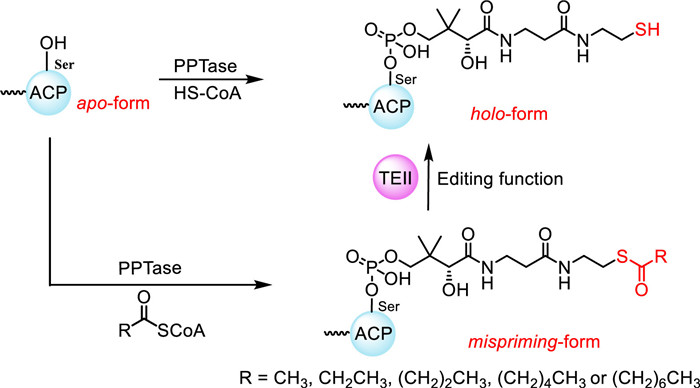
Scheme 1.
Editing TEIIs hydrolyze the acyl residues tethered to ACP, converting the ACP from mispriming-form to holo-form.

Most polyketides in fungi are synthesized by iterative polyketide synthases (PKSs), which have one copy of catalytic and carrier protein domain that used iteratively in each catalytic cycle of chain elongation [1-3]. Notably, before attaching extend units, the acyl carrier protein (ACP) domain of PKS requires posttranslational modification by phosphopantetheinyl transferase (PPTase), which is a type of enzyme that introduces a 4′-phosphopantetheine (4′-PP) arm derived from coenzyme A (CoA) to the serine side chain within the ACP domain, priming the inactive apo-form ACP to the active holo-form ACP (Scheme 1) [2,4]. The biosynthetic intermediates are covalently tethered to the 4′-PP arm of ACP domain through a thioester linkage and catalyzed by ketosynthase (KS), ketoreductase (KR), dehydratase (DH), and/or other catalytic domains of PKS, until the mature polyketide chains are ultimately released [2,5]. Thioesterases (TE), including both type Ⅰ thioesterases (TEIs) integrated into PKS assembly lines and trans-standalone type Ⅱ thioesterases (TEIIs), play a crucial role in releasing polyketide chains. These TEs mainly function to release the polyketide chains through hydrolysis or macrocyclization [6,7]. For instance, LovG catalyzes the release of dihydromonacolin L acid via hydrolysis, leading to a chain product during the biosynthesis of lovastatin [8]. While BerkB catalyzes the macrolactonization of a C16 polyketide chain, resulting in a cyclic product berklactone A [9]. Additionally, reductase (R) domains [10] and acyltransferase-like enzymes [11] have also been reported to release final products of fungal PKSs.
A particular class of TEIIs encoded in fungal gene clusters containing nonreducing PKS with reductase domain (nrPKS-R) is of particular interest. Knocking out these TEIIs in their hosts decreased but did not completely abolish the production of final polyketide products. Examples of such TEIIs include CitA [10], MrpigG [12], AfoC [13], and DbaE [14], which are encoded in the biosynthetic gene clusters of citrinin, azaphilones, asperfuranone, and 2,4-dihydroxy-3-methyl-6-(2-oxopropyl)benzaldehyde, respectively. Since the R domains in nrPKSs, such as CitS and MrpigA, are certainly responsible for chain release [10,12,15], these TEIIs seem to be unnecessary for the biosynthesis of these products and are instead considered as post-modification enzymes [13,16].
Here, we demonstrate that fungal TEIIs in nrPKS-R gene clusters possess editing function, instead of product releasing. TEIIs with the editing function rectify the mispriming-form ACP to the holo-form to ensure the correct extension of the polyketide chains (Scheme 1). Besides, TEIIs encoded in gene clusters containing highly-reducing PKS with condensation domain (hrPKS-C) were also found to display editing function. Therefore, editing TEIIs are applied to improve the yield of polyketide products, which provide a valuable strategy for metabolic engineering.
It has been reported that occasionally decarboxylation of extender units by KS domain occurs before condensation, resulting acetyl and propionyl residues covalently attach to the ACP [17]. In addition, bacterial PPTase was reported to recognize acylated CoAs and attach them to ACP, leading to the function blocking of the PKS [18]. Indeed, the PPTase is often the case, since lots of CoAs in the cell are in the acylated form [19]. To explore the substrate promiscuity of fungal PPTase, we expressed and purified the fungal PPTase, NpgA and the ACP domain of nrPKS, CitS (CitS-ACP) from Escherichia coli (Fig. S1 in Supporting information). The enzymatic activity of NpgA was tested by using CitS-ACP as 4′-PP arm acceptor and various CoAs (CoA, acetyl-CoA, propionyl-CoA, butyryl-CoA, hexanoyl-CoA, and octanoyl-CoA, respectively) as 4′-PP arm donors. Ultraperformance liquid chromatography coupled with a time-of-flight mass spectrometer (UPLC-TOF-MS) analyses of the tested samples from in vitro assays suggested that NpgA could convert apo-form ACP not only to holo-form, but also to acyl-mispriming-form in the presence of acyl-CoAs (Scheme 1 and Figs. S3–S9 in Supporting information), showing the substrate promiscuity of fungal PPTase. Once the thiol group of 4′-PP arm is occupied by acyl residues, the extend units could not be attached to ACP, thus blocking the function of PKS to form exact polyketide scaffolds. Therefore, the mispriming-ACPs must be re-activated before the catalytic function of PKS can be performed.
Editing TEII could be a potential strategy to recover PKS in fungi, and CitA in citrinin biosynthetic gene cluster was a typical example for further investigation. Hence, the full-length of citA was amplified from the cDNA of Monascus ruber [8], and CitA was successfully expressed and purified from E. coli. Incubation of CitA with acyl-mispriming-CitS-ACPs (acetyl-, propionyl-, butyryl-, hexanoyl-, and octanoyl-mispriming-CitS-ACP, respectively) was carried out and UPLC-TOF-MS was used to detect the mass of those mispriming-CitS-ACPs after incubation with CitA. The results showed that loss of mass from acyl hydrolysis was observed for all acyl-mispriming-ACPs (Scheme 1 and Figs. S10–S14 in Supporting information). Therefore, CitA was identified as an editing TEII converting acyl-mispriming-ACP to holo-ACP, recovering the function of PKS which was blocked by substrate promiscuous PPTase.
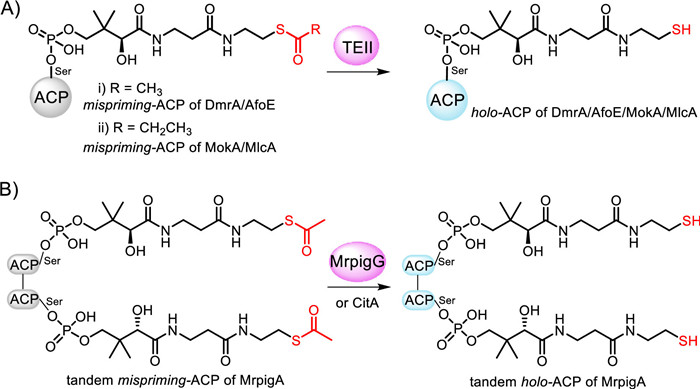
To explore the editing TEIIs in fungi systematically, we conducted the phylogenetic analysis of CitA and other TEIIs in fungi (Fig. 1A). The results showed that the TEIIs encoded in nrPKS gene clusters are clustered into a separate clade from those in hrPKS gene clusters. The multiple sequence alignments showed that the protein sequences of TEIIs in nrPKS gene clusters possess high similarity, while TEIIs in hrPKS gene clusters are not (Fig. S15 in Supporting information). The nrPKS-R gene clusters, dmr, mrpig, and afo responsible for the biosynthesis of compound 2 (Fig. 1C), azaphilone pigment and asperfuranone, respectively, were selected for further exploration of TEII function [12,13]. We heterologously expressed and purified these three TEIIs, DmrB, AfoC, and MrpigG and their corresponding ACP, DmrA-ACP, AfoE-ACP, and MrpigA-ACP from E. coli separately. In vitro assays were conducted and the resulted samples were analyzed by UPLC-TOF-MS. The results showed that these ACP domains could be mis-primed by PPTase, and these three TEIIs can re-activate ACPs by hydrolyzing acetyl group from acetyl-mispriming-DmrA-ACP, acetyl-mispriming-AfoE-ACP, and tandem-acetyl-mispriming-MrpigA-ACP, respectively (Fig. 2 and Figs. S16–S30 in Supporting information). Particularly, two ACP domains, ACP1 and ACP2 in MrpigA were both activated by PPTase, and the acetyl groups of them were hydrolyzed by TEII MrpigG simultaneously.
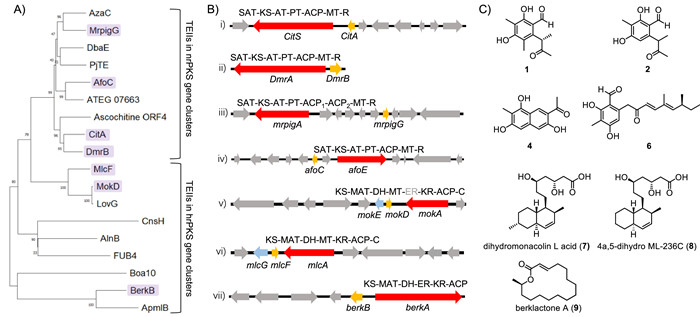
Although TEIIs in hrPKS gene clusters are normally considered to release polyketide chains [8,9], it is still worth to evaluate their potential editing function. So, we choose three typical hrPKS gene clusters, mok, mlc, and berk responsible for the biosynthesis of monacolin K (lovastatin) [20], compactin [21] and A26771B [9], respectively, for further investigation. Phylogenetic analysis showed that the TEIIs, MokD and MlcF, are close to the clade of CitA homologous, while BerkB is not (Fig. 1A). Interestingly, the C-terminal of the PKSs, MokA and MlcA, possess C domain, which is not occur in BerkA (Fig. 1B). We conducted in vitro assays using purified hrPKS-ACPs, NpgA and TEIIs. The results showed that MokD and MlcF have editing function, as evidenced in the assays for catalyzing propionyl-mispriming-MokA-ACP and propionyl-mispriming-MlcA-ACP, respectively (Fig. 2A and Figs. S31–S39 in Supporting information). However, no editing function was found for BerkB to hydrolyze propionyl-mispriming-BerkA-ACP (Figs. S40–S43 in Supporting information). Therefore, TEIIs such as MokD and MlcF in fungal hrPKS gene clusters could be bifunctional enzymes which are responsible for not only releasing product but also hydrolyzing mispriming-ACP to holo-form. Meanwhile, we realized that the editing TEIIs encoded in hrPKS gene clusters is not universal, since BerkB does not display hydrolytic activity in vitro. We speculate that other endogenous enzymes could hydrolyze the terminal acyl of mispriming-BerkA-ACP, or the PKS, BerkA, is hard to be misprimed by PPTase in vivo. In addition, we predicted the structures of typical ACPs and TEIIs in this research using AlphaFold2 and docked ACPs and TEIIs (CitS-ACP and CitA, MokA-ACP and MokD, BerkA-ACP and BerkB) using HDOCK
After identifying the editing function of TEIIs in vitro, we further confirmed their function in vivo. Heterologously expressing the gene citS encoding PKS alone in Aspergillus nidulans led to the production of trace 1 [10], while co-expressing citA and citS increased the yield of 1 (Fig. 3A). We then heterologously expressed dmrA in A. nidulans, which led to the production of 2 whose structure was identified based on 1D and 2D NMR data (Figs. S49–S52 in Supporting information). Co-expressing dmrA and dmrB (encoding TE) significantly increased the yield of 2 (Fig. 3B). In addition, heterologous expression of mrpigA in A. nidulans led to the production of 3, which could be produced via spontaneous transformation of 4 or catalyzed by other endogenous enzymes (Fig. 3C) [12]. Co-expression of mrpigG and mrpigA in A. nidulans led to the higher production of 3 than that expressing mrpigA alone (Fig. 3D). Taken together, our results indicated that expression of editing TEIIs could improve the yield of PKS products in vivo.
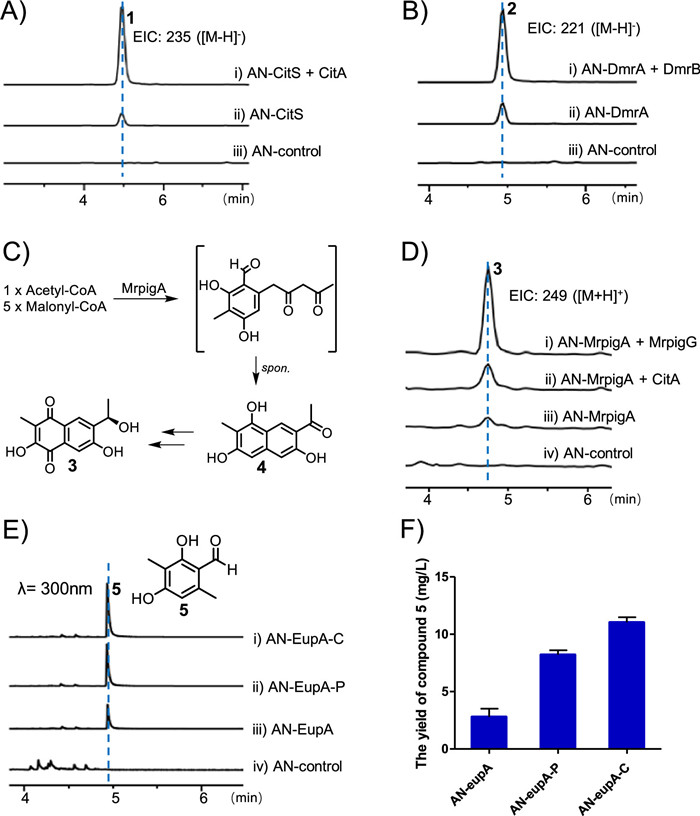
Considering the improved yield of polyketide products by expressing editing TEIIs in vivo and the catalytic promiscuity of CitA in hydrolyzing the acyl-mispriming-ACPs in vitro, we supposed that CitA and its homologs could serve as metabolic engineering tool to improve the yield of polyketides from other gene clusters. Therefore, we heterologously co-expressed citA with different genes encoding PKS in A. nidulans to verify the application of editing TEIIs. We firstly transformed citA to A. nidulans expressing mrpigA which produced 3, and detected the increased yield of 3, suggesting the potential of CitA as metabolic engineering tool (Fig. 3D). Then, we further verified its application in vivo for the product of EupA (a R domain contained nrPKS) in eup gene cluster which contains no TEII from Penicillium janthinellum [22]. When we expressed eupA alone in A. nidulans, the production of 5 was detected; while co-expressed eupA and citA, the yield of 5 was increased by 3.9-fold higher than that expressing eupA alone (Figs. 3E and F). Although there is no TEII in the eup gene cluster, we detected a CitA homolog PjTE (Fig. 1A and Fig. S15A) in the genome of P. janthinellum and supposed that PjTE could also be an editing TEII. As expected, the yield of 5 was significantly improved when co-expressing pjTE and eupA in A. nidulans, which is 2.9-fold higher than that expressing eupA alone (Figs. 3E and F). In summary, expressing editing TEII with PKSs in vivo could improve the yield of various polyketides, which suggested that editing TEII is a potential metabolic engineering tool to facilitate the production of polyketides with high efficiency.
Our study was motivated by the presence of highly conserved TEIIs in fungi nrPKS-R gene clusters. By successfully elucidating their editing function, we were able to address the issue that why deleting TEIIs in the hosts decreased the yield of polyketides [10,13,16,19]. Unlike transcription factors that regulate the yield of secondary metabolites at the transcriptional level, these TEIIs improve the yield of polyketides through their editing function. In prokaryotes, TEIs and TEIIs act synergistically to release product and remove incorrect intermediates, respectively [17,19]. Similarly, in the fungal nrPKS-R and TEII system, the R domain is responsible for releasing correct product, while the TEII is responsible for removing incorrect intermediates. Although fungal TEI in nrPKS gene cluster has been reported to display editing function during aflatoxin biosynthesis [23], our study demonstrated that TEIIs in nrPKS gene clusters function to hydrolyze acyl group tethered to ACP by PPTase, correcting the posttranslational modification process of PKS itself. Based on the elaboration of the editing function of CitA and its homologous, the role of this class of TEIIs in fungal polyketide biosynthesis is fully clarified.
During the in vitro assay for TEIIs in fungal hrPKS gene clusters, MokD and MlcF were found to possess both releasing and editing functions, which hydrolyzed the correct polyketide chains and the nonreactive acyl groups. There is a possibility that these TEIIs have a wide substrate spectrum, the hydrolysis of thioester substrate in vivo depends on how long they stay on the ACP domain. The correct intermediates will fast shuttle on the PKS assembly line and are difficult to be caught by TEIIs.
The different effect of heterologous expression of TEIIs, mrpigG and citA, with mrpigA showed that the editing TEIIs may exhibit cluster-specific manner (Fig. 3D). This result may rely on the interaction between the TEII and the PKS. Since the protein sequences of CitA, MrpigG, and their homologous TEIIs possess high similarity (Fig. S15A), a specific recognition region in TEIIs may critical for the interaction with PKS. It was reported that CitA show weak activity for the thioester bond of acyl-CoA, which is not attached to ACP, suggested that the ACP is essential for the function of TEIIs [18]. Indeed, the interaction of different domains, such as SAT and TE domains, of nrPKS has been reported [24]. Therefore, further research devoted to study the interactions between TEII and PKS is necessary, which could facilitate the utilization of editing TEIIs to precisely control the yield of polyketide products.
To summarize, we have clarified the editing function of TEIIs in the biosynthesis of fungal polyketides, demonstrated that TEIIs in nrPKS-R and hrPKS-C gene clusters are responsible for hydrolyzing acyl groups that can stall the PKS assembly line. Additionally, editing TEIIs are applied as metabolic engineering tool to improve the yield of polyketide products. This finding is unprecedented in fungal polyketide biosynthesis and opens a door for future tuning of the yield of fungal polyketides by editing TEIIs.
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
This work was supported financially by the National Natural Science Foundation of China (Nos. 82225042, T2192973) and the CAMS Innovation Fund for Medical Sciences (No. CIFMS 2021-I2M-1-029).
Supplementary material associated with this article can be found, in the online version, at doi:
J. Staunton, K.J. Weissman, Nat. Prod. Rep. 18 (2001) 380–416. doi: 10.1039/a909079g
J. Wang, Z. Deng, J. Liang, Z. Wang, Nat. Prod. Rep. 40 (2023) 1498–1520. doi: 10.1039/d3np00015j
H. Zhang, Y. Huang, Y. Tang, D. Kong, Y. Zou, Chin. Chem. Lett. 35 (2024) 108968. doi: 10.1016/j.cclet.2023.108968
E. Skellam, Nat. Prod. Rep. 39 (2022) 754–783. doi: 10.1039/d1np00056j
A.T. Keatinge-Clay, Nat. Prod. Rep. 29 (2012) 1050–1073. doi: 10.1039/c2np20019h
R.F. Little, C. Hertweck, Nat. Prod. Rep. 39 (2022) 163–205. doi: 10.1039/d1np00035g
L. Du, L. Lou, Nat. Prod. Rep. 27 (2010) 255–278. doi: 10.1039/B912037H
W. Xu, Y.H. Chooi, J.W. Choi, et al., Angew. Chem. Int. Ed. 52 (2013) 6472–6475. doi: 10.1002/anie.201302406
Y. Zhang, J. Bai, L. Zhang, C. Zhang, B. Liu, Y. Hu, Angew. Chem. Int. Ed. 60 (2021) 6639–6645. doi: 10.1002/anie.202015442
Y. He, R.J. Cox, Chem. Sci. 7 (2016) 2119–2127. doi: 10.1039/C5SC04027B
X. Xie, M.J. Meehan, W. Xu, P.C. Dorrestein, Y. Tang, J. Am. Chem. Soc. 131 (2009) 8388–8389. doi: 10.1021/ja903203g
W. Chen, R. Chen, Q. Liu, et al., Chem. Sci. 8 (2017) 4917–4925. doi: 10.1039/C7SC00475C
Y.M. Chiang, E. Szewczyk, A.D. Davidson, et al., J. Am. Chem. Soc. 131 (2009) 2965–2970. doi: 10.1021/ja8088185
J. Gerke, O. Bayram, K. Feussner, et al., Appl. Environ. Microbiol. 78 (2012) 8234–8244. doi: 10.1128/AEM.01808-12
M.W. Mullowney, R.A. McClure, M.T. Robey, N.L. Kelleher, R.J. Thomson, Nat. Prod. Rep. 35 (2018) 847–878. doi: 10.1039/c8np00013a
A.O. Zabala, W. Xu, Y.H. Chooi, Y. Tang, Chem. Biol. 19 (2012) 1049–1059. doi: 10.1016/j.chembiol.2012.07.004
M.L. Heathcote, J. Staunton, P.F. Leadlay, Chem. Biol. 8 (2001) 207–220. doi: 10.1016/S1074-5521(01)00002-3
P.A. Storm, C.A. Townsend, Chem. Commun. 54 (2017) 50–53.
M. Kotowska, K. Pawlik, Appl. Microbiol. Biotechnol. 98 (2014) 7735–7746. doi: 10.1007/s00253-014-5952-8
Y. Zhang, Z. Chen, Q. Wen, et al., Food Funct. 11 (2020) 5738–5748. doi: 10.1039/d0fo00691b
Y. Abe, T. Suzuki, C. Ono, et al., Mol. Genet. Genomics 267 (2002) 636–646. doi: 10.1007/s00438-002-0697-y
J. Liu, J. Lu, C. Zhang, et al., Nat. Chem. 15 (2023) 1083–1090. doi: 10.1038/s41557-023-01260-8
A.L. Vagstad, S.B. Bumpus, K. Belecki, N.L. Kelleher, C.A. Townsend, J. Am. Chem. Soc. 134 (2012) 6865–6877. doi: 10.1021/ja3016389
Q. Liu, D. Zhang, S. Gao, et al., Angew. Chem. Int. Ed. 62 (2023) e202214379. doi: 10.1002/anie.202214379

Scheme 1 Editing TEIIs hydrolyze the acyl residues tethered to ACP, converting the ACP from mispriming-form to holo-form.

Figure 1 Identification of TEIIs in fungal polyketide biosynthetic gene clusters. (A) Phylogenetic analysis of TEIIs in nrPKS and hrPKS gene clusters. The TEIIs identified in this study are shown in purple background. (B) Biosynthetic gene clusters of nrPKS (i–iv) and hrPKS (v–vii) with TEIIs in this study. The red arrows represent PKSs, the orange arrows represent TEIIs, the blue arrows represent trans-enoylreductases. (C) Products of PKSs. Compounds 1, 2, 4, 6, 7, 8, and 9 are the products of CitS, DmrA, MrpigA, AfoE, MokAE, MlcAG, and BerkA, respectively. SAT: Starter-unit acyltransferase; KS: Ketosynthase; AT: Acyltransferase; PT: Product template; MT: Methyltransferase; R: Reductase; DH: Dehydratase; ER: Enoylreductase; KR: Ketoreductase; MAT: Malonyl-acetyl transferase; C: Predicted condensation domain.

Figure 2 The editing function of TEIIs in in vitro assays. (A) The in vitro editing functions of DmrB, AfoC, MokD, and MlcF for mispriming-DmrA-ACP, mispriming-AfoE-ACP, mispriming-MokA-ACP and mispriming-MlcA-ACP, respectively. (B) The in vitro editing functions of MrpigG for the tandem-mispriming-ACP of MrpigA.

Figure 3 Verifying the function of editing TEIIs in vivo. (A) The production of 1 in A. nidulans co-expressing citS and citA. (B) The production of 2 in A. nidulans co-expressing dmrA and dmrB. (C) The proposed biosynthetic pathway of 3. (D) The production of 3 in A. nidulans co-expressing mrpigA and various TEs (mrpigG or citA). (E) The production of 5 in A. nidulans expressing eupA and editing TEIIs (citA or pjTE). (F) Comparison of the yield of 5 in A. nidulans expressing eupA and editing TEIIs. Values are presented as the mean yield from three biological replicates.

 扫一扫看文章
扫一扫看文章

扫一扫关注我们
 DownLoad:
DownLoad:
 下载:
下载:
 下载:
下载: