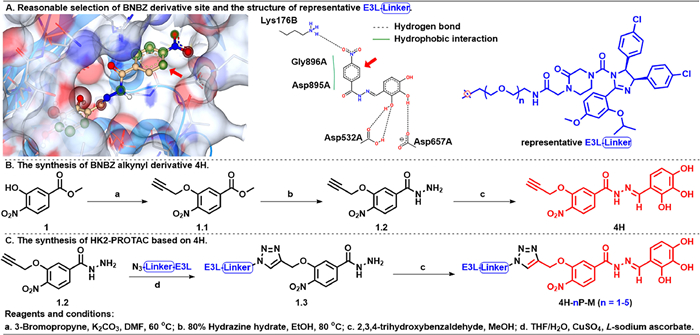
Figure 1.
The selection of HK2 ligand derivatization site and the synthesis route of representative HK2-PROTAC molecules.
A selective HK2 degrader suppresses SW480 cancer cell growth by degrading HK2
Yang Liu , Yan Liu , Kaiyin Yang , Zhiruo Zhang , Wenbo Zhang , Bingyou Yang , Hua Li , Lixia Chen

Cancer cells mainly rely on aerobic glycolysis to meet the metabolic needs of cell proliferation [1,2]. Hexokinase 2 (HK2) plays an important role in aerobic glycolysis of cancer cells, which catalyzes the first step of glycolysis by phosphorylating glucose to glucose-6-phosphate. Studies show that HK2 antagonizes the proapoptotic proteins of the Bcl-2 family such as Bax and Bak, and binds to voltage-dependent anion channel (VDAC) to stabilize the mitochondrial outer membrane, which inhibits the apoptosis of cancer cells [3–5]. In addition, HK2 expression level is closely associated with tumorigenesis, progression, metastasis, and treatment resistance of various malignancies. It can be upregulated by insulin, glucose, and hypoxia [6,7] and be overexpressed in various solid tumors [8–10]. HK2 has been recognized as a promising target for cancer therapy.
In our previous study, benserazide, a DOPA decarboxylase inhibitor used to treat Parkinson's disease, was identified as a lead compound, displaying inhibitory activity against HK2 [11]. Benitrobenrazide (BNBZ), a superior HK2 inhibitor, was designed and synthesized based on the structure of benserazide. It showed significant anti-tumor activity in vitro and in vivo [12]. Although HK2 has become a potential target for cancer treatment, several challenges remain in the discovery and design of efficient and selective HK2 inhibitors due to its highly polar active site and complex protein functions.
PROTAC technology has been applied to target crucial oncoproteins involved in cancer cell proliferation, apoptosis, angiogenesis, immune evasion, inflammation, and cancer metastasis [13,14], while there are few cases of applying PROTAC technology to energy metabolism targets. Herein, we designed and synthesized a series of selective degradation agents targeting HK2 and conducted activity studies, expanding the applicability of PROTAC technology in the field of cancer and providing new options for the treatment of HK2-related diseases.
Firstly, to determine the reasonable BNBZ derivatization sites, SeeSAR12.1.0 and PoseView (
To evaluate the bioactivity of the synthesized compounds, HK2 enzyme inhibition assay and cytotoxicity test on SW480 cells were performed. All HK2 PROTACs exhibited weaker enzyme inhibitory activity than BNBZ (0.13 ± 0.03 µmol/L), due to the introduction of E3L-Linker, which increased the steric hindrance of the compounds. However, this structural modification has little effect on the enzyme-inhibitory activity of most compounds. Compared to BNBZ (10.67 ± 0.12 µmol/L), 4H-5P-M, 4H-5P-Th, 5H-1P-M, and 5H-5P-M showed higher cytotoxicity to SW480, with half maximal inhibitory concentration (IC50) values all less than 10 µmol/L (Table S1 and Fig. S1 in Supporting information). To expand the screening range of HK2 degradation activity, we defined the compound selection conditions, with IC50 (HK2 enzyme inhibition) < 5 µmol/L, and IC50 (SW480 cells) < 50 µmol/L.
Subsequently, we further investigated the degradation of active compounds to HK2 protein in SW480 cells. Western blot results showed that 4H-5P-M, 4H-6C-V1, 4H-3P-Th, 4H-5P-Th, and 5H-5P-M showed the HK2 degradation activity in SW480 cells, and the effect diminished with decreasing concentrations. Among them, the effect of 4H-5P-M was the most significant (Figs. S2A and S3 in Supporting information). The 50% protein degradation concentration (DC50) of 4H-5P-M inducing HK2 degradation in SW480 cells was 1.50 ± 0.15 µmol/L (Fig. S2A in Supporting information). In addition, we found that 4H-5P-M degraded HK2 in a time-dependent manner within 48 h in SW480 cells (Fig. S4 in Supporting information). In addition, 4H-5P-M induced the degradation of the HK2 interacting protein voltage dependent anion channel protein 1 (VDAC1) (Fig. S5 in Supporting information). Due to the excellent HK2 enzyme inhibitory activity, cytotoxic activity, and HK2 protein degradation activity of 4H-5P-M, we selected 4H-5P-M as a preferred HK2 degrader to further evaluate the anti-tumor effect and conduct the mechanism study.
We used 5-ethynyl-2′-deoxyuridine (EdU) staining to further determine the effect of 4H-5P-M on SW480 cell proliferation. It was found that the rate of proliferating cells treated with 4H-5P-M showed a significant dose-dependent decrease (Fig. S6 in Supporting information).
We employed the microscale thermophoresis (MST) to investigate the binding affinity of 4H-5P-M and hexokinases at molecular level [15]. The Kd value of 4H-5P-M with HK2 in buffer was 3.01 ± 0.64 µmol/L while 4H-5P-M had no significant binding affinity with HK1 or HK3 (Fig. S7 in Supporting information), indicating that it showed a better binding affinity with HK2 at the molecular level than HK1 or HK3. We performed a Western blot assay for the degradation activity of 4H-5P-M on four hexokinases [HK1, HK2, HK3, and glucokinase (GCK)] (Fig. S2B in Supporting information). The results showed that 0.1–25 µmol/L 4H-5P-M was able to degrade HK2 in a dose-dependent manner, while it displayed no significant degradation of HK1, HK3, or GCK. It suggested that 4H-5P-M was a specific HK2 degrader.
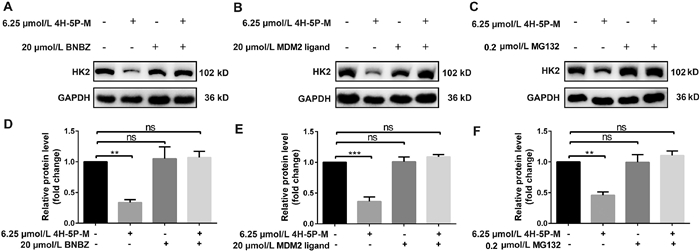
To confirm whether 4H-5P-M degrades HK2 via the PROTAC mechanism, we used the HK2 ligand BNBZ and the MDM2 ligand as competitive molecules to occupy the binding sites of HK2 or MDM2 proteins, respectively. The proteasome inhibitor MG132 was used as a tool to verify the action of the ubiquitin proteasome system in degrading target protein. It was found that SW480 cells treated with either BNBZ or MDM2 ligand could effectively resist the degradation of 4H-5P-M to HK2 (Figs. 2A, B, D and E). In addition, when MG132 was added to inhibit the activity of the proteasome, HK2 did not undergo degradation, indicating that 4H-5P-M induced the degradation of the target protein through the ubiquitin-proteasome system (Figs. 2C and F).
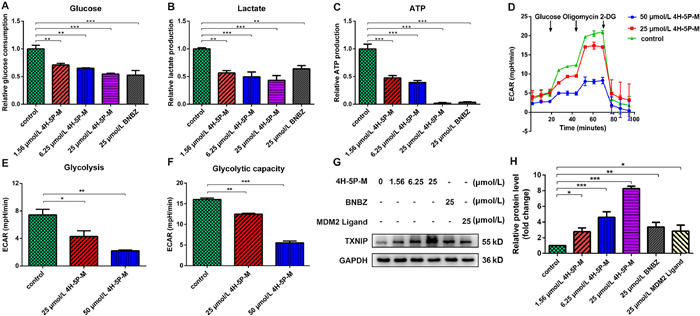
To investigate whether there is an effect on cancer cell glycolysis after 4H-5P-M degradation of HK2, we measured the glycolytic levels in SW480 cells after administration using the glucose, lactate, the ATP assay kits, and the seahorse XF glycolysis stress test kit. After 4H-5P-M or BNBZ treatment, the level of glucose uptake, lactate secretion, and ATP production decreased in SW480 cells (Figs. 3A–C). Corroborating this notion, the seahorse XF glycolysis stress assay measuring the extracellular acidification rate (ECAR), a proxy measure of glycolytic flux, showed that glycolysis and glycolytic capacity of SW480 cells reduced after 4H-5P-M treatment. 4H-5P-M downregulated the ECAR in SW480 cells (Figs. 3D–F). It was also found that 4H-5P-M more significantly upregulated intracellular thioredoxin-interacting protein (TXNIP) protein than BNBZ or MDM2 ligand, a recognized regulator of glucose metabolism that inhibits the expression of glycolytic enzymes associated with glucose metabolism (Figs. 3G and H). Collectively, these findings suggested that 4H-5P-M could reduce the glycolysis level in SW480 cells.
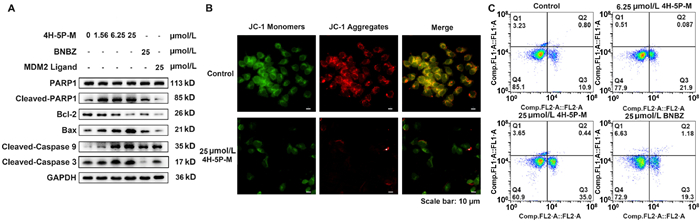
To identify whether there is an effect on HK2-mediated maintenance of mitochondrial membrane permeability and anti-apoptosis effect after degradation of HK2 by 4H-5P-M in SW480 cells, we analyzed the levels of apoptosis-related factors. The results showed that 4H-5P-M upregulated cleaved-PARP1, cleaved-caspase 9, and cleaved-caspase 3, and decreased intracellular Bcl-2/Bax levels more significantly than BNBZ or MDM2 ligand (Fig. 4A). It was also found that 4H-5P-M decreased cellular mitochondrial membrane potential (Fig. 4B) and increased the apoptosis proportion of cancer cells (Fig. 4C), indicating that 4H-5P-M induces apoptosis in SW480 cells.
We report a novel strategy for the selective degradation of HK2 by PROTAC technology to improve the efficacy of anti-tumor agents. We designed and synthesized 22 HK2 PROTACs. Among them, 4H-5P-M, the PROTAC with MDM2 ligand, was identified as the most active HK2 degrader, whose DC50 was 1.50 ± 0.15 µmol/L. MDM2, an E3 ubiquitin ligase, is also a key negative regulatory factor of the tumor suppressor p53 [16,17]. PROTACs based on MDM2 ligand not only degrade the target protein but also occupy the p53-MDM2 binding site blocking the interaction of p53 and MDM2, which results in a synergistic anti-tumor effect [18,19].
In addition, 4H-5P-M induced HK2 degradation in dose- and time-dependent manners. 4H-5P-M is bound to HK2 rather than HK1 or HK3, with a Kd value of 3.01 ± 0.64 µmol/L. 4H-5P-M, as a specific HK2 degrader, selectively degraded HK2 over other hexokinases. Moreover, it could effectively inhibit the enzymatic activity of HK2, cancer cell growth and glycolysis, and accelerate cancer cell apoptosis. By converting a HK2 inhibitor into a HK2 PROTAC, we developed a more effective HK2-targeting anti-tumor agent [20,21].
Meanwhile, the design and synthesis of dual-target PROTACs according to our study may be a good option [22]. Studies have shown that simultaneous inhibition of tumor energy metabolic pathways and other oncogenic signaling pathways can significantly improve the efficacy of anti-cancer drugs [5]. It is possible to overcome the resistance of oncology patients to inhibitors by combining selective HK2 PROTAC with other anti-tumor agents of therapeutic targets or with standard chemotherapy. Our study presented insights into the design and study of degraders targeting HK2. These findings may have important clinical implications.
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
The authors thank the Chunhui Program-Cooperative Research Project of the Ministry of Education, Liaoning Province Natural Science Foundation (No. 2022-MS-241), and Shenyang Young and Middle-aged Innovative Talents Support Program (No. RC210446) for financial supports. And we acknowledged the support from National-Local Joint Engineering Research Center for Molecular Biotechnology of Fujian & Taiwan TCM, Fujian Key Laboratory of Chinese Materia Medica, Fujian University Key Laboratory for Research and Development of TCM Resources, at Fujian University of Traditional Chinese Medicine.
Supplementary material associated with this article can be found, in the online version, at doi:
D. Hanahan, R. Weinberg, Cell 144 (2011) 646–674. doi: 10.1016/j.cell.2011.02.013
N. Pavlova, C. Thompson, Cell Metab. 23 (2016) 27–47. doi: 10.1016/j.cmet.2015.12.006
S.P. Mathupala, Y.H. Ko, P.L. Pedersen, Oncogene 25 (2006) 4777–4786. doi: 10.1038/sj.onc.1209603
A. Magrì, S. Reina, V. Pinto, Front. Chem. 6 (2018) 108. doi: 10.3389/fchem.2018.00108
F. Xu, J. Yan, Y. Gan, et al., Mol. Ther. Nucl. Acids 18 (2019) 308–319. doi: 10.1016/j.omtn.2019.09.002
G.Y. Gwak, J.H. Yoon, K.M. Kim, et al., J. Hepatol. 42 (2005) 358–364. doi: 10.1016/j.jhep.2004.11.020
G.L. Semenza, J. Clin. Investig. 123 (2013) 3664–3671. doi: 10.1172/JCI67230
K.C. Patra, Q. Wang, P.T. Bhaskar, et al., Cancer Cell 24 (2013) 213–228. doi: 10.1016/j.ccr.2013.06.014
D. Dewaal, V. Nogueira, A.R. Terry, et al., Nat. Commun. 9 (2018) 446. doi: 10.1038/s41467-017-02733-4
S.N. Garcia, R.C. Guedes, M.M. Marques, Curr. Med. Chem. 26 (2020) 7285–7322. doi: 10.2174/0929867326666181213092652
W. Li, M.Z. Zheng, S.P. Wu, et al., J. Exp. Clin. Cancer Res. 36 (2017) 58. doi: 10.1080/15567036.2016.1200166
M.Z. Zheng, C.R. Wu, K.Y. Yang, et al., Pharmacol. Res. 164 (2021) 105367. doi: 10.1016/j.phrs.2020.105367
A.C. Lai, C.M. Crews, Nat. Rev. Drug Discov. 16 (2017) 101–114. doi: 10.1038/nrd.2016.211
Y. Liu, M.Z. Zheng, Z.L. Ma, Chin. Chem. Lett. 34 (2023) 107762. doi: 10.1016/j.cclet.2022.107762
I. Churcher, J. Med. Chem. 61 (2018) 444–452. doi: 10.1021/acs.jmedchem.7b01272
X.Y. Li, W.C. Pu, Q.Q. Zheng, et al., Mol. Cancer 21 (2022) 99. doi: 10.1186/s12943-021-01434-3
L. Khavrutskii, J. Yeh, O. Timofeeva, et al., J. Vis. Exp. 78 (2013) 50541.
M.Y. Chang, F. Gao, J. Chen, et al., Acta Mater. Med. 1 (2022) 333–342.
Y.B. Li, J.L. Yang, A. Aguilar, et al., J. Med. Chem. 62 (2019) 448–466. doi: 10.1021/acs.jmedchem.8b00909
J. Hines, S. Lartigue, H.Q. Dong, et al., Cancer Res. 79 (2019) 251–262. doi: 10.1158/0008-5472.can-18-2918
Q. Zhao, T. Lan, S. Su, et al., Chem. Commun. 55 (2019) 369–372. doi: 10.1039/c8cc07813k
M.Z. Zheng, J.H. Huo, X.X. Gu, et al., J. Med. Chem. 64 (2021) 7839–7852. doi: 10.1021/acs.jmedchem.1c00649

Figure 1 The selection of HK2 ligand derivatization site and the synthesis route of representative HK2-PROTAC molecules.

Figure 2 Exploration of specific degradation pathways. (A, B, D, E) Effect of BNBZ and MDM2 ligand on HK2 expression in SW480 cells. (C, F) MG132 reverses the degradation effect of 4H-5P-M on HK2 in SW480 cells. The quantification of the relative HK2 protein content in the immunoblots is presented as mean ± standard deviation (SD) (n = 3) in the bar graph. Statistical significance was calculated with unpaired two-tailed Student's t-test. **P < 0.01; ***P < 0.001. ns means no significant difference. GAPDH, glyceraldehyde-3-phosphate dehydrogenase.

Figure 3 4H-5P-M inhibits glycolysis in SW480 cells. 4H-5P-M inhibits the glucose uptake (A), lactate secretion (B), and ATP production (C). 4H-5P-M were subjected to glycolysis stress assay in response to glucose, oligomycin, and 2-DG (D). The glycolysis (E) and the glycolytic capacity (F) of SW480 cells treated with different concentrations of 4H-5P-M was calculated from ECAR. The expression of TXNIP in SW480 cells was determined by Western blot after treating the cells with different concentrations of 4H-5P-M, BNBZ or MDM2 ligand for 48 h (G, H). The quantification is presented as mean±SD (n = 3) in the bar graph. Statistical significance was calculated with unpaired two-tailed Student's t-test. *P < 0.05; **P < 0.01; ***P < 0.001.

Figure 4 4H-5P-M induces apoptosis in SW480 cells. (A) The expression of apoptosis-related proteins in SW480 cells was determined by Western blot after treating cancer cells with different concentrations of 4H-5P-M, BNBZ or MDM2 ligand for 48 h. (B) Confocal imaging of cellular mitochondrial membrane potential of SW480 cells treated with different concentrations of 4H-5P-M for 48 h was determined. (C) Flow cytometry was used to analyze apoptosis of SW480 cells by Annexin V-FITC/PI staining.

 扫一扫看文章
扫一扫看文章

扫一扫关注我们
 DownLoad:
DownLoad:
 下载:
下载:
 下载:
下载: