Advanced Research Institute of Multidisciplinary Science, School of Life Science, Key Laboratory of Molecular Medicine and Biotherapy, Beijing Institute of Technology, Beijing 100081, China
b.
BSL-3 Laboratory (Guangdong), Guangdong Provincial Key Laboratory of Tropical Disease Research, School of Public Health, Department of Laboratory Medicine, Zhujiang Hospital, Southern Medical University, Guangzhou 510280, China
c.
National Clinical Research Center for infectious disease, Shenzhen Third People's Hospital, Second Hospital Affiliated to Southern University of Science and Technology, Shenzhen 518112, China
yangmh17@bit.edu.cn (M. Yang). 1 These authors contributed equally to this work.
Received Date:
30 August 2023 Accepted Date:
29 October 2023 Revised Date:
25 October 2023 Available Online:
15 August 2024
Abstract:
The monkeypox virus (MPXV) outbreak, declared a Public Health Emergency of International Concern (PHEIC) by the World Health Organization (WHO) in 2022, continues to pose a significant threat due to the absence of vaccines or drugs for MPXV infection. In this study, we developed an mRNA vaccine that expressing the A29L antigen, a specific protein of the intracellular mature virus. Our vaccine utilizes a thermostable ionizable lipid nanoparticle (iLNP) platform and has been administered to mice. Our findings demonstrate that the MPXV A29L mRNA vaccine candidate induces robust cross-neutralizing immune responses against both vaccinia virus (VACV) and MPXV live virus. Furthermore, immunization with the vaccine candidate provided protection against the VACV challenge in mice. These findings underscore the potential of mRNA-LNP vaccines as safe and effective candidates against monkeypox epidemics. Given the current absence of specific interventions for MPXV infection, our study represents a significant step forward in developing a viable solution to combat this ongoing public health threat.
Monkeypox, a zoonotic viral disease caused by the monkeypox virus (MPXV), has become a global concern. Since May 2022, a multi-country outbreak of monkeypox has been identified spanning various geographical regions. This outbreak prompted the World Health Organization (WHO) to declare it a Public Health Emergency of International Concern (PHEIC) [1]. From June 1st to July 31st, 2023, mainland China reported 597 new cases of confirmed monkeypox [2]. The MPXV is a member of the Orthopoxvirus genus in the DNA virus family, which also includes viruses like vaccinia virus (VACV), variola virus (VARV), cowpox (CPXV), and rabbitpox virus (RPXV) [3]. Phylogenetic analysis has revealed two distinct clades of MPXV: the Central Africa clade, known as the Congo Basin strain causing severe symptoms and the West Africa strain, typically leading to milder symptoms [4,5]. Despite these differences, most protein-coding genes remain highly conserved among orthopoxviruses [6]. Despite the global persistence of the monkeypox epidemic for over a year, there are currently no vaccines or specific antiviral drugs available for MPXV.
Prior studies have documented the utilization against smallpox vaccines for other orthopoxvirus infections, suggesting the presence of cross-protective immunity in both animals and humans [7–10]. Data from surveillance during the November 2005 to November 2007 outbreaks indicated that 3.8% of monkeypox cases showed signs of prior smallpox vaccination, as opposed to 26.4% in the general population [11]. Consequently, two live-attenuated VACV-based vaccines (ACAM2000 and JYNNEOS), initially developed as part of the smallpox eradication program, have been approved by the U.S. Food and Drug Administration (FDA) to prevent MPXV [5]. Nevertheless, these vaccines have been associated with notable adverse effects, including injection site reactions, myalgia, headache, and fatigue [12–15]. Consequently, there is an urgent need for novel vaccines that offer enhanced safety, efficacy, and specific targeting against MPXV.
The mRNA technology has played a pivotal role in the remarkable success of coronavirus disease 2019 (COVID-19) mRNA vaccines. These vaccines were stabilized using new mRNA technology and lipid nanoparticles (LNPs) formulation for delivery and translation into the S protein in vivo, effectively triggering an immune response [16–21]. In comparison to traditional vaccines, such as inactivated-virus, live attenuated, recombinant protein, and adenovirus vector vaccines, mRNA vaccines have garnered increasing attention due to its great application prospects and advantages, which include short development cycle, easy industrialization, simple production process, flexibility to respond to new variants, and the capacity to induce better immune response [22,23]. Inspired by this success, the development of an MPXV-specific mRNA vaccine emerges as the most promising strategy to combat the MPXV outbreak. Like other orthopox-viruses, MPXV exists in two infectious forms: the intracellular mature virion (IMV) and the extracellular enveloped virion (EEV), both capable of causing disease [24,25]. Among the orthologous homologous to VACV, the A27L antigen is a well-known target for neutralizing antibodies against IMV [7,26]. In this study, MPXV mRNA-LNP vaccines were developed using A29L (which shares a high degree of homology, 94%, with A27L in VACV) as the antigenic target. We conducted experiments to evaluate the immune response induced by individual vaccine injections as well as their combination in mice.
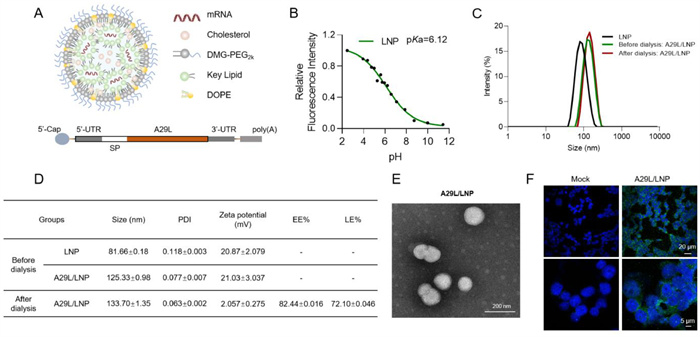
To develop a comprehensive and effective vaccine against MPXV, we designed an mRNA vaccine encoding the full-length A29L gene of the virus. The modified mRNA molecules included a 5′-cap, followed by the 5′ untranslated region (5′-UTR), coding region, and 3′-UTR, ending with a poly(A) tail. Additionally, we incorporated a human interleukin-2 (IL-2) signal peptide (MYRMQLLSCIALSLALVTNS) at the N-terminus of the A29L mRNA to enhance the expression of its protein outside the cell and promote an immune response against the antigen. The A29L mRNA was transcribed in vitro and then encapsulated into ionizable lipid nanoparticles (iLNPs) [27] to form the final A29L/LNP formulation, as illustrated in Fig. 1A. The pKa value of A29L/LNP was determined to be 6.12 (Fig. 1B), which aligns with our expectations. Typically, to achieve effective release, the pKa value of iLNPs should fall between 5.5 and 6.5. This finding indicated that A29L/LNP could efficiently escape from endosomes/lysosomes and exert its biological functions. The particle size of empty LNP was approximately 81.66 nm, while that of A29L/LNP was around 133.70 nm. The polydispersity index (PDI) was below 0.2, suggesting a narrow size distribution of the particles (Figs. 1C and D). Importantly, the non-dialyzed empty vector LNP and A29L/LNP exhibited no significant differences in particle size and PDI compared to those after dialysis, confirming that the addition of mRNA and dialysis did not affect the fundamental structure of the LNPs. Additionally, A29L/LNP demonstrated high encapsulation efficiency (82.44%) and loading efficiency (72.10%), indicating its favorable loading capacity. The morphology of A29L/LNP observed by transmission electron microscopy (TEM) exhibited uniform spherical nanoparticles with a size of approximately 130 nm (Fig. 1E). Collectively, these results demonstrated that A29L/LNP possessed favorable physical and chemical properties, making it suitable for subsequent cellular and animal studies. To confirm the successful translation and expression of A29L, immunofluorescence was conducted using serum from a mouse immunized with A29L recombinant subunits as the primary antibody. The results showed successful expression of A29L in cells transfected with A29L/LNP (Fig. 1F).
Figure 1
Figure 1.
Design and encapsulation of the A29L/LNP mRNA vaccine. (A) Schematic representation of the A29L/LNP mRNA vaccine. (B) Determination of the pKa value for A29L/LNP. (C) Particle size analysis of A29L/LNP before and after dialysis. (D) Particle size, PDI statistics and the detection of encapsulation and loading rates of LNP-mRNA complexes. (E) The TEM image shows the morphology of A29L/LNP mRNA vaccine particles. (F) Immunofluorescence showed the expression of A29L mRNA in vitro.
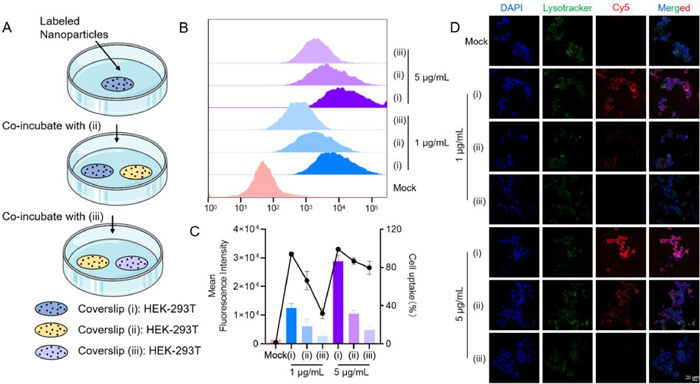
To validate the transcytosis capability of A29L/LNP, we conducted a coverslip transfer experiment. In a nutshell, HEK-293T cells were subjected to transfection with LNP containing Cy5-labeled nucleic acid (Cy5-NA) for a duration of 8 h, followed by co-cultivation with non-transfected cells, as illustrated in Fig. 2A. In Figs. 2B and C, HEK-293T cells were transfected with LNP/Cy5-NA at concentrations of 1 and 5 µg/mL. Notably, the Cy5 signals were observed in (ⅰ), (ⅱ), and (ⅲ). The Cy5 signal in (ⅱ) emanated from cells in (ⅰ) and subsequently underwent endocytosis by (ⅱ). Furthermore, the Cy5 signal in (ⅲ) originated from cells in (ⅱ) and was absorbed by (ⅲ).
Figure 2
Figure 2.
Transcytosis capability of LNP. (A) Diagram of transcytosis. (B, C) Transcytosis efficiency of LNP/Cy5-nucleic acid in HEK-293T cells measured by flow cytometry and quantitative analysis, all of the data are presented as the mean ± SEM. Statistical difference was analyzed by Student's t-test (n = 3). (D) Visualization and quantification of Cy5 signaling for transcytosis capacity of LNP/mRNA complexes in HEK-293T cells utilizing CLSM, scale bar: 20 µm.
The results obtained from confocal laser scanning microscopy (CLSM) and fluorescence-activated cell sorting (FACS) were consistently aligned, as depicted in Fig. 2D. These experimental findings unequivocally affirm the intercellular transfer of nanoparticles across distinct coverslips, thereby validating the transcytosis-inducing potential of A29L/LNP.
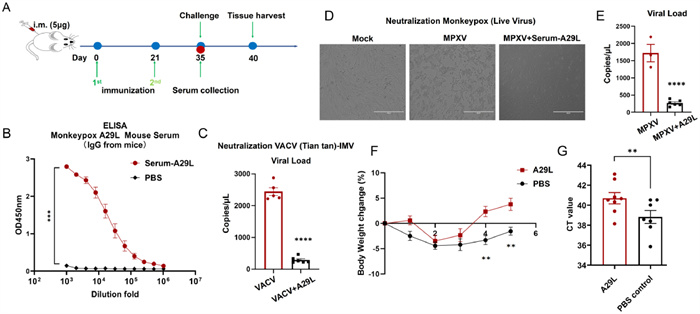
Subsequently, we evaluated the immunogenicity and efficacy of the A29L/LNP vaccine in mice. All experimental procedures were approved by the Animal Care Committees of Beijing Institute of Technology. Female BALB/c mice aged 5–6 weeks were segregated into two groups (n = 7 in PBS group and 8 in A29L group) and immunized at 21-day intervals (Fig. 3A). Mouse sera from subjects immunized with the MPXV A29L mRNA vaccine exhibited a positive binding response to MPXV A29L in the enzyme-linked immunosorbent assay (ELISA), as shown in Fig. 3B. Furthermore, these sera demonstrated significant neutralizing activity against the IMV strain of VACV Tiantan (Fig. 3C). The neutralizing activity was assessed using mouse sera at 20-fold dilutions, with the results indicating substantial efficacy against the IMV strain of VACV Tiantan. In relation to the MPXV live virus, the sera from immunized mice not only markedly inhibited MPXV-induced cellular cytopathic effects (Fig. 3D) but also substantially reduced the viral load post-MPXV infection (Fig. 3E). To further gage the in vivo protective efficacy of the vaccine, immunized mice were subsequently infected with the VACV Tiantan strain. In comparison to the control group, the immunized mice exhibited accelerated weight recovery, indicating that this mRNA vaccine can induce protective immune responses in vivo (Fig. 3F). Additionally, using qPCR technology, we quantified viral loads in the lungs of mice. The results revealed higher cycle threshold (CT) values in the immunized mice as compared to the control group. A higher CT value corresponds to a lower viral copy number, signifying a heightened capacity to clear the virus in the immunized group of mice. Consequently, this linear mRNA vaccine demonstrates a measure of in vivo protective efficacy (Fig. 3G).
Figure 3
Figure 3.
Humoral immune response in A29L/LNP-vaccinated mice against MPXV and cross-protection against VACV challenge in mice. (A) Schedule of immunization and viral infection in mice. (B) Measurement IgG antibody titers specific to MPXV antigen A29L using ELISA, PBS group (n = 7), A29L group (n = 8). (C) Evaluation of neutralizing activity against the IMV strain of VACV Tiantan. (D) Observation of cellular cytopathic effects after MPXV infection. (E) Evaluation of neutralizing activity against the IMV strain of MPXV. (F) The body weight records of vaccinated mice after VACV challenge. (G) Detection of VACV live virus detection in the lungs using qPCR. i.m.: Intramuscular injection; PBS: Phosphate-buffered saline. All of the data are presented as the mean ± SEM. Statistical difference was analyzed by Student's t-test. **P < 0.01, ***P < 0.005, ****P < 0.0001.
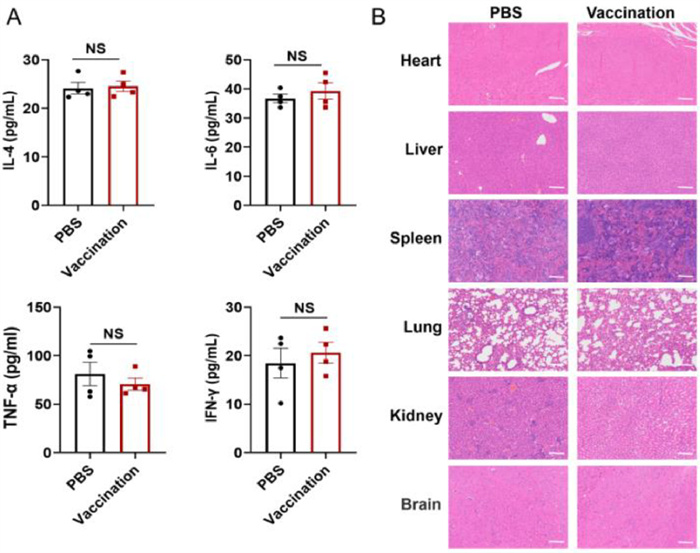
We conducted an in vivo safety assessment of our vaccine. The animals that received the vaccine displayed a consistent general status, resembling that of the control mice, with regards to appearance, eating habits, and mobility. No local skin reactions or signs of inflammation at the injection site were observed post-vaccination. Furthermore, we assessed innate cytokine responses and examined histopathological changes following immunization. Sera were collected from all mice 48 h after the initial vaccination to measure cytokine levels associated with the innate immune response. Specifically, we quantified the secretion levels of IL-6, tumor necrosis factor alpha (TNF-α), IL-4 and interferon‐gamma (IFN‐γ) in both immunized and naive mice. The results revealed no significant differences in cytokine levels between the immunized mice and naive mice at 48 h (Fig. 4A). This indicates that the administration of A29L/LNP did not induce an inflammatory response in the mice. Additionally, we conducted histopathological examinations on various tissues from both vaccination and control groups. These examinations revealed no pathological alterations, further affirming the safety profile of our vaccine in mice (Fig. 4B). In conclusion, multiple lines of evidence support the conclusion that our vaccine possesses a satisfactory safety profile.
Figure 4
Figure 4.
Safety evaluation of A29L/LNP mRNA vaccine in mice. (A) Plasma cytokines detection in innate immunity (n = 4). (B) Representative hematoxylin and eosin (H&E) stains of different tissues, including liver, heart, spleen, lung, kidney and brain from naive mice or mRNA vaccine-immunized mice. The H&E stained sections displayed in the data represent the results from three test mice 48 h post-inoculation, Scale bar: 100 µm. All of the data are presented as the mean ± SEM. Statistical difference was analyzed by Student's t-test. NS: Not significant.
The widespread use of the live VACV has played a pivotal role eradicating smallpox. However, research on VACV vaccines has declined over time. Currently, the approved vaccines for MPXV infection prevention, such as JYNNEOS, ACAM 2000, and LC16m8, were originally designed for smallpox prevention and treatment [28]. In our current study, we have developed an mRNA-LNP vaccine candidate against MPXV encoding A29L, one of the IMV surface proteins. Our vaccine candidate has demonstrated its ability to generate high levels of neutralizing antibodies against both orthopoxviruses and live MPXV in vitro. This suggests that our mRNA vaccine not only provides effective protection against MPXV but also offers cross-protection against VACV infections. Subsequently, we undertook the in vivo infection experiments to substantiate our findings. Consistent with the results of other reported MPXV mRNA vaccines [29,30], mice immunized with the A29L mRNA displayed effective resistance against VACV infection. This was evidenced by the mitigation of virus-induced weight loss and reduced viral load in the lungs.
LNPs have emerged as successful delivery systems for nucleic acid therapeutics due to their established manufacturing technology, high cellular uptake, low toxicity, and minimal immunogenicity. However, clinically applicable LNPs, particularly those with thermostable lipids, are still limited in availability. Notably, the two COVID-19 mRNA vaccines currently in use require storage at very low temperatures, ranging from −25 ℃ to −80 ℃ [31,32]. In our study, the ionizable LNP used in our MPXV vaccine was carefully selected and has demonstrated superior thermostability compared to SM-102, as shown in our previous work [27]. This improved stability enhances the shelf life of our mRNA vaccine formulation. Additionally, in vitro experiments have indicated that our LNP-mRNA vaccine has good transcytosis ability (Fig. 2), potentially contributing to enhanced immune responses in both deep muscle tissue and surface regions.
Regarding safety evaluation, our A29L/LNP mRNA vaccine injection did not result in significant adverse reactions, underscoring its safety. Furthermore, unlike the ACAM2000 vaccine, which often leads to skin reactions at the injection site, and VACV-based vaccines associated with adverse events, including a 0.6% risk of myocarditis according to clinical trial data [30,33], we did not observe any abnormal skin reactions at the injection site with our A29L/LNP vaccine. This suggests that our vaccine does not provoke a significant cutaneous reaction.
One limitation of our study is that we did not explore other antigen mRNAs as vaccine candidates or combinations of multiple antigens. However, the A29L antigen has been shown to induce high levels of neutralizing antibodies [34–36], which is consistent with our study. To date, DNA vaccines and viral vector-based vaccines against MPXV have demonstrated protection against lethal MPXV infection in animal models, including rhesus macaques and 129 stat1−/− mice [7,37], However, our data from BALB/c mice only demonstrate that our MPXV mRNA vaccine candidate effectively prevents weight loss and reduces viral load in the lungs after VACV infection. Further testing in other animal models, especially nonhuman primates challenged with circulating MPXV strains, is warranted.
Overall, we have successfully developed and evaluated an mRNA vaccine candidate against monkeypox, and the promising efficacy data from mice support the further development of this mRNA vaccine platform.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
We knowledge the Beijing Institute of Technology Research Fund Program for Young Scholars. We thank the Biological and Medical Engineering Core Facilities, and Analysis & Testing Center, Beijing Institute of Technology for supporting experimental equipment, and staffs for valuable help with technical support.
This work was supported by National Key Research & Development Program of China (Nos. 2021YFA1201000 and 2021YFC2302400), Beijing Institute of Technology Research Fund Program for Young Scholars (No. XSQD-6120220072), National Natural Science Foundation of China (No. 82371846) and China Postdoctoral Science Foundation (No. 2022M720438).
Supplementary materials
Supplementary material associated with this article can be found, in the online version, at doi:10.1016/j.cclet.2023.109270.
[1]
M.E. DeWitt, C. Polk, J. Williamson, et al., EClin. Med. 54 (2019) 101710.
[2]
China CDC Rep. (9 Augest 2023) https://www.chinacdc.cn/jkzt/crb/qt/szkb_13037/gwjszl_13092/202308/t20230809_268502.html.
Y. Jiang, Z. Ren, Y. Yang, et al., J. Med. Virol. 95 (2023) e28983. doi: 10.1002/jmv.28983
[36]
M. Li, Z. Ren, Y. Wang, et al., Emerg. Microbes Infect. 2 (2023) 2223669.
[37]
V. Franceschi, S. Parker, S. Jacca, et al., PLoS Negl. Trop. Dis. 9 (2015) e0003850. doi: 10.1371/journal.pntd.0003850
Figure 1
Design and encapsulation of the A29L/LNP mRNA vaccine. (A) Schematic representation of the A29L/LNP mRNA vaccine. (B) Determination of the pKa value for A29L/LNP. (C) Particle size analysis of A29L/LNP before and after dialysis. (D) Particle size, PDI statistics and the detection of encapsulation and loading rates of LNP-mRNA complexes. (E) The TEM image shows the morphology of A29L/LNP mRNA vaccine particles. (F) Immunofluorescence showed the expression of A29L mRNA in vitro.
Figure 2
Transcytosis capability of LNP. (A) Diagram of transcytosis. (B, C) Transcytosis efficiency of LNP/Cy5-nucleic acid in HEK-293T cells measured by flow cytometry and quantitative analysis, all of the data are presented as the mean ± SEM. Statistical difference was analyzed by Student's t-test (n = 3). (D) Visualization and quantification of Cy5 signaling for transcytosis capacity of LNP/mRNA complexes in HEK-293T cells utilizing CLSM, scale bar: 20 µm.
Figure 3
Humoral immune response in A29L/LNP-vaccinated mice against MPXV and cross-protection against VACV challenge in mice. (A) Schedule of immunization and viral infection in mice. (B) Measurement IgG antibody titers specific to MPXV antigen A29L using ELISA, PBS group (n = 7), A29L group (n = 8). (C) Evaluation of neutralizing activity against the IMV strain of VACV Tiantan. (D) Observation of cellular cytopathic effects after MPXV infection. (E) Evaluation of neutralizing activity against the IMV strain of MPXV. (F) The body weight records of vaccinated mice after VACV challenge. (G) Detection of VACV live virus detection in the lungs using qPCR. i.m.: Intramuscular injection; PBS: Phosphate-buffered saline. All of the data are presented as the mean ± SEM. Statistical difference was analyzed by Student's t-test. **P < 0.01, ***P < 0.005, ****P < 0.0001.
Figure 4
Safety evaluation of A29L/LNP mRNA vaccine in mice. (A) Plasma cytokines detection in innate immunity (n = 4). (B) Representative hematoxylin and eosin (H&E) stains of different tissues, including liver, heart, spleen, lung, kidney and brain from naive mice or mRNA vaccine-immunized mice. The H&E stained sections displayed in the data represent the results from three test mice 48 h post-inoculation, Scale bar: 100 µm. All of the data are presented as the mean ± SEM. Statistical difference was analyzed by Student's t-test. NS: Not significant.

 DownLoad:
DownLoad:

 下载:
下载:





 下载:
下载:
