High Magnetic Field Laboratory, CAS Key Laboratory of High Magnetic Field and Ion Beam Physical Biology, Hefei Institutes of Physical Science, Chinese Academy of Sciences, Hefei 230031, China
b.
University of Science and Technology of China, Hefei 230026, China
c.
University of California, San Diego, La Jolla, CA 92093, United States
d.
International Magnetobiology Frontier Research Center, Hefei 230031, China
* Corresponding author. E-mail address: makun@hmfl.ac.cn (K. Ma).
Received Date:
26 September 2023 Accepted Date:
19 December 2023 Revised Date:
10 November 2023 Available Online:
15 October 2024
Abstract:
For a significant duration, enhancing the efficacy of cancer therapy has remained a critical concern. Magnetotactic bacteria (MTB), often likened to micro-robots, hold substantial promise as a drug delivery system. MTB, classified as anaerobic, aquatic, and gram-negative microorganisms, exhibit remarkable motility and precise control over their internal biomineralization processes. This unique ability results in the formation of magnetic nanoparticles arranged along filamentous structures in a catenary fashion, enclosed within a membrane. These bacteria possess distinctive biochemical properties that facilitate their precise positioning within complex environments. By harnessing these biochemical attributes, MTB could potentially offer substantial advantages in the realm of cancer therapy. This article reviews the drug delivery capabilities of MTB in tumor treatment and explores various applications based on their inherent properties. The objective is to provide a comprehensive understanding of MTB-driven drug delivery and stimulate innovative insights in this field.
Countless people worldwide succumb to cancer each year, making it a global menace due to its high fatality rate, high metastasis and formidable resistance to treatment [1–6]. Currently, there exist diverse methods for treating cancer, including: (1) Radiotherapy: This method employs radiation to target and eradicate local tumor tissues. While it is a common clinical approach, it proves ineffective against non-radiosensitive tumor regions. (2) Light/magnetic thermotherapy: This technique involves injecting materials with high light conversion rates and targetability into the patient's body. These materials accumulate around the targeted area and destroy tumor tissue through the application of external light or alternating magnetic fields. (3) Immunotherapy: This approach involves artificially modulating the patient's immune system to enhance its ability to combat tumors. (4) Chemotherapy: Chemotherapy relies on anti-tumor chemicals to eliminate cancerous tissues. However, its significant drawback is the extensive damage it inflicts on healthy tissues due to the non-specific nature of drug distribution. The use of nanoparticles for drug delivery has mitigated some of these issues. Nevertheless, the genotypic characteristics of tumor tissues hinder drug penetration, and the targeting precision of certain nanomedicines remains inadequate, with only a fraction reaching the intended destination [7–9]. Concurrently, tumor cells can develop drug resistance following prolonged exposure to low drug concentrations [10–12]. Consequently, terminal cancer patients often develop multiple drug resistances and possess more compact tumor tissues. In such cases, most of the aforementioned treatment approaches become ineffective or yield unacceptable side effects. Thus, there is a pressing need to develop new treatment strategies that enhance therapeutic efficacy and improve the quality of life for patients.
One approach to enhance the effectiveness of tumor therapy involves designing strategies to enhance the reliability and targetability of drug delivery. The efficiency of drug delivery can be hindered by factors such as tissue density, drug-serum protein interactions, phagocytic activity, and vascular permeability [13]. Previous drug carriers have enclosed drugs within various inclusions but often lack stability and precision. This is where engineering microrobots come into play, referring to steerable microparticles that gain kinetic energy from other forms of energy to achieve autonomous drug delivery. Engineering microrobots encompass various entities, including bacteria, immune cells, sperm cells, and more [14–17]. These microrobots exhibit the capability for self-directed motion, allowing for flexible speed regulation. They shield drugs from dilution by bodily fluids while maintaining high penetrability, thereby enabling tailored drug compositions and delivery modes. Furthermore, these microrobots can be monitored in vitro, facilitating precise spatio-temporal control over drug delivery.
1.2
Bacterial delivery
In laboratory cell cultures, tumor cells are relatively straightforward to cultivate compared to normal cells due to their ability for infinite passage and rapid growth. However, they are also highly susceptible to microbial contamination, which hints at an intricate relationship between tumor cells and bacteria. This interaction forms the foundation of a microbial drug delivery system that specifically targets the most chemotherapy-resistant tissues [18]. The growth and survival of bacteria within tumors hinge on factors such as their mode of locomotion, biological characteristics, and, notably, their reliance on oxygen [19,20]. Several bacterial species have been proposed for drug delivery to the core anaerobic regions of tumor tissues, gradually establishing bacteria as a viable solution to the issue of drug specificity in cancer therapy [21]. Commonly used engineered bacteria for tumor therapy include: (1) Salmonella Typhimurium: Despite being a pathogenic bacterium capable of causing gastroenteritis in humans, S. Typhimurium is considered one of the most promising bacteria for cancer immunotherapy. It has also been tailored for targeted cancer therapy due to its ease of manipulation [22,23]. For instance, Yoon et al. developed a Salmonella capsule encapsulating TNF- α, a cytokine that induces apoptosis in tumor cells. This led to an 80%−100% inhibition of tumor growth and prolonged the survival of mice in a melanoma model [24]. Furthermore, Park et al. investigated the motility characteristics of S. Typhimurium when combined with drug-carrying liposomes [25]. However, it should be noted that S. Typhimurium's pathogenicity raises safety concerns, necessitating further exploration for its potential as a drug carrier. (2) Escherichia coli (E. coli): Intravenous injection of genetically engineered E. coli has shown the ability to target and locate anoxic tumor regions. These modified E. coli strains can secrete ClyA for the treatment of colon cancer and melanoma in mice [26]. Alternatively, they can be engineered to carry doxorubicin, release targeted nanobodies, and more [27,28]. (3) Lactic acid bacteria: Lactic acid bacteria exhibit tolerance to low pH levels and possess the capability to migrate to solid tumors following intravenous administration [20]. Additionally, fermented foods containing strains such as Lactobacillus acidophilus, Lactobacillus bulgaricus, Streptococcus lactis, or Bifidobacterium have been shown to suppress breast cancer proliferation. Some of these bacterial strains have also been harnessed as drug carriers [29–31].
Regrettably, clinical data suggest that steering and tracking bacteria using external instruments present formidable challenges. In accordance with the carrier targeting classification, carriers can be categorized into passive targeting (PT), active targeting (AT), and magnetic targeting (MT) [32]. Among these three targeting modalities, MT exhibits superior efficacy compared to AT, which, in turn, surpasses PT. In recent years, there has been a gradual development of numerous targeted drugs incorporating magnetic guidance, yielding highly significant therapeutic outcomes [33–36]. Regarding bacterial microrobots, general anaerobic bacteria possess both PT and AT capabilities, but they lack inherent magnetic responsiveness in their natural state. However, in response to this issue, magnetotactic bacteria (MTB), which function as naturally occurring microrobots, overcome this limitation through their easily controllable magnetic particles. They stand as promising tools for bacterial tumor therapy, offering distinct biophysical properties, especially in the context of targeted therapies, and demonstrating significant potential for practical application. MTB is poised to serve as a highly effective vehicle for drug delivery to specific tissues or organs in vivo [37–40]. This review provides a comprehensive overview of MTB applications in tumor therapy, highlighting their advantages in drug delivery, with the objective of fostering a deeper understanding of the MTB drug delivery system.
2.
Magnetotactic bacteria and their drug delivery advantages
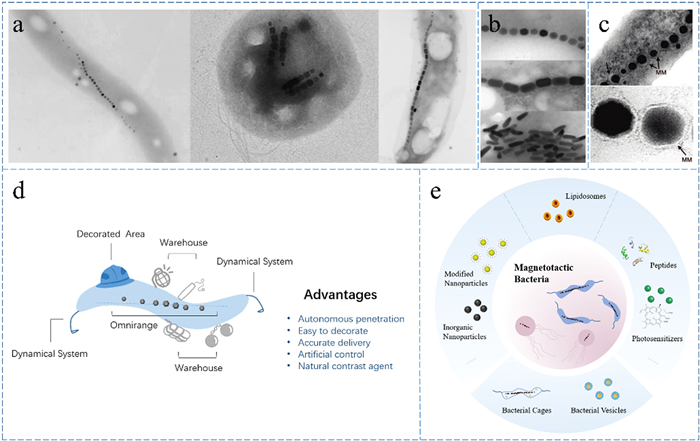
MTB constitute a group of anaerobic aquatic Gram-negative bacteria thriving near distinct redox boundaries in water [41–43]. Over time, the classification of MTB has evolved, with an increasing number of bacterial groups identified by researchers. What initially began with two phyla, Proteobacteria and Nitrospirae, has now expanded to encompass 16 phyla [41]. Remarkably, adaptive MTB have also been discovered in extreme environments characterized by high temperatures and salinity [44,45]. MTB, as depicted in Fig. 1a, display a diverse array of natural morphologies, including arc, sphere, spiral and more. They navigate by orienting themselves along the lines of a magnetic field, employing flagella as their propellers [46–48]. This intriguing motion is orchestrated by the magnetosomes. Magnetosomes come in various shapes, as illustrated in Fig. 1b, and consist of magnetic nanocrystals ensconced within phospholipid membranes, forming chains interconnected by proteins (Fig. 1c) [49,50]. This magnetosome chain functions as an internal compass, directing the movement of MTB. Some exceptional strains of MTB even possess multiple chains, deviating particles, and exhibit variations in the tightness of particle-to-particle connections [51].
Figure 1
Figure 1.
Morphology of MTB and magnetosomes and summary of MTB to deliver drugs. (a) MTB have various shapes including spirochetes, cocci, vibrios, and more. Reprinted with permission [43]. Copyright 2008, American Chemical Society. (b) There are many types of magnetosomes including cuboctahedron, prismatic, bullet-shaped, and more. Reprinted with permission [50]. (c) Magnetosome chains are made up of multiple magnetosomes arranged in rows, each of which is structured as a magnetic nanoparticle encapsulated by membrane, pointed out by the arrows. Reprinted with permission [43]. Copyright 2008, American Chemical Society. (d) MTB act as microrobots, consisting of decorated area, omnirange, dynamical system, and warehouse, and there are numerous advantages to utilizing MTB for drug delivery. (e) Summary of different modes of drug delivery by MTB.
Magnetosomes represent a prominent example of biomineralization, typically measuring 30–50 nm in length. Magnetite and greigite particles within this size range exhibit stable single magnetic domains, retaining their permanent magnetism at room temperature. Microstructural characterization reveals that magnetosomes are essentially devoid of internal defects and exhibit nearly perfect crystallographic features. For instance, a typical cuboctahedron structure consists of the {100} and {111} forms and grows nearly equidimensionally [43]. Furthermore, fine-tuning through biological processes ensures that each magnetic particle attains optimal magnetic properties. The magnetosome chain, formed by the accumulation of individual particles with single magnetic domains, enhances the directional magnetic induction property of MTB. Consequently, MTB are rightfully expected to possess distinctive magnetotaxis capabilities. However, it is worth noting that there are some exceptions where linear chains are not formed.
As delineated in Fig. 1d, MTB are inherently motile and remarkably permeable, thanks to their efficient navigation and power systems. Their performance surpasses that of passive diffusion in various aspects. The presence of magnetosomes within their bodies facilitates in vitro detection and allows for precise artificial magnetic control. Moreover, MTB offer versatility in terms of modification. It is feasible to engineer tailor-made microrobots with multiple actuation modes. For instance, they can be combined with molecular ligands specific to particular tumors, ensuring the safe and accurate delivery of therapeutic drugs to target areas, thereby minimizing damage to healthy tissues. Furthermore, novel molecularly targeted therapeutic tools can be designed and constructed, utilizing warehouse-like MTB as carriers for a range of anti-tumor active substances (Fig. 1e). All in all, MTB are highly efficient and naturally controllable microrobots.
2.2
Drug delivery advantages of magnetotactic bacteria
A substantial body of prior research has primarily focused on utilizing magnetosomes themselves as drug carriers, largely overlooking the potential of considering MTB as a whole [52–54]. As previously mentioned, intravascular injection typically results in a low proportion of drugs actually reaching the tumor site [55]. Consequently, direct intravascular injection at the tumor region is limited in its applicability, making it particularly suitable for certain well-localized tumors like prostate cancer. Additionally, it is imperative to ensure the dispersion of magnetosomes to prevent their aggregation in vivo, which could pose biosafety concerns related to vascular issues.
In comparison to magnetosomes, MTB as drug carriers may offer distinct advantages for several specific reasons: (1) Growth characteristics: The biological properties of MTB align well with the pathological characteristics of solid tumors. Tumor tissues often feature abnormal vessel structures, irregular blood flow, and extend beyond the reach of the blood supply [56,57]. This tumor microenvironment is characterized by conditions such as hypoxia (oxygen concentration ≤ 10 mmHg), slightly acidic pH, and a rich concentration of metabolites, all of which are precisely conducive to MTB. Furthermore, the proliferation ability of MTB within tumors may surpass that in normal tissues [58–63]. (2) Surface proteins for modification: MTB possess an abundant array of proteins on their surface that are readily modifiable and can be linked to various drugs through multiple mechanisms. This facilitates efficient and precise spatiotemporal drug transport, safeguarding drugs from dilution by bodily fluids. (3) Magnetic responsiveness: MTB respond to magnetic fields as their axis of motion [64]. They exhibit robust self-propulsion and can be controlled based on external alterations in magnetic field strength and direction. This allows drugs to be precisely delivered to designated in vivo destinations while avoiding non-targeted tissues [65]. MTB can also penetrate deeper into solid tumors compared to passive diffusion-based drug carriers [66,67]. Ongoing research suggests that a uniformly rotating magnetic field enhances MTB's exploration of the tumor surface and tissue penetration [68]. (4) Biocompatibility: Concerning normal biological tissues, numerous studies have consistently reported that MTB exhibit high biocompatibility. They do not induce apoptosis in normal cells and do not significantly stimulate immune system activation. Injections of MTB with proper concentration into mice have not resulted in obvious inflammation, with biochemical parameters remaining within the normal range [66,67]. (5) Immune sensitization: In the more complex tumor tissue environment characterized by factors like lower pH, hypoxia, and the presence of specific metalloenzymes, MTB tend to proliferate, leading to increased production of bacterial metabolites. Furthermore, this environment may induce the production of unique antigens, which could potentially trigger a stronger immune response from tumor cells [69]. (6) Natural contrast agent: The unique magnetosome structure inherent in MTB serves as a natural T2-weighted contrast agent for positional imaging, obviating the need for additional tagging. This inherent feature may facilitate the in vitro tracking of MTB [50,70–72].
3.
Drug delivery application of magnetotactic bacteria
To date, several delivery models for MTB have been developed, encompassing a range of components such as photosensitizers, peptides, nanoparticles, liposomes, and others, as meticulously outlined in Table 1.
Table 1
Table 1.
Different delivery models of MTB and information including bacteria, clicking strategies, delivery of goods, drugs, applications and references.
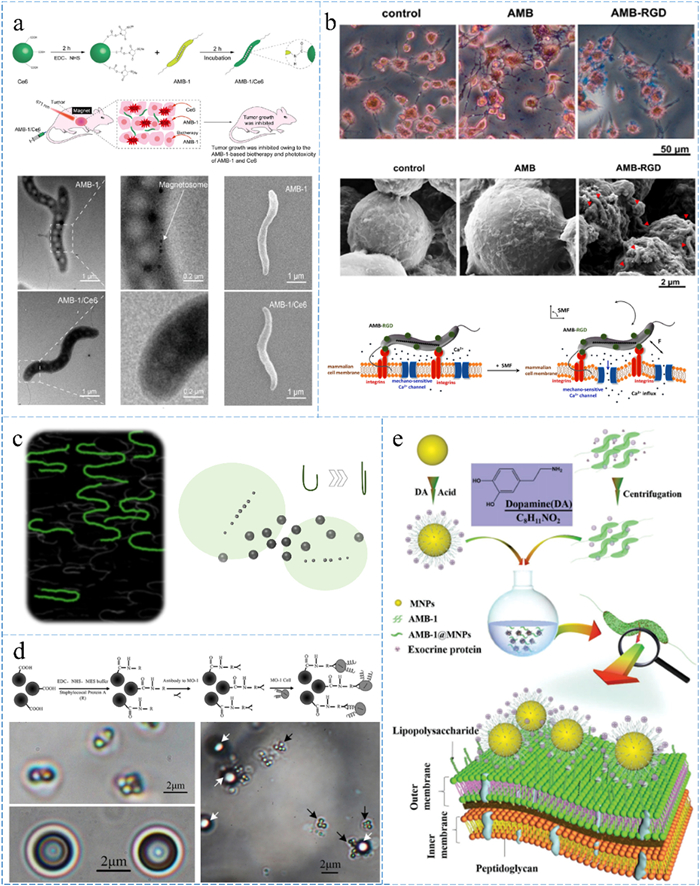
Researchers have directly connected MTB to biomolecules to enable rapid functionalization, as demonstrated by Wang et al., who constructed a structure involving AMB-1 and the photosensitizer Ce6 (Fig. 2a), which exhibited prolonged stability and efficient tumor cell inhibition when subjected to laser irradiation. This complex demonstrated prolonged stability, which holds potential for extended in vivo studies and targeted delivery. Upon laser irradiation, AMB-1/Ce6 produced substantial levels of reactive oxygen species, effectively eradicating tumor cells. Experimental results showcased significant inhibition of tumor growth [73]. Metabolomic studies revealed that the majority of Ce6 is metabolized and eliminated within a day after injection. Interestingly, a simple intravenous injection of AMB-1 significantly enhanced the suppression of tumor growth, with this effect persisting for approximately 14 days post-injection. This phenomenon might be attributed to the competitive nutrient utilization by the proliferating AMB-1 within the tumor microenvironment. Furthermore, it could involve the release of certain metabolites that inhibit tumor growth following the successful targeting of bacteria to the tumor site [74].
Figure 2
Figure 2.
Diagram of MTB-molecules/nanoparticles. (a) AMB-1 and photosensitizers Ce6 are connected via EDC reaction for biotherapy and light therapy, including flow chart and morphology schema. Reprinted with permission [73]. Copyright 2023, American Chemical Society. (b) AMB-1 modificated with RGD specifically targets tumor cells and then induces calcium ion influx by mechanical oscillation under an applied alternating magnetic field. Reprinted with permission [75]. Copyright 2022, Elsevier. (c) MTB carry 200 nm superparamagnetic iron oxide nanoparticles and show increased response speed at U-turns. Reprinted with permission [81,82]. Copyright 2021, American Chemical Society. (d) 2 µm magnetic beads couple to multiple MO-1s by immunoreactivity; white arrows point to magnetic beads, black arrows point to MO-1s. Reprinted with permission [84]. Copyright 2012, AIP Publishing. (e) MTB are deposited with Fe3O4 magnetic nanoparticles on the surface to achieve magnetic control of SAMTB. Reprinted with permission [85]. Copyright 2019, John Wiley and Sons.
Wang et al. introduced an innovative therapeutic approach leveraging MTB and magnetic fields. Through surface modification with FITC-RGD peptides, MTB were capable of targeting and interacting with tumor cells (Fig. 2b). When subjected to an alternating magnetic field, MTB bound to tumor cell surfaces underwent mechanical oscillations, potentially triggering a significant influx of Ca2+ ions and subsequent tumor growth suppression [75]. This research marked the first instance of MTB inhibiting tumor growth by mediating cell signaling regulation, presenting a novel therapeutic tool.
3.2
Inorganic nanoparticles
Numerous studies have sought to engineer MTB to enhance its capabilities as a drug delivery carrier. This primarily involves improving various properties of MTB. The role of magnetic nanoparticles in medical detection and drug delivery is well-established [76–81]. One study proposed the coupling of MTB with superparamagnetic iron oxide nanoparticles to enhance drug delivery (Fig. 2c). The results indicated that MTB-MNPs exhibited a lower moving speed but a higher response speed for U-turns [82]. This heightened sensitivity suggests potential applications in future solid tumor targeting and therapy. A study demonstrated that MTB could induce the migration of micro-objects and assessed their speed performance [83]. Afterward, in another study, Ma et al. developed a novel microrobot by attaching 2 µm microbeads to ovoid marine MTB (MO-1) through immunoreaction (Fig. 2d). This approach significantly increased attachment efficiency by up to 30% through immunoreactivity. Additionally, motility performance assessments revealed that MTB could travel at a speed of 21 µm/s, maintaining continuous movement and viability for over 30 min [84]. This microrobot construction holds promise for applications in medical diagnosis and drug transportation. Furthermore, Li et al. refined a semi-artificial MTB (SAMTB) with tunable magnetic properties achieved by depositing Ferric Oxide on the MTB surface (Fig. 2e). The connection between the synthesized Fe3O4 magnetic nanoparticles and AMB-1 was maintained through electrostatic interactions. Adjustment of SAMTB's magnetic properties was achieved by modifying the concentration, size, and surface charge of Fe3O4 magnetic nanoparticles, allowing precise control of mobility and magnetic sensitivity within the system [85]. This approach serves as a noteworthy example of controlling microrobots through a combination of the material's intrinsic properties and artificial design.
3.3
Liposomes
Prior studies have combined MTB with liposomes to construct a biocompatible microrobot. The number of liposomes attached to a single MTB relies on drug concentration and incubation duration. Notably, the marine MTB (MC-1) can successfully connect with approximately 70 liposomes (Fig. 3a). Although the swim speed of the complex decreases by approximately 27% compared to the wild type, its motility and functionality remain largely intact. Moreover, cytotoxicity tests demonstrate that this modification enhances the biocompatibility of MTB when introduced into the human body [69]. SEM, FACS and confocal microscopy results collectively confirm that exogenous liposomes form robust connections with MTB and are resistant to detachment. Importantly, the majority of liposomes attach to the main body of MTB, with less than 2% adhering to the flagellum. The optimization objective of this system is to attach an ideal number of liposomes without significantly affecting the swimming speed of MTB. Subsequent calculations indicate that if liposome attachment occupies less than 23% of the total surface area of MTB, it does not appreciably impact the mobility of MTB- liposomes or their response to the applied magnetic field.
Figure 3
Figure 3.
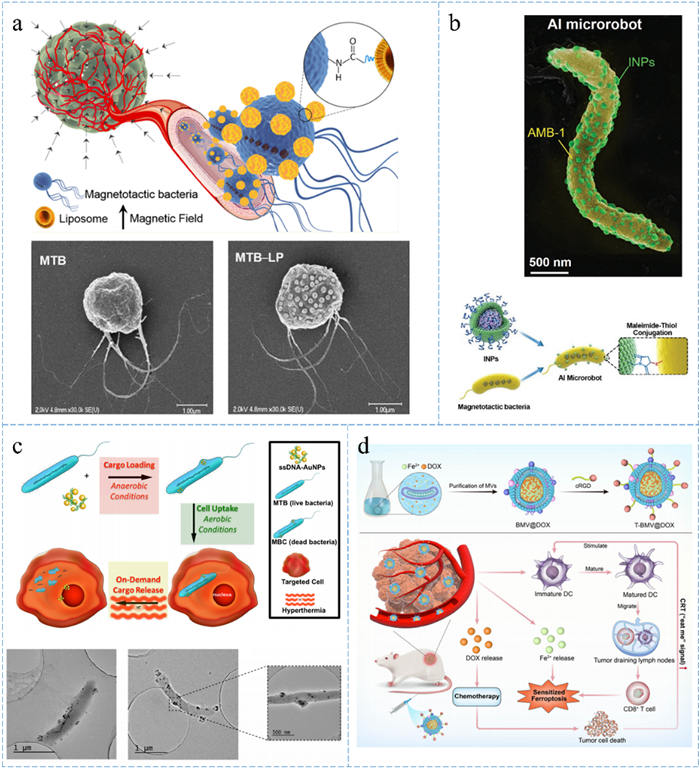
Diagram of MTB-liposomes and other special delivery models (a) MC-1 is connected to about 70 liposomes with an average targeting ratio of greater than 50%, and the molecules inside the liposome can effectively inhibit the growth of tumor cells. Reprinted with permission [66,69]. Copyright 2014, American Chemical Society. (b) Approximately 150 INPs are localized on AMB-1, providing imaging of fluorescence and photothermal therapy. Reprinted with permission [67]. Copyright 2020, John Wiley and Sons. (c) Whole MTB were used as carriers to construct MBC by wrapping ssDNA-AuNPs, switching with and without oxygen. Reprinted with permission [88]. Copyright 2016, Elsevier. (d) Vesicles are obtained from medium containing doxorubicin for synergistic therapy including chemotherapy, ferrotherapy and immunotherapy. Reprinted with permission [89]. Copyright 2023, Royal Society of Chemistry.
Organoids, in vitro cell cultures with three-dimensional structures that closely mimic in vivo environmental conditions, resemble tumors in terms of structure. The inner regions of these cultures exhibit increased compactness, lower pH levels, and reduced oxygen concentrations, forming a gradient with depth. It is precisely within this core region that most chemotherapy drugs struggle to reach and exert their effects. Recent tracking data from embedded imaging systems demonstrate that the marine MTB (MC-1) can penetrate spherical cell cultures constructed from colon cancer cells (LS174T) [86]. This indicates that MTB have the capability to autonomously navigate within tumor environments, functioning as micro robots for drug delivery under computer-controlled magnetic fields. Additionally, Afkhami et al. discovered that enveloping MC-1 in giant vesicles enhances targeting delivery efficiency, illustrating the positive correlation between encapsulation modifications and MTB penetrability [87].
The anti-tumor molecule SN-38 inhibits topoisomerase, impacting DNA molecule synthesis and leading to frequent DNA single-strand breaks. SN-38 is encapsulated in liposomes and then modified with carboxyl residues. These liposomes are subsequently attached to MTB through straightforward incubation or EDC reactions, leveraging the free amino groups on the surface protein of MTB. The effectiveness of this therapy has been evaluated in mice. The constructed complexes are injected into the vicinity of tumors induced by xenografts of human colon cancer cells (HTC 116), with approximately 55% of the modified MC-1 actively penetrating deep into the lesion region to release drugs. Tissue sections reveal that MC-1-liposomes is primarily distributed in the tumor centers, particularly within hypoxic and necrotic regions, achieving an estimated average tumor targeting ratio exceeding 50%. When a magnetic field is concurrently applied, MTB's penetration efficiency can be further augmented, directly targeting tumors [66].
Xing et al. engineered a magneto-actuated and optics-triggered microrobot for targeted cancer therapy, comprising two main components: MTB for autonomous targeting and indocyanine green nanoparticles as fluorescent imaging agents and for photothermal therapy (Fig. 3b). The nanoparticles' structure consists of an indocyanine green PLGA core enclosed by a phospholipid shell. Roughly 150 INPs are localized on the surface of AMB-1 and interconnected. Experiments have been conducted to track these microrobots in vivo using fluorescence and magnetic resonance imaging. Furthermore, these microrobots autonomously migrate into the hypoxic regions within tumors, subsequently executing precise tumor tissue destruction through photothermal therapy. This represents an exemplary model for contemporary imaging diagnostics and targeted therapy [67].
3.4
Other delivery models
The previously discussed section summarizes common drug delivery systems involving MTB. However, there are also unique cases worth mentioning. For instance, Alsaiari et al. employed MTB as carriers for cages [88]. Initially, MTB absorbed ssDNA-AuNPs under anaerobic conditions and were subsequently eliminated under aerobic conditions, leading to the creation of magnetotactic bacterial cages (MBC). These MBCs were then incubated with leukemia cells (THP-1), adhering to the cells and undergoing phagocytosis due to bioadhesion. Subsequently, magnetic thermotherapy was utilized to disintegrate the bacteria, resulting in the release of ssDNA-AuNPs (Fig. 3c). Cytotoxicity and immunogenicity assessments affirmed the biosafety and high efficacy of MBC drug delivery. Moreover, since this drug delivery occurs while the bacteria are alive, maximum loading is achievable through endocytosis. This innovative approach offers new possibilities for various external loadings, and exploring different drug types could further assess its applicability and limitations.
Jiang et al. cultured MTB in a medium containing doxorubicin and harnessed membrane vesicles encapsulating ferric ions and doxorubicin, released by MTB, as nanocarriers. This approach aimed to achieve synergistic cancer therapy that combines chemotherapy, ferrotherapy, and immunotherapy (Fig. 3d). Additionally, vesicles modified with DSPE-PEG-cRGD polypeptides exhibited reduced systemic toxicity and enhanced tumor targetability. Experimental results indicated the effectiveness of this therapy in breast and lung cancer models, effectively suppressing the growth and metastasis of drug-resistant tumors in vivo, thereby offering valuable insights into synergistic tumor therapy [89].
4.
Conclusion and outlook
4.1
Conclusion
Cancer now stands as the primary cause of death for approximately one-sixth of the global population. Given the often severe side effects of chemotherapy and radiotherapy, a pressing challenge in modern medicine revolves around discovering innovative approaches to treating tumors, especially in the context of compact and hypoxic tumor tissues [14,90–96]. To address this challenge and enhance the overall effectiveness of tumor therapy, personalized drug delivery systems are under consideration. Bacteria have demonstrated significant therapeutic potential and have garnered considerable interest across various medical domains. Tumor-targeting bacteria possess distinctive characteristics, including specific growth environments and tumor selectivity, making them ideal candidates for the targeted delivery of therapeutic agents, particularly for cancer treatment. Beyond well-known bacteria like Salmonella and E. coli, MTB also emerge as promising candidates for drug delivery in tumor therapy.
MTB harbor intracellular magnetosome chains, often referred to as "compasses," which have multifaceted applications in environmental biology and medicine, including tumor treatment. MTB can be effectively engineered and coupled with various ligands such as doxorubicin, siRNA, antibodies, and liposomes. This modification enhances their biocompatibility and precision as drug carriers, responsive to magnetic fields. This novel delivery model expands the range of innovative therapeutic options for cancer treatment, with the ultimate goal of achieving comprehensive and highly efficient targeting. However, due to the intrinsic heterogeneity among tumors at the cellular and tissue levels, a singular anti-tumor drug or treatment may not suffice for complete eradication. Consequently, combinations of drugs or the integration of alternative therapeutic modalities may be necessary to achieve more comprehensive anti-tumor therapy. It is important to note that this strategy may not be suitable for certain groups, including: (1) individuals with compromised immune systems, such as patients who have previously undergone specific types of chemotherapy, as their immune responses may be suppressed, rendering them less able to react effectively to bacterial injections; (2) individuals with implanted medical devices like artificial hearts, artificial joints, and other medical prosthetics, as bacteria could potentially colonize these devices, leading to extensive infections [97].
4.2
Limitations and future prospects
The utilization of MTB as drug delivery carriers for tumor therapy requires further evaluation before becoming mainstream. (1) Building credibility: Establishing trust is a critical challenge. Despite bacterial antitumor therapy's early clinical use dating back to the 19th century, it still remains a relatively novel approach in the field of tumor therapy, especially in clinical application [98]. While there are currently no studies demonstrating evident biotoxicity associated with MTB, it is imperative to consider the issue of biocompatibility when designing and utilizing MTB. This consideration should encompass the evaluation of potential immunogenicity, particle size, distribution within various tissues and organs, and the eventual elimination pathway within the body. Research has indicated that mice exhibit increased spleen weights on days 1 and 7 following MTB injection, with subsequent decreases observed on day 21, coinciding with the elimination of most MTB from the body [73]. Moreover, a study has identified MTB within the human intestinal microbiome and suggested that MTB may serve as a potential biological source of magnetite in the human brain [99]. Regarding the metabolism of MTB post-injection, the exact pathway remains unclear. Nevertheless, research has demonstrated that MTB are taken up and internalized by cells. Additionally, different cell types exhibit varying degrees of MTB uptake, with phagocytic cells displaying a greater uptake capacity compared to non-phagocytic cells [69,100]. Hence, it is advisable to conduct comprehensive scientific investigations, including the deletion of possibly toxic genes without compromising MTB's activity, to address the limitations and possibly potential side effects associated with bacterial therapy. This approach will facilitate the enhancement of their applicability and the establishment of credibility in this field. (2) Culture optimization: Precise manipulation of MTB is likely to become an essential aspect of this therapy in the future. Despite their abundance in nature, the cultivation of MTB presents difficulties due to their limited reproductive capacity, demanding survival conditions, specific requirements for oxygen, nutrients, and trace elements, and the complexity of culture solutions. The formation of internal magnetosomes in MTB, a fundamental structure for targeted therapy, is intricately tied to gene regulation and growth conditions [101–106]. When wild-type MTB are transferred to laboratory settings for cultivation, many MTB lose the ability to synthesize magnetosomes. This decline is attributed to reduced migratory requirements in the controlled laboratory environment over successive generations of culture. Additionally, their flagella tend to deteriorate, resulting in a significant reduction in overall motility and responsiveness to magnetic fields. Presently, only a handful of MTB species can be successfully cultured in the laboratory. Therefore, it becomes essential to screen strains more suitable for long-term laboratory culture and gain a deeper understanding of the specific mechanisms underlying magnetosome formation. This knowledge will facilitate the optimization of culture conditions, ultimately leading to an increased survival rate and higher growth potential for MTB. (3) Engineering transformation: There remains a deficit in fundamental research and genetic modification techniques concerning MTB. A comprehensive understanding of MTB's specific physiology could potentially lead to efficient genetic modifications. For instance, it might enable the utilization of MTB's inherent secretory pathway to expel specifically expressed drugs directly, eliminating the need for in vitro drug loading. Consequently, there is a growing need for future investigations into the genetic engineering of MTB, which can greatly facilitate MTB modification efforts.
An ideal MTB therapy approach would involve the use of diverse strains, each with specific modifications and treatments, enabling the attachment or encapsulation of various antitumor drugs tailored to different tumor types and individual patient conditions. Once the drug delivery system is established, it can be combined effectively with other therapeutic methods, offering the potential for highly personalized treatments and optimizing targeted properties for enhanced efficacy. In summary, the potential of MTB in tumor treatment and various applications is undeniable. In the future, we anticipate the design of more clinical trials involving MTB to further explore and harness their therapeutic potential. We eagerly await the development of fully controllable and intelligent MTB microrobots, which could usher in a new era in the battle against cancer.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (No. 3190110313 to K. Ma), Special Foundation of President of the Chinese Academy of Sciences (No. YZJJ2022QN44), HFIPS Director's Fund (Nos. E16CWK123X1; YZJJQY202201), the Heye Health Technology Chong Ming Project (No. HYCMP-2022012 to Y. Wang). A portion of this work was performed on the Steady High Magnetic Field Facility, the High Magnetic Field Laboratory, CAS.
[1]
B.S. Chhikara, K. Parang, Chem. Bio. Lett. 10 (2023) 451.
S. Martel, M. Mohammadi, O. Felfoul, Z. Lu, P. Pouponneau, Int. J. Robotics Res. 28 (2009) 571–582. doi: 10.1177/0278364908100924
[66]
O. Felfoul, M. Mohammadi, S. Taherkhani, et al., Nat. Nanotechnol. 11 (2016) 941–947. doi: 10.1038/nnano.2016.137
[67]
J.H. Xing, T. Yin, S.M. Li, et al., Adv. Funct. Mater. 31 (2021) 2008262. doi: 10.1002/adfm.202008262
[68]
T. Gwisai, N. Mirkhani, M.G. Christiansen, et al., Sci. Robotics 7 (2022) eabo0665. doi: 10.1126/scirobotics.abo0665
[69]
S. Taherkhani, M. Mohammadi, J. Daoud, S. Martel, M. Tabrizian, ACS Nano 8 (2014) 5049–5060. doi: 10.1021/nn5011304
[70]
L. Yan, H.Y. Da, S. Zhang, V.M. López, W.D. Wang, Microbiol. Res. 203 (2017) 19–28. doi: 10.1016/j.micres.2017.06.005
[71]
O. Felfoul, N. Mokrani, M. Mohammadi, S. Martel, in: Proceedings of the Annual International Conference of the IEEE Engineering in Medicine and Biology, 2010, pp. 4367–4370.
[72]
N. Lee, H. Kim, S.H. Choi, et al., Proc. Nat. Acad. Sci. U. S. A. 108 (2011) 2662–2667. doi: 10.1073/pnas.1016409108
[73]
B. Wang, Y.W. Qin, J. Liu, et al., ACS Appl. Mater. Interfaces 15 (2023) 2747–2759. doi: 10.1021/acsami.2c19960
[74]
M. Fdez-Gubieda, J. Alonso, A.García Prieto, et al., J. Appl. Phys. 128 (2020) 070902. doi: 10.1063/5.0018036
K. Ma, S. Xu, T.X. Tao, et al., Proc. Nat. Acad. Sci. U. S. A. 119 (2022) e2211228119. doi: 10.1073/pnas.2211228119
[79]
K. Ma, Z. Wang, T.X. Tao, et al., ACS Appl. Nano Mater. 5 (2022) 7410–7417. doi: 10.1021/acsanm.2c01392
[80]
S.H. Ji, Y.D. Chen, X.L. Zhao, et al., Biomater. Sci. 9 (2021) 2732–2742. doi: 10.1039/d0bm01993c
[81]
H. Chen, H.M. Zhang, T.T. Xu, J.F. Yu, ACS Nano 15 (2021) 15625–15644. doi: 10.1021/acsnano.1c07363
[82]
Y. Majedi, D. Loghin, M. Mohammadi, S. Martel, in: Proceedings of the International Conference on Manipulation, Automation and Robotics at Small Scales (MARSS), 2017, pp. 1–6.
[83]
S. Martel, C.C. Tremblay, S. Ngakeng, G. Langlois, Appl. Phys. Lett. 89 (2006) 233904. doi: 10.1063/1.2402221
Q.L. Li, H.T. Chen, X.Y. Feng, et al., Small 15 (2019) 1900427. doi: 10.1002/smll.201900427
[86]
N. Mokrani, O. Felfoul, F.A. Zarreh, et al., in: Proceedings of the Annual International Conference of the IEEE Engineering in Medicine and Biology, 2010, pp. 4371–4374.
[87]
F. Afkhami, S. Taherkhani, M. Mohammadi, S. Martel, in: Proceedings of the Annual International Conference of the IEEE Engineering in Medicine and Biology Society, 2011, pp. 6668–6671.
A. Dieudonné, D. Pignol, S. Prévéral, Appl. Microbiol. Biotechnol. 103 (2019) 3637–3649. doi: 10.1007/s00253-019-09728-9
[105]
U. Heyen, D. Schüler, Appl. Microbiol. Biotechnol. 61 (2003) 536–544. doi: 10.1007/s00253-002-1219-x
[106]
D. Faivre, N. Menguy, M.l. Pósfai, D. Schüler, Am. Mineralog. 93 (2008) 463–469. doi: 10.2138/am.2008.2678
Figure 1
Morphology of MTB and magnetosomes and summary of MTB to deliver drugs. (a) MTB have various shapes including spirochetes, cocci, vibrios, and more. Reprinted with permission [43]. Copyright 2008, American Chemical Society. (b) There are many types of magnetosomes including cuboctahedron, prismatic, bullet-shaped, and more. Reprinted with permission [50]. (c) Magnetosome chains are made up of multiple magnetosomes arranged in rows, each of which is structured as a magnetic nanoparticle encapsulated by membrane, pointed out by the arrows. Reprinted with permission [43]. Copyright 2008, American Chemical Society. (d) MTB act as microrobots, consisting of decorated area, omnirange, dynamical system, and warehouse, and there are numerous advantages to utilizing MTB for drug delivery. (e) Summary of different modes of drug delivery by MTB.
Figure 2
Diagram of MTB-molecules/nanoparticles. (a) AMB-1 and photosensitizers Ce6 are connected via EDC reaction for biotherapy and light therapy, including flow chart and morphology schema. Reprinted with permission [73]. Copyright 2023, American Chemical Society. (b) AMB-1 modificated with RGD specifically targets tumor cells and then induces calcium ion influx by mechanical oscillation under an applied alternating magnetic field. Reprinted with permission [75]. Copyright 2022, Elsevier. (c) MTB carry 200 nm superparamagnetic iron oxide nanoparticles and show increased response speed at U-turns. Reprinted with permission [81,82]. Copyright 2021, American Chemical Society. (d) 2 µm magnetic beads couple to multiple MO-1s by immunoreactivity; white arrows point to magnetic beads, black arrows point to MO-1s. Reprinted with permission [84]. Copyright 2012, AIP Publishing. (e) MTB are deposited with Fe3O4 magnetic nanoparticles on the surface to achieve magnetic control of SAMTB. Reprinted with permission [85]. Copyright 2019, John Wiley and Sons.
Figure 3
Diagram of MTB-liposomes and other special delivery models (a) MC-1 is connected to about 70 liposomes with an average targeting ratio of greater than 50%, and the molecules inside the liposome can effectively inhibit the growth of tumor cells. Reprinted with permission [66,69]. Copyright 2014, American Chemical Society. (b) Approximately 150 INPs are localized on AMB-1, providing imaging of fluorescence and photothermal therapy. Reprinted with permission [67]. Copyright 2020, John Wiley and Sons. (c) Whole MTB were used as carriers to construct MBC by wrapping ssDNA-AuNPs, switching with and without oxygen. Reprinted with permission [88]. Copyright 2016, Elsevier. (d) Vesicles are obtained from medium containing doxorubicin for synergistic therapy including chemotherapy, ferrotherapy and immunotherapy. Reprinted with permission [89]. Copyright 2023, Royal Society of Chemistry.
Table 1.
Different delivery models of MTB and information including bacteria, clicking strategies, delivery of goods, drugs, applications and references.

 DownLoad:
DownLoad:


 下载:
下载:




 下载:
下载:
