) e5. doi: 10.1017/S1462399410001766
Y. Chen, Z. Li, Y. Yin, et al., Chin. Chem. Lett. 34 (2023) 107948. doi: 10.1016/j.cclet.2022.107948
C.F. Liu, Z.X. Li, H. Yu, et al., Chin. Chem. Lett. 32 (2021) 1385–1389. doi: 10.1016/j.cclet.2020.10.048
Q.X. Zeng, W. Wei, T. Fan, et al., CCS Chem. 4 (2022) 1535. doi: 10.31635/ccschem.021.202100986
W. Wei, Q.X. Zeng, Y. Wang, et al., Acta Pharm. Sin. B 13 (2023) 2138–2151. doi: 10.1016/j.apsb.2022.12.009
M. Tillhon, L.M. Guamán Ortiz, P. Lombardi, et al., Biochem. Pharmacol. 84 (2012) 1260–1267. doi: 10.1016/j.bcp.2012.07.018
X. Li, Y. Song, L. Wang, et al., Front. Cell Infect. Microbiol. 11 (2021) 660431. doi: 10.3389/fcimb.2021.660431
H. Yao, L. Cui, H. Liu, et al., Chin. Chem. Lett. 35 (2024) 108511. doi: 10.1016/j.cclet.2023.108511
G. Chen, L.R. Swem, D.L. Swem, et al., Mol. Cell 42 (2011) 199–209. doi: 10.1016/j.molcel.2011.04.003
T.L. Harris, R.J. Worthington, L.E. Hittle, et al., ACS Chem. Biol. 9 (2014) 122–127. doi: 10.1021/cb400490k
J. Sianturi, P. Priegue, J. Hu, et al., Angew. Chem. Int. Ed. 61 (2022) e202209556. doi: 10.1002/anie.202209556
S. Siricilla, K. Mitachi, J. Yang, et al., J. Med. Chem. 60 (2017) 2869–2878. doi: 10.1021/acs.jmedchem.6b01805
A. Hammad, N.S. Abutaleb, M.M. Elsebaei, et al., J. Med. Chem. 62 (2019) 7998–8010. doi: 10.1021/acs.jmedchem.9b00720
H. Sun, M.F. Ansari, N. Battini, et al., Org. Chem. Front. 6 (2019) 319–334. doi: 10.1039/c8qo01180j
T.Y. Fan, Y.X. Wang, S. Tang, et al., Eur. J. Med. Chem. 157 (2018) 877–886. doi: 10.1016/j.ejmech.2018.08.050
G.B. Zhang, S.K. Maddili, V.K.R. Tangadanchu, et al., Sci. China Chem. 61 (2017) 557–568.
W.W. Gao, L. Gopala, R.R.Y. Bheemanaboina, et al., Eur. J. Med. Chem. 146 (2018) 15–37. doi: 10.4103/wjtcm.wjtcm_4_18
L. Di, E.H. Kerns, Hong Y, et al., Int. J. Pharm. 297 (2005) 110–119. doi: 10.1016/j.ijpharm.2005.03.022
A. Ianevski, A.K. Giri, T. Aittokallio, Nucl. Acids Res. 48 (2020) W488–W493. doi: 10.1093/nar/gkaa216
Y. Tao, L. Duma, Y. Rossez, Pathogens 10 (2021) 1483. doi: 10.3390/pathogens10111483
M. Zwama, K. Nishino, Antibiotics 10 (2021) 774. doi: 10.3390/antibiotics10070774
K.Y. Hsin, S. Ghosh, H. Kitano, PLoS One 8 (2013) e83922. doi: 10.1371/journal.pone.0083922
M. Kollef, H. Dupont, D.E. Greenberg, et al., Int. J. Antimicrob. Agents 62 (2023) 106882. doi: 10.1016/j.ijantimicag.2023.106882
X.C. Liu, Y.J. Chang, Q.Y. Xu, et al., mSystems 8 (2023) e0129122.

Scheme 1 Chemical structures of antibiotic adjuvants against A. baumannii, and structure modification strategy on BBR.

Figure 1 Synergistic antibacterial effects in vitro and in vivo for 2d against A. baumannii. (A) Heat map for the antibacterial activity of 2d in combination with ATM/AVI in ESBL-producing A. baumannii 22-LP37, 22-LP41 and 22-LP42 in checkerboard assay. (B) Heat map for the antibacterial activity of 2d in combination with MEM, AMI, CIP, CPM, CAP and SUL against A. baumannii ATCC 19606 in checkerboard assay. (C) Bar chart representing the FICI values of 2d in combination with MEM, AMI, CIP, CPM, CAP and SUL against A. baumannii ATCC 19606. The red dash line delineates the cut-off of synergy with an FICI of 0.5. (D) Survival curves of G. mellonella larvae following the infection of A. baumannii ATCC 19606 and the treatment of PBS, 2d, ATM or 2d/ATM combination (n = 10). (E) Checkerboard assay of 2d with ATM/AVI against autoluminescent A. baumannii strain UAlAb. (F) Survival curves of G. mellonella larvae following the infection of A. baumannii strain UAlAb and the treatment of PBS, 2d, ATM or 2d/ATM combination (n = 5). Luminescence images of G. mellonella larvae (n = 5) collected 24 h following the infection of autoluminescent A. baumannii strain UAlAb and the treatment of (G) PBS, (H) ATM/AVI or (I) 2d/ATM/AVI combination.

Figure 2 Target proteins analysis and verification. (A) Volcano plot and (B) Heatmap analysis of the differential expression genes in A. baumannii after exposing the combination of ATM (4 µg/mL) plus 2d (4 µg/mL) or ATM (4 µg/mL). (C) The up-regulation of efflux-pump related genes of 2d and ATM (4 and 4 µg/mL)-treated A. baumannii in comparison to ATM (4 µg/mL) in transcriptome analysis. (D) Quantification of efflux pump genes expression normalized with control (ATM 4 µg/mL). ATM and 2d were added at 4 and 4 µg/mL for 4 h. (E) The relative fluorescence intensity of ethidium bromide in A. baumannii ATCC 19606 with the treatment of 2d and PAβN during 60 min incubation. (F) The fold change of relative fluorescence intensity of ethidium bromide in A. baumannii ATCC 19606 with treatment of 2d and PAβN strain at 60 min post incubation. (G) SPR sensorgrams obtained on a purified AdeA-coated chip at different concentrations of 2d. (H) SPR sensorgrams obtained on an AdeB fragment-coated chip at different concentrations of 2d. (I) Molecular docking for AdeB (PDB code: 6OWS) with 2d. (J) Molecular docking for AdeB (PDB code: 6OWS) with BBR. (K) SPR sensorgrams obtained on an AdeB fragment-coated chip at different concentrations of BBR. Values are mean ± standard deviation (SD) (n = 3). ***P < 0.001.

Figure 3 The influence on iron uptake by the interaction of compound 2d and AdeB protein. (A) Electrophoresis of PCR products of standard and clinical isolated A. baumannii strains, designing to verify the existence of adeB gene. Lane 1, molecular weight marker. Lanes 2 and 19, PCR products obtained from standard strains and clinical isolates. (B) Electrophoresis of PCR products of the extracted plasmids from mutant strain 19606-p-adeB and A. baumannii ATCC 19606 wild type. (C) Quantification of adeB gene expression levels. (D) Changes in transcriptional levels of iron uptake related genes of 2d and ATM (4 and 4 µg/mL)-treated A. baumannii in comparison to the ATM (4 µg/mL) group. (E) Growth curve of A. baumannii ATCC 19606 in M9 iron deficient media as measured at OD600. (F) Structure of acinetobactin (Aci) and synthesis of Fe(Ⅲ)-[Aci]2. (G) Ultraviolet–visible spectrophotometry (UV–vis) absorption spectra of Aci (200 µmol/L) and Fe(Ⅲ)-[Aci]2 (200 µmol/L) in HEPES (1 mol/L, pH 7.4). SPR sensorgrams obtained on a purified AdeB fragment-coated chip at different concentrations of (H) Aci or (I) Fe(Ⅲ)-[Aci]2. (J) Molecular docking for AdeB (PDB code: 6OWS) with Aci. (K) Growth curve of AdeB overexpressing and wild-type A. baumannii 19606 strains in M9 iron deficient media as measured at OD600. (L) Structure of cefiderocol and results of checkerboard assay for 2d/cefiderocol combination against A. baumannii ATCC 19606. Values are mean ± SD (n = 3).

 扫一扫看文章
扫一扫看文章

扫一扫关注我们
 DownLoad:
DownLoad:
 下载:
下载:

 下载:
下载: