Citation:
Yan Liu, Yang Wang, Jiayi Zhu, Xuxian Su, Xudong Lin, Liang Xu, Xiwen Xing. Employing pH-responsive RNA triplex to control CRISPR/Cas9-mediated gene manipulation in mammalian cells[J]. Chinese Chemical Letters,
2024, 35(9): 109427.
doi:
10.1016/j.cclet.2023.109427
Employing pH-responsive RNA triplex to control CRISPR/Cas9-mediated gene manipulation in mammalian cells
English
Employing pH-responsive RNA triplex to control CRISPR/Cas9-mediated gene manipulation in mammalian cells
Guangdong Provincial Key Laboratory of Sensor Technology and Biomedical Instrument, School of Biomedical Engineering, Shenzhen Campus of Sun Yat-sen University, Sun Yat-sen University, Shenzhen 518107, China
b.
MOE Key Laboratory of Bioinorganic and Synthetic Chemistry, School of Chemistry, Sun Yat-sen University, Guangzhou 510275, China
c.
Department of Cardiology, the first people's hospital of Foshan, Foshan 528000, China
d.
Department of Biotechnology, College of Life Science and Technology, Jinan University, Guangzhou 510632, China
xingxiwen@jnu.edu.cn (X. Xing). 1 These authors contributed equally to this work.
Received Date:
21 September 2023 Accepted Date:
12 December 2023 Revised Date:
10 December 2023 Available Online:
15 September 2024
Abstract:
The clustered regularly interspersed short palindromic repeats/CRISPR-associated protein 9 (CRISPR/Cas9) system is an RNA-guided platform for highly efficient and specific genome targeting in diverse organisms, which has been exploited for various applications in gene manipulation. Compared with the constantly active CRISPR/Cas9 function, conditional control of its activity can improve the performance of the system with reduced side effects and high spatiotemporal precision. The pH-responsive triplex RNA was successful used in CRISPR-derived RNA/trans-activating crRNA (crRNA/tracrRNA) of CRISPR/Cas9, thus affecting RNA/dead Cas9 (dCas9) complex to target DNA in vitro and in vivo. This design of triplex RNA opens a new window towards the broad involvement of eukaryotic cells for conditional control of CRISPR/Cas9 function.
Clustered regularly interspersed short palindromic repeats/CRISPR-associated protein 9 (CRISPR/Cas9) technology has attracted extensive attention in biochemical and therapeutic fields due to its highly versatile applications in gene editing [1], transcriptional regulation [2], nucleobase modification [3], and genomic imaging [4]. Due to the typical constitutive activity of CRISPR/Cas9, flexible and controllable manipulation through specific stimuli is particular important for reducing adverse side effects and achieving temporal-spatial control [5–9]. At present, Cas9 protein or guide RNA (gRNA) can be engineered, but the modified Cas9 protein tends to be global rather than selective to specific genes [10–13], so the manipulation of specific genes is often achieved by engineering the guide RNA [14,15].
In the cellular environment, physiological pH conditions are maintained through tight homeostatic control, and intracellular pH changes plays a critical role in numerous important biological processes, such as cancer development [16–18]. Thus, it is a great value to develop a programmable engineering molecular system which can respond to pH changes in the cellular environment. Some nucleic acid structures are reported to be able to response to the intracellular pH changes, such as DNA i-motif [19,20] and DNA/RNA triplex [21]. However, these nucleic acids are mostly utilized as detection elements for monitoring the cells pH conditions, rather than as manipulative elements to regulate cell behaviors. Here, we aim to translate pH change into regulation of cellular processes, which may provide a potential way to manipulate cellular responses under different pH conditions.
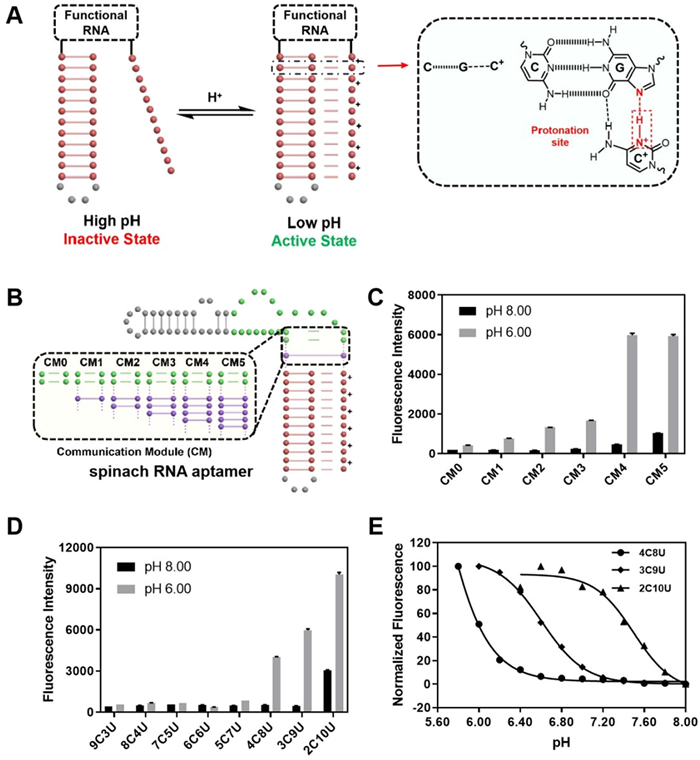
RNA triplex are formed through Hoogsteen interactions, such as C+·G-C, G·G-C, U·A-U and A·A-U [22,23]. As protonated cytosine is necessary for the C+·G-C structure, the triplex-forming strands are often designed as a pH-responsive structure probe. In this work, we incorporated the triplet elements into the guide RNA allowing us to fine-tune triad-mediated gRNA activity through the formation and deformation of Hoogsteen interactions at different pH conditions. Thus, we can combine the gene-manipulation capabilities of CRISPR/Cas9 with programmable triple RNA to give/pioneer the pH response properties of the Cas9 system (Fig. 1A).
Figure 1
Figure 1.
Regulation of functional RNA through the pH-responsive triplex structure. (A) Design schematics of pH-responsive triplexes combined with functional RNA. Protonated cytosine can form C+·G-C through Hoogsteen interactions. When the pH of the system decreases, the triplex structure forms; otherwise, the triplex dissociates. (B) Design schematics of the communication modules (CMs) between the triplex and the core structure of spinach RNA aptamer. (C) The fluorescence intensities of triplex spinach RNA-DFHBI complexes with different CM lengths under pH 6.00 and 8.00. (D) The fluorescence intensities of triplex spinach RNA-DFHBI complexes with the same CM4 but different cytosine content under pH 6.00 and 8.00. (E) Normalized Fluorescence intensities and the fit curves of triplex spinach-DFHBI complexes with different cytosine content (4C8U, 3C9U, and 2C10U) at pH 6.00–8.00. Error bars indicate the standard deviations derived from three independent biological replicates.
Subsequently, we demonstrated the function of CRISPR/Cas9 complex could be precisely controlled and the expressions of target gene could be conditionally manipulated within different pH values, providing a universal approach to respond to pH changes by controlling CRISPR/Cas9 function in mammalian cells.
We first selected a luminescent system in vitro (spinach aptamer DFHBI) to verify the feasibility of triplex RNA regulating by pH response (Fig. S1 in Supporting information) [24]. This aptamer emits stable green fluorescence after binding to a small molecular substrate (DFHBI) at the pH range of 6.00–8.00 (Fig. S2 in Supporting information). Theoretically, the formation of double or three strands with 3′- and 5′- of RNA sequence closed to the position of aptamer binding to the DFHBI can effectively increase the binding ability of the aptamer and substrate. Once the triplet RNA is stably formed, these short stems can promote the binding of the spinach ligand to the aptamer region, and make the spinach ligand fluorescently. We prepared in vitro transcripts of spinach aptamer RNA containing different stem lengths (0–12 bp) and found that stem length shortening below 5 bp was unstable and aptamer fluorescence in spinach was greatly diminished due to decreased binding levels (Fig. S3 in Supporting information).
At the same time, considering that steric or orientation effects may affect the successful transition between the triplet and the aptamer binding region, we introduce a double-stranded region called a communication module (CM, different lengths) to help the stable formation of both structures (Fig. 1B, Table S3 in Supporting information) [25]. As shown in Fig. 1C, spinach aptamers with different lengths of CM had different binding abilities to spinach ligands at different pH values. The higher the fluorescence intensity, the more stable of the triplex stem mediates aptamer. Notably, when the number of base pairs was 4 bp (CM = 4), the relative fluorescence signal change was the largest (FpH 6/FpH 8), indicating that longer CM would cause triplex-spinach RNA itself to be too stable, while short CM would not be stable enough even triplex structure formatting well (Fig. S4 in Supporting information).
We then fixed CM length at 4 base pairs to further verify the stability effects of triplex stem length and triplet composition (Fig. 1D). It was found that when cytosine levels were between 75% and 19% (Table S4 in Supporting information), cytosine content was critical to the pH response, and the more cytosine, the lower the pH response (Fig. 1D). Here we mainly showed the three 12-mer RNA triplex (4C8U, 3C9U, and 2C10U) that are highly responsive to common physiological environments (pH 6–8, Fig. 1E).
The sensitivity of 4C8U to acidic conditions is greater than that to high pH value. When pH > 7, the sensitivity of 2C10U to pH is more significant. Therefore, we can regulate the functional stability of spinach aptamers under different pH conditions by fine-tuning the sequence design of RNA triplets.
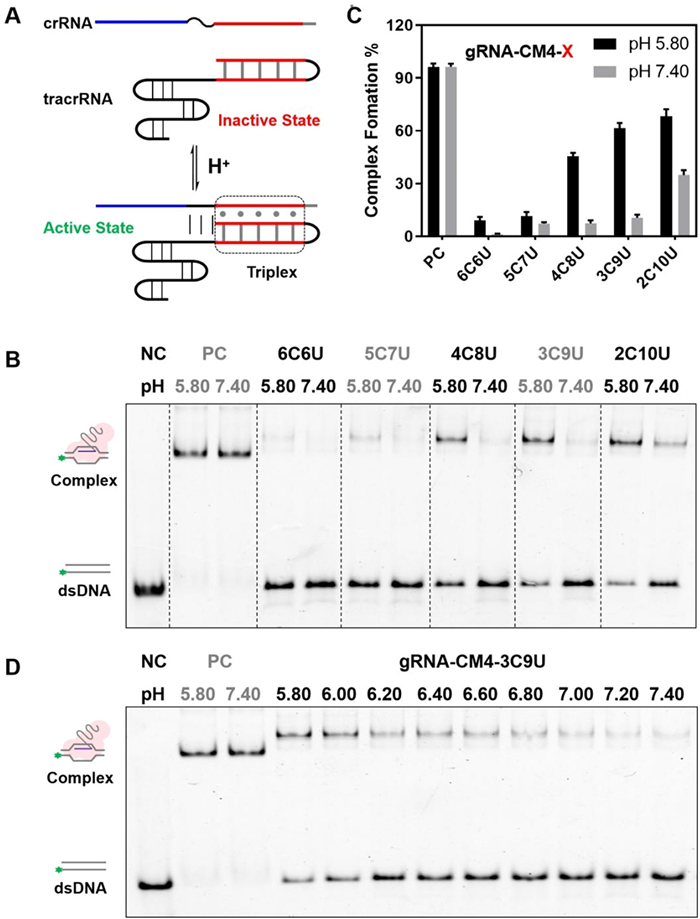
Since the RNA aptamer could be controlled by the formation of triplex, we then wondered whether the same approach could be applied to the CRISPR/Cas9 gRNA to regulate the binding between CRISPR-derived RNA (crRNA) and trans-activating crRNA (tracrRNA). As shown in Fig. 2A, in order to minimize the impacts on its original structure, when introducing the triplex elements into the duplex structure of crRNA/tracrRNA, we kept the triplex outside the protein recognition sites and still retained its effect on the activity of the CRISPR/Cas9 complex (Fig. S5 in Supporting information) [26]. In the native CRISPR/Cas9 system, the original crRNA and tracrRNA can spontaneously bind with each other through complementary base pairing to form active gRNA. When the triplex RNA is introduced into the variable duplex region, the binding behavior between crRNA and tracrRNA could be modulated by the triplex formation. Typically, if the system is under alkaline conditions, the triplex structure is unstable, resulting in loose crRNA and tracrRNA assembly. Whereas, when the pH is reduced to acidic conditions, the triplex structure tends to be stable and inducing the assembled crRNA and tracrRNA to jointly bind with the Cas9 protein to the target DNA. Clearly, this RNA triplex may be employed to the activity of CRISPR/Cas9 complex in a pH-dependent manner.
Figure 2
Figure 2.
pH-responsive of triplex RNA was applied in crRNA/tracrRNA of CRIPSR/Cas9. (A) Design schematics of the triplex at the 3′-crRNA and the 5′-tracrRNA. When the system pH decreases, the triplex structure forms well and induces crRNA/tracrRNA formation and a functional guide RNA. (B) Electrophoretic mobility shift assay (EMSA) of triplex-crRNA/trRNA with different cytosine content at pH 5.80 and 7.40. (C) Statistical analysis of RNA/protein/double-stranded DNA (dsDNA) complex formation. (D) EMSA of the triplex-crRNA/trRNA with gRNA-CM4–3C9U at pH levels ranging from 5.80 to 7.40. Error bars indicate the standard deviations derived from three independent biological replicates.
We firstly analyzed the binding ability of dCas9-crRNA-tracrRNA to the target DNA duplex using gel shift assays to determine the appropriate base pairing numbers of crRNA/tracrRNA (Table S5 in Supporting information). As shown in Fig. S6 (Supporting information), when the base pairing region of crRNA/tracrRNA was reduced to 4 bp, the complex could not be fully formed. Hence, we introduced the triplex RNA structure into this 4-bp crRNA/tracrRNA (CM = 4 bp, a critically stable structure) to restore CRISPR/Cas9 complex function by forming triplex structure (Fig. S6A). What is more, under different physical pH conditions, the target affinity to DNA of initial crRNA-tracrRNA after bind with dCas9 and there are no significant changes for dCas9 binding affinity with pH 5.80–7.40 (Fig. S6B).
Next, we examined the binding situation of five triplex RNA sequences (with different cytosine contents, 6C6U, 5C7U, 4C8U, 3C9U and 2C10U) to the shortened crRNA/tracrRNA (Table S6 in Supporting information). In the native electrophoretic gel experiment (Fig. 2B), these triplexes exhibited a significant pH-responsive to the formation of CRISPR/Cas9 complex. As shown in Fig. 2C, the cytosine content is too high (6C6U and 5C7U) to be sensitive to the pH changes between 5.8 and 7.4. When cytosine content was reduced (4C8U, 3C9U and 2C10U), the modified crRNA/tracrRNA was sensitive to pH changes (pH 5.80 and 7.40). Here NC is negative control means the native without triplex engineering modified gRNA, and the binding ability was not significantly affected under different pH conditions (pH 5.80 and 7.40). Interestingly, 3C9U exhibited the largest responsing transform, with a nearly 6-fold change in structure from pH 7.40 to 5.80 among all these triplex engineering modified RNAs (Fig. 2D). These results clearly indicate that the binding activity of the CRISPR/Cas9 complex could be directly regulated by the introduction of triplex RNA structures.
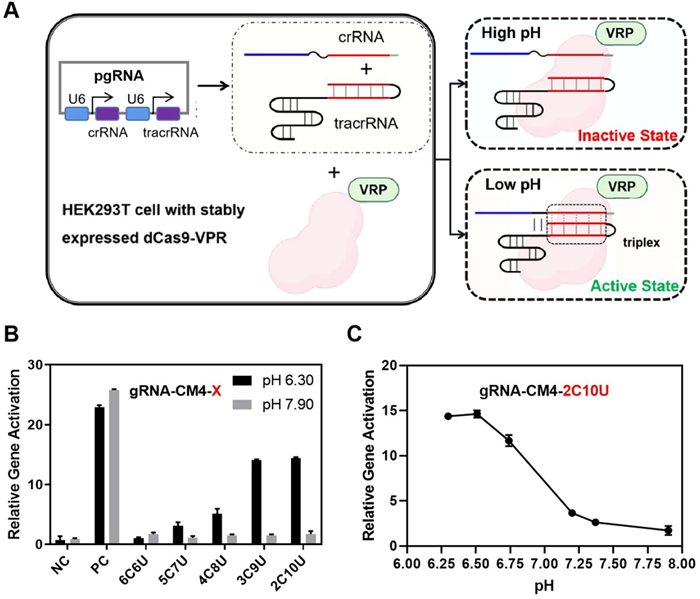
We then tried to further demonstrate that CRISPR/Cas9 function in vivo can be also regulated by environmental pH. We selected the lentivirus-infected HEK293T cell line with stably expressed dCas9-VP64-p65-Rta (VPR) protein for CRISPR-mediated transcriptional activation (CRISPRa) as the model system to check whether there is a regulatory linkage between CRISPRa and triplex mediated pH response [27]. We transfected crRNA and tracrRNA expressing plasmid into HEK293T cells (Fig. 3A). The relative expression level of the target gene ASCL1 was then detected by quantitative polymerase chain reaction (qPCR) to determine the pH response behavior of functional crRNA/tracrRNA. Using the unmodified gRNA as reference, we examined the effects of different RNA triplexes on ASCL1 gene expression levels with different pH conditions.
Figure 3
Figure 3.
pH-controlled gene expression in human cells. (A) Design schematics for pH-induced transcriptional activation in HEK293T cells. (B) Relative transcriptional levels of ASCL1 with different cytosine percentages at pH 6.30 and 7.90. The negative control (NC) indicated the gRNA could not target the ASCL1 gene; the positive control (PC) demonstrated the effect of the standard gRNA without the introduction of the triplex. (C) Relative transcriptional levels of the ASCL1 gene with plasmid triplex gRNA-CM4–2C10U at different pH conditions in HEK293T cells. Error bars indicate the standard deviations derived from three independent biological replicates.
The cellular pH was altered by finetuning the culture medium with phosphate buffer solution (PBS) and the intracellular pH value was determined by commercially available Intracellular pH test kit (BBcellProbe) (Fig. S7 in Supporting information). For the unmodified gRNA, the expression level of ASCL1 was not affected by pH value. In contrast, when the triplex RNA was introduced into the modified crRNA/tracrRNA, the effect of pH environment on ASCL1 expression was clearly observed (Fig. 3B). Among these five RNA triplex structures, 3C9U and 2C10U showed the most dynamic response between pH 6.30 and 7.90 with over 14-fold changes on ASCL1 transcriptional. By systematically measuring ASCL1 expression levels at different pH conditions, we found that this is similar to the previous in vitro biochemical results (Fig. 3B and Fig. S8 in Supporting information). In order to verify the universality of the method, we also tested the regulatory effect of the method on the transcription of another gene, CXCR4 (Table S6), and found that it also had a significant effect. Along with the increase of pH from 6.30 to 7.90, the effect of transcriptional activation for CXCR4 was also gradually enhanced accordingly, revealing a clearly pH-dependent regulation (Fig. S9 in Supporting information). Together, these results suggested that these engineered gRNAs could serve as regulatory elements for gene expressions in the cellular contexts in a pH-dependent pathway.
In conclusion, we integrated the RNA triplex into functional nucleic acids and controlled the activity of guide RNA within the CRISPR/Cas9 complex under different pH conditions. Moreover, with the pH-dependent manipulation of gRNA, we successfully achieved regulation of mammalian gene expressions in different pH conditions through controllable CRISPR/Cas9 function. When the intracellular pH values altered around the neutral condition, the target gene expressions could be significantly modulated. In fact, in addition to the gene expression investigated in this work, other Cas9-based effectors could be also manipulated through the same approach to control their activities under different pH conditions in the cellular contexts. This design established a general approach to manipulate the functionalities of CRISPR/Cas9 complexes in a pH-responsive manner, which further enriched the gene manipulating toolbox with conditional control of RNA functionalities [28].
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
This research was supported by the National Key R&D Program of China (Nos. 2022YFC2804101, 2020YFA0211200), National Natural Science Foundation of China (Nos. 22377056, 22222706, 21977122).
Supplementary materials
Supplementary material associated with this article can be found, in the online version, at doi:10.1016/j.cclet.2023.109427.
Figure 1
Regulation of functional RNA through the pH-responsive triplex structure. (A) Design schematics of pH-responsive triplexes combined with functional RNA. Protonated cytosine can form C+·G-C through Hoogsteen interactions. When the pH of the system decreases, the triplex structure forms; otherwise, the triplex dissociates. (B) Design schematics of the communication modules (CMs) between the triplex and the core structure of spinach RNA aptamer. (C) The fluorescence intensities of triplex spinach RNA-DFHBI complexes with different CM lengths under pH 6.00 and 8.00. (D) The fluorescence intensities of triplex spinach RNA-DFHBI complexes with the same CM4 but different cytosine content under pH 6.00 and 8.00. (E) Normalized Fluorescence intensities and the fit curves of triplex spinach-DFHBI complexes with different cytosine content (4C8U, 3C9U, and 2C10U) at pH 6.00–8.00. Error bars indicate the standard deviations derived from three independent biological replicates.
Figure 2
pH-responsive of triplex RNA was applied in crRNA/tracrRNA of CRIPSR/Cas9. (A) Design schematics of the triplex at the 3′-crRNA and the 5′-tracrRNA. When the system pH decreases, the triplex structure forms well and induces crRNA/tracrRNA formation and a functional guide RNA. (B) Electrophoretic mobility shift assay (EMSA) of triplex-crRNA/trRNA with different cytosine content at pH 5.80 and 7.40. (C) Statistical analysis of RNA/protein/double-stranded DNA (dsDNA) complex formation. (D) EMSA of the triplex-crRNA/trRNA with gRNA-CM4–3C9U at pH levels ranging from 5.80 to 7.40. Error bars indicate the standard deviations derived from three independent biological replicates.
Figure 3
pH-controlled gene expression in human cells. (A) Design schematics for pH-induced transcriptional activation in HEK293T cells. (B) Relative transcriptional levels of ASCL1 with different cytosine percentages at pH 6.30 and 7.90. The negative control (NC) indicated the gRNA could not target the ASCL1 gene; the positive control (PC) demonstrated the effect of the standard gRNA without the introduction of the triplex. (C) Relative transcriptional levels of the ASCL1 gene with plasmid triplex gRNA-CM4–2C10U at different pH conditions in HEK293T cells. Error bars indicate the standard deviations derived from three independent biological replicates.

 DownLoad:
DownLoad:

 下载:
下载:




 下载:
下载:
