School of Public Health, Research Center of Public Health, Renmin Hospital of Wuhan University, Wuhan University, Wuhan 430071, China
b.
Department of Radiation and Medical Oncology, Zhongnan Hospital of Wuhan University, Wuhan University, Wuhan 430071, China
c.
Key Laboratory of Magnetic Resonance in Biological Systems, State Key Laboratory of Magnetic Resonance and Atomic and Molecular Physics, National Center for Magnetic Resonance in Wuhan, Wuhan Institute of Physics and Mathematics, Innovation Academy for Precision Measurement Science and Technology, Chinese Academy of Sciences-Wuhan National Laboratory for Optoelectronics, Wuhan 430071, China
d.
University of Chinese Academy of Sciences, Beijing 101408, China
e.
Institute of Neuroscience and Brain Diseases; Xiangyang Central Hospital, Affiliated Hospital of Hubei University of Arts and Science, Xiangyang 441000, China
f.
Phase I Clinical Trial Laboratory, Zhongnan Hospital of Wuhan University, Wuhan 430071, China
bfyuan@whu.edu.cn (B.-F. Yuan). 1 These authors contributed equally to this work.
Received Date:
21 July 2023 Accepted Date:
26 October 2023 Revised Date:
06 October 2023 Available Online:
15 August 2024
Abstract:
Sleep deprivation (SD) is a widespread issue that disrupts the lives of millions of people. These effects initiate as changes within neurons, specifically at the DNA and RNA level, leading to disruptions in neuronal plasticity and the dysregulation of various cognitive functions, such as learning and memory. Nucleic acid epigenetic modifications that could regulate gene expression have been reported to play crucial roles in this process. However, there is a lack of comprehensive research on the correlation of SD with nucleic acid epigenetic modifications. In the current study, we aimed to systematically investigate the landscape of modifications in DNA as well as in small RNA molecules across multiple tissues, including the heart, liver, kidney, lung, hippocampus, and spleen, in response to chronic sleep deprivation (CSD). Using liquid chromatography-tandem mass spectrometry (LC-MS/MS) analysis, we characterized the dynamic changes in DNA and RNA modification profiles in different tissues of mice under CSD stress. Specifically, we observed a significant decrease in the level of 5-methylcytosine (5mC) and a significant increase in the level of 5-hydroxymethylcytosine (5hmC) in the kidney in CSD group. Regarding RNA modifications, we observed an overall increased trend for most of these significantly changed modifications across six tissues in CSD group. Our study sheds light on the significance of DNA and RNA modifications as crucial epigenetic markers in the context of CSD-induced stress.
Sleep is an essential physiological process and it occupies one-third of our lives, playing a fundamental role for physical, mental, and emotional health [1]. Experimental evidence has shown that sleep serves a restorative function by facilitating the clearance of metabolic waste products that accumulate in the brain during wakefulness [2]. However, sleep deprivation (SD) has become a concerning issue in modern societies [3]. Lifestyle habits, sleep disorders such as insomnia and obstructive sleep apnea, as well as neurological disorders, can contribute to insufficient sleep [3]. SD has been shown to have detrimental effects on cognitive function, particularly in tasks requiring sustained attention and working memory [4].
The prevalence of SD leading to excessive daytime sleepiness ranges from 9% to 24%, making it a common reason for visits to sleep clinics [5]. Long-term SD can lead to the development of physiological and neurobehavioral problems. It has been associated with an increased risk of diabetes, heart diseases, depression, and impulsive behaviors [6–8]. Inadequate sleep is widely recognized as a risk factor for various acute and chronic diseases, contributing to higher mortality and morbidity rates, especially in cardiovascular diseases [5]. In recent years, there has been a growing interest in sleep-related research [9,10]. SD has been found to be associated with increased oxidative stress, altered inflammatory responses, impaired synaptic plasticity, and a reduction in cell proliferation and neurogenesis [11,12]. However, the impact of SD on epigenetic modifications remains largely unexplored. Investigation into the effects of SD on epigenetic regulation is crucial for understanding its underlying mechanisms and potential implications for health.
Epigenetic modifications occurring in both DNA and RNA play significant roles in various physiological and pathological processes [13–16]. One well-known DNA modification is the addition of a methyl group to cytosine, resulting in 5-methylcytosine (5mC) [17–20]. 5mC is crucial in regulating gene expression [17]. The ten-eleven translocation (Tet) family can further convert 5mC to 5-hydroxymethylcytosine (5hmC), which also plays important roles in modulating gene expression [21–25]. In addition to DNA modifications, emerging studies have demonstrated the importance of RNA modifications in maintaining normal cellular functions [26]. RNA modifications are prevalent in RNA molecules [27–30], including transfer RNAs (tRNAs) [31,32], which typically range in size from 70 to 90 nucleotides [33]. In fact, tRNAs comprise the majority (~90%) of small RNA molecules [34]. Modifications in tRNAs are essential for preserving their characteristic clover-leaf-shaped structures and ensuring accurate translation of genetic information [31]. These modifications contribute to various aspects of tRNA function, such as intracellular localization, codon decoding fidelity, and structural stability [35]. Disruptions in the normal pattern of tRNA modifications can have profound effects on protein synthesis and cellular homeostasis [35]. Therefore, alterations in tRNA modifications have the potential to impact cell function and may be implicated in various diseases and pathological conditions.
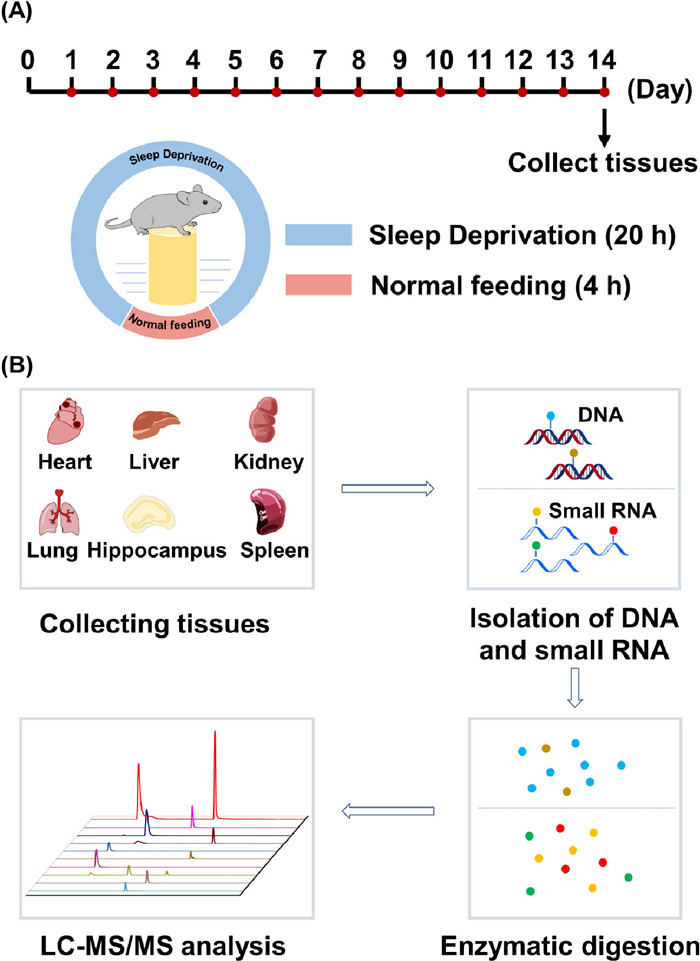
In the current study, we conducted a comprehensive investigation into the effects of chronic sleep deprivation (CSD) on DNA and RNA modifications in six major mouse tissues, including the heart, liver, kidney, lung, hippocampus, and spleen. Liquid chromatography-tandem mass spectrometry (LC-MS/MS) that is capable of sensitive detection of biological molecules [36–38] was employed to analyze these modifications. The CSD treatment and confirmation were carried out according to our previous study [39]. Typically, we used nine-week-old male C57BL/6J mice, which were group-housed in a specific pathogen-free (SPF) room maintained at a temperature of 23 ± 2 ℃ with a light-dark cycle of 8:00-20:00 and 20:00-8:00, respectively. The mice were randomly divided into two groups, with six mice in each group. The control group was allowed to sleep naturally, while the CSD group underwent 14 days of sleep deprivation. The sleep deprivation period for the CSD group was 20 h per day, from 2:00 pm to 10:00 am the next day, followed by a 4-h sleep opportunity from 10:00 am to 2:00 pm. On the 14th day, we sacrificed the mice and collected various tissues (heart, liver, kidney, lung, hippocampus, and spleen) for DNA and small RNA (< 200 nt) isolation according to the schematic illustration shown in Fig. 1A. Subsequently, both DNA and small RNA samples underwent enzymatic digestion and were subjected to LC-MS/MS analysis, as depicted in Fig. 1B. The isolated small RNAs were also assessed using polyacrylamide gel electrophoresis (Fig. S1 in Supporting information). The animal experiments were approved by the animal care and use committee in Innovation Academy for Precision Measurement Science and Technology, Chinese Academy of Sciences.
Figure 1
Figure 1.
Schematic illustration for CSD of mice and mass spectrometry profiling of DNA and RNA modifications. (A) The timeline for CSD treatment is depicted, showing the schedule for the CSD group with a 14-day period. (B) The workflow for the profiling of DNA and RNA modifications in different mouse tissues using LC-MS/MS analysis. The procedure involves the collection of tissue samples from control and CSD groups, followed by DNA and small RNA isolation. Enzymatic digestion is performed on the isolated samples, and LC-MS/MS analysis is conducted to detect and quantify DNA and RNA modifications.
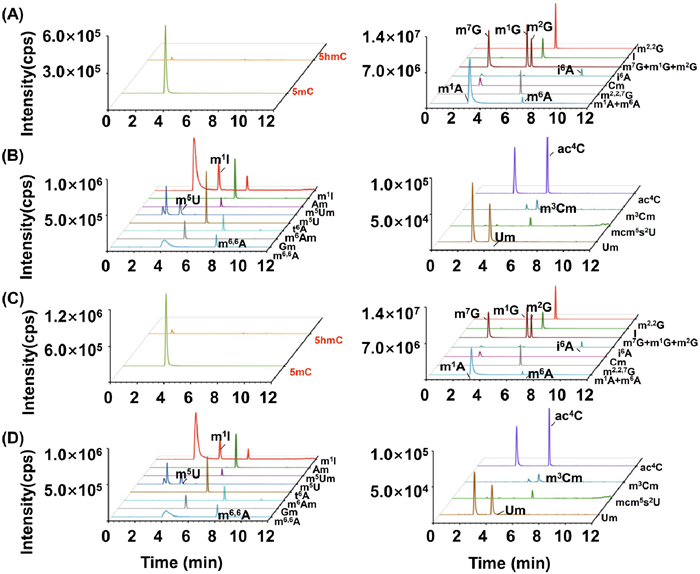
We first established a LC-MS/MS method to comprehensively analyze DNA and RNA modifications. Through optimization of the chromatographic separation conditions, we successfully achieved simultaneous detection of 47 RNA modifications and 2 DNA modifications (Figs. S2–S4, Tables S1 and S2 in Supporting information). With the LC-MS/MS method, we conducted an analysis of modifications in DNA and small RNA from various tissues obtained from both the control and CSD groups. Our findings revealed the presence of 2 DNA modifications (5mC and 5hmC) and 22 RNA modifications (Am, m6A, m1A, m6,6A, m6Am, t6A, i6A, I, m1I, Cm, m3Cm, ac4C, Gm, m1G, m2G, m7G, m2,2G, m2,2,7G, Um, mcm5s2U, m5U and m5Um) in all the examined tissues, including the heart, liver, kidney, lung, hippocampus, and spleen (Fig. 2 and Figs. S5-S13 in Supporting information).
Figure 2
Figure 2.
The representative extracted-ion chromatograms of 2 DNA modifications and 22 RNA modifications from mouse hippocampus. (A, B) Modifications detected in DNA and small RNA of mouse hippocampus from control group. (C, D) Modifications detected in DNA and small RNA of mouse hippocampus from CSD group.
We proceeded to construct calibration curves to accurately quantify the DNA and RNA modifications. The results demonstrated excellent linearity, with coefficients of determination (R2) exceeding 0.99. The limits of detection (LODs), defined as the minimum amounts of analytes required to achieve a signal-to-noise ratio of 3, ranged from 0.07 fmol to 26.8 fmol (Tables S3 and S4 in Supporting information). To assess the accuracy and precision of the method, we calculated the relative errors (REs) as well as the intra- and inter-day relative standard deviations (RSDs). The results indicated that REs were below 18.4% and RSDs were below 12.7% (Tables S5 and S6 in Supporting information), which demonstrated the good accuracy and precision of the method.
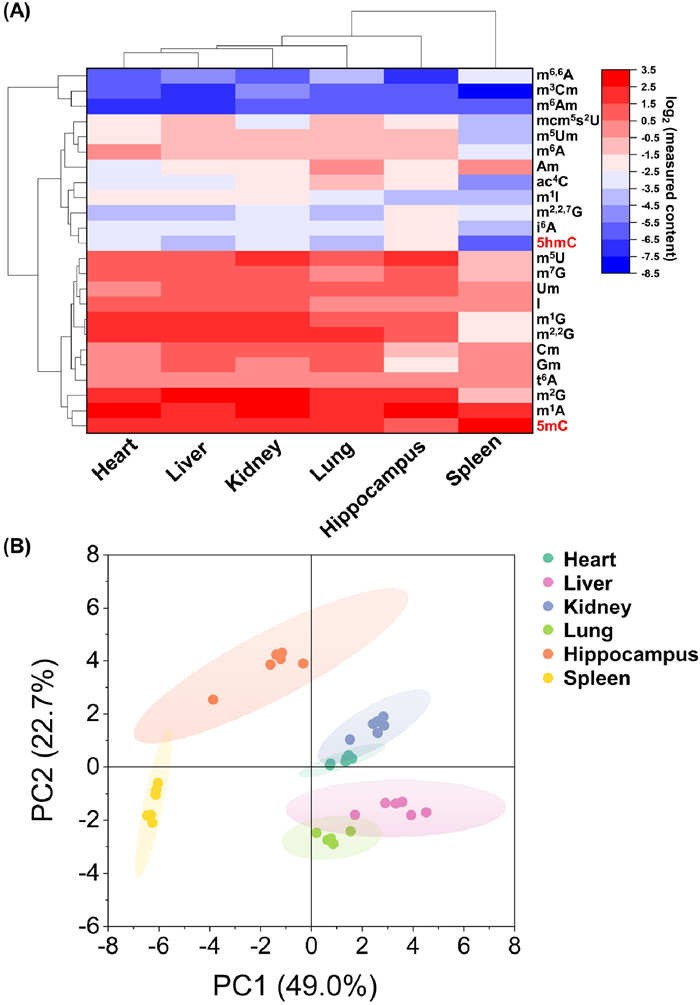
With the utilization of the constructed calibration curves, we successfully quantified 2 DNA modifications and 22 RNA modifications in mouse tissues (Figs. S14-S20 in Supporting information). The measured levels of RNA modifications were comparable to the previously reported levels in genomic DNA and small RNA of mammals (Table S7 in Supporting information). Modifications such as m6,6A, m3Cm, m6Am, mcm5s2U, m5Um, ac4C, m1I, m2,2,7G, and i6A, and 5hmC generally exhibited lower levels across different mouse tissues (Fig. 3A). On the other hand, modifications like 5mC, m1A, m2G, t6A, Gm, Cm, m2,2G, I, Um, m7G and m5U generally displayed higher levels across different mouse tissues (Fig. 3A). The cluster analysis revealed that the relative level of 5mC was comparable across different mouse tissues. However, 5hmC exhibited differential expression patterns among the tissues (Fig. 3A). As for RNA modifications, the signature profiles in the spleen exhibited distinct features compared to all other tissues, while the modifications in the heart showed similarities to those in the liver (Fig. 3A). To further analyze the data, we performed principal component analysis (PCA) on the measured RNA modifications. The results demonstrated that different tissues could be effectively grouped based on their RNA modification profiles, indicating that the overall levels of RNA modifications are tissue-specific (Fig. 3B).
Figure 3
Figure 3.
DNA and RNA modification landscapes in mouse multiple tissues. (A) The heatmaps showed the relative levels of DNA and RNA modifications across six mouse tissues. The average level of each modification from six mice (control group) was calculated and then subjected to log2 transformation for subsequent cluster analysis. (B) Principal component analysis (PCA) of RNA modifications in small RNA across six mouse tissues.
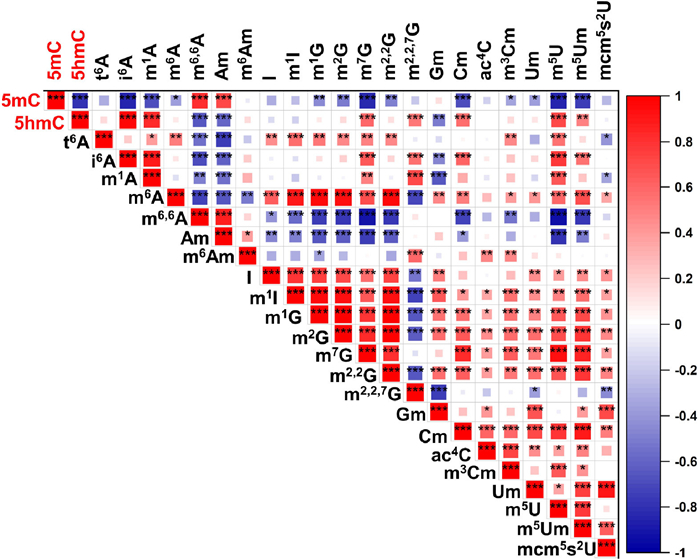
It is well established that various epigenetic marks play a coordinated role in regulating gene expression and cellular processes [40,41]. However, the correlation between DNA modifications and different RNA modifications has been relatively unexplored. In our study, we observed that 5mC displayed a general negative correlation with most RNA modifications (Fig. 4). On the other hand, many RNA modifications demonstrated positive correlations with other types of modifications, including t6A, i6A, m6A, I, m1I, m1G, m2G, m7G m2,2G, Cm, ac4C, m3Cm, Um, m5U, m5Um, and mcm5s2U. However, a small subset of RNA modifications exhibited negative correlations with other modifications, such as m6,6A, Am, and m2,2,7G (Fig. 4). These findings suggest that there may be prevalent coordinated regulation between DNA and RNA modifications. To maintain tissue-specific homeostasis, it is likely that a coordinated regulatory network exists to regulate RNA modifications. By identifying specific RNA modification patterns in different tissues and investigating the relationships between different RNA modifications, we can delve deeper into the complex regulatory functions and mechanisms that underlie tissue specificity. This opens up new avenues for further research in understanding the intricate interplay between DNA and RNA modifications.
Figure 4
Figure 4.
Correlation analysis between different DNA and RNA modifications across multiple tissues. *P < 0.05; **P < 0.01; ***P < 0.001. The level of each modification in multiple tissues from six mice (control group) was used for the correlation analysis. The numbers displayed in the right bar indicate the Pearson correlation coefficient.
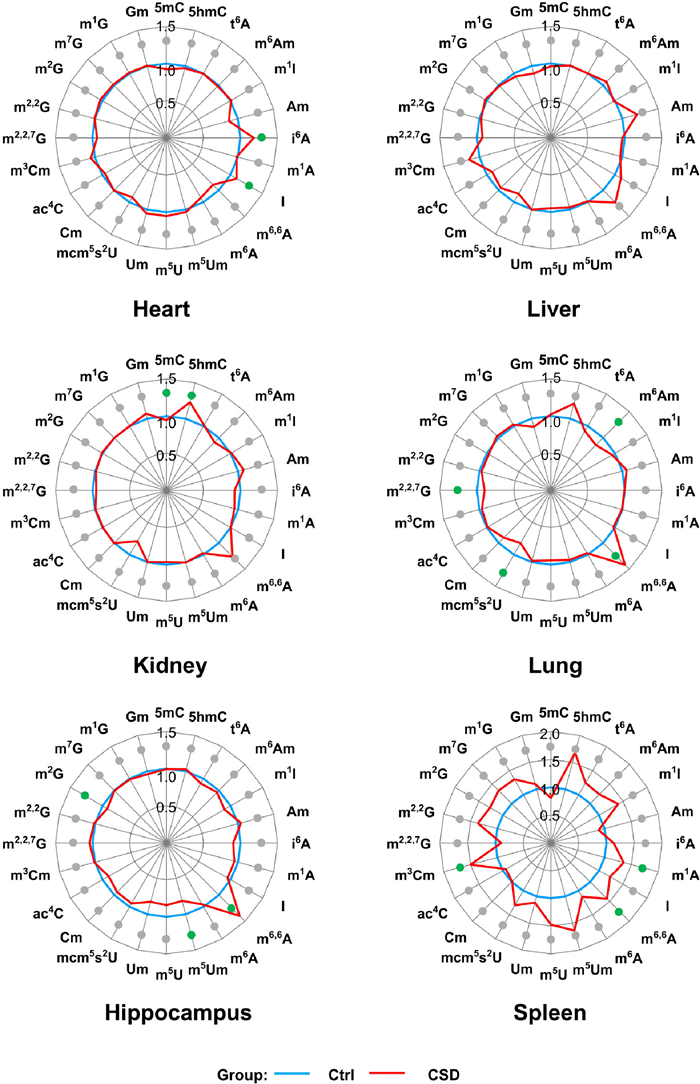
The quantification results revealed comparable levels of 5mC and 5hmC in different tissues, except in the kidney where significant changes were observed between the control and CSD groups (Fig. 5 and Fig. S14). Specifically, in the kidney, we observed a significant decrease in the level of 5mC and a significant increase in the level of 5hmC in the CSD group (Fig. 5 and Fig. S14). In eukaryotes, 5mC is commonly associated with the transcriptional repression of transposable elements, whereas 5hmC has been linked to marking actively transcribed genes [42]. It has been reported in previous studies that alterations in sleep duration can have detrimental effects on renal function [43]. The observed changes in the levels of 5mC and 5hmC in the DNA of the kidney from CSD mice may result in the dysregulation of specific gene expressions, ultimately contributing to the decline in renal function. However, the underlying mechanism behind the significant changes in the levels of 5mC and 5hmC, as well as the specific genes affected, requires further investigation.
Figure 5
Figure 5.
Relative alteration of the levels of DNA and RNA modifications in six mouse tissues in CSD group compared to control group. The average level of each RNA modification from six mouse tissues in CSD group was normalized to the control group. Dot in gray represents no significant difference; dot in green represents significant difference.
Regarding RNA modifications, we observed an overall increased trend for most of these modifications across six tissues in the CSD group compared to the control group (Fig. 5 and Figs. S15-S20). Specifically, the levels of i6A and I in heart, m6,6A in lung and hippocampus, m1A, m6,6A, and m3Cm in spleen, were significantly increased in CSD group compared to control group (Fig. 5). Additionally, we also observed significant decreases in certain modifications, such as m6Am, mcm5s2U, and m2,2,7G in lung, and m5Um and m2G in hippocampus from the CSD group compared to the control group (Fig. 5 and Figs. S15-S20). These findings demonstrate that CSD can impact RNA modifications in mouse tissues. However, it is important to note that the alterations in RNA modifications induced by CSD are tissue-specific (Fig. 5). This suggests that RNA modifications may play different roles in the context of CSD across various tissues. Further investigations are needed to elucidate the specific functions and mechanisms of these tissue-specific RNA modifications in the context of CSD.
It has been well-documented that RNA modifications are sensitive to environmental exposures, such as heat shock, hypoxia, and chemicals [44–50]. These modifications play an active role in facilitating adaptation to environmental challenges. However, the investigation of RNA modifications in animals subjected to CSD stress is relatively limited. In our current study, we aimed to systematically characterize the landscape of RNA modifications in small RNA molecules across multiple tissues, including heart, liver, kidney, lung, hippocampus, and spleen, in response to CSD stress. Our findings revealed dynamic changes in the RNA modification profiles in different tissues under CSD stress. These significantly altered modifications in small RNA molecules under CSD stress have the potential to impact translation rates and ultimately contribute to the development of various diseases. This study sheds light on the role of RNA modifications as novel epigenetic marks in the context of CSD stress and highlights their potential implications in disease pathogenesis. Apart from small RNAs, it is worth noting that other RNA species, including mRNA and rRNA, also contain numerous modifications [51–54]. Therefore, future studies to explore the correlation between CSD and modifications in these RNA species could provide a broader understanding of the impact of CSD on RNA modifications.
Recent studies have shown that SD has a significant impact on the consolidation of synaptic connections [55]. SD can lead to various molecular, genetic, and structural changes in neurons. However, the specific molecular changes associated with SD from an epigenetic perspective are still unclear. In our study, we focused on identifying tissue-specific tRNA modifications and their dynamic changes in mouse tissues under CSD stress. Although our current methods have limitations in pinpointing site-specific RNA modifications and characterizing alterations at single-base resolution under CSD stress, the use of sequencing technology to map tRNA modifications can provide a more comprehensive understanding of the landscape of alterations in tRNA modifications and the regulatory mechanisms involved in the response to CSD stress. Overall, our study offers new insights into the epigenetic mechanisms underlying CSD and paves the way for future research in this area.
In summary, we conducted a comprehensive analysis using LC-MS/MS to investigate the impact of CSD on DNA and RNA modifications in mouse tissues. Our findings revealed that CSD led to significant alterations in both DNA and RNA modifications across different mouse tissues. To the best of our knowledge, this is the first study to demonstrate a correlation between CSD and modifications in both DNA and RNA. These results underscore the potential influence of CSD on epigenetic DNA and RNA modifications. Future studies that delve into the intricate mechanisms underlying the correlation between CSD and epigenetic modifications on both DNA and RNA would offer valuable insights into the epigenetic implications of sleep disturbances.
Declaration of competing interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
The work is supported by the National Key R&D Program of China (Nos. 2022YFC3400700, 2022YFA0806600), the National Natural Science Foundation of China (Nos. 22277093, 22074110, 21721005), the Interdisciplinary Innovative Talents Foundation from Renmin Hospital of Wuhan University (No. JCRCGW-2022-008), the Wuhan Knowledge Innovation Project (No. 2022020801010111), and the Natural Science Foundation of Hubei Province (No. 2022CFB569).
Supplementary materials
Supplementary material associated with this article can be found, in the online version, at doi:10.1016/j.cclet.2023.109259.
J. Xiong, K.K. Chen, N.B. Xie, et al., Chin. Chem. Lett. 34 (2023) 108953.
[55]
F. Raven, E.A. Van der Zee, P. Meerlo, R. Havekes, Sleep Med. Rev. 39 (2018) 3–11. doi: 10.1016/j.smrv.2017.05.002
Figure 1
Schematic illustration for CSD of mice and mass spectrometry profiling of DNA and RNA modifications. (A) The timeline for CSD treatment is depicted, showing the schedule for the CSD group with a 14-day period. (B) The workflow for the profiling of DNA and RNA modifications in different mouse tissues using LC-MS/MS analysis. The procedure involves the collection of tissue samples from control and CSD groups, followed by DNA and small RNA isolation. Enzymatic digestion is performed on the isolated samples, and LC-MS/MS analysis is conducted to detect and quantify DNA and RNA modifications.
Figure 2
The representative extracted-ion chromatograms of 2 DNA modifications and 22 RNA modifications from mouse hippocampus. (A, B) Modifications detected in DNA and small RNA of mouse hippocampus from control group. (C, D) Modifications detected in DNA and small RNA of mouse hippocampus from CSD group.
Figure 3
DNA and RNA modification landscapes in mouse multiple tissues. (A) The heatmaps showed the relative levels of DNA and RNA modifications across six mouse tissues. The average level of each modification from six mice (control group) was calculated and then subjected to log2 transformation for subsequent cluster analysis. (B) Principal component analysis (PCA) of RNA modifications in small RNA across six mouse tissues.
Figure 4
Correlation analysis between different DNA and RNA modifications across multiple tissues. *P < 0.05; **P < 0.01; ***P < 0.001. The level of each modification in multiple tissues from six mice (control group) was used for the correlation analysis. The numbers displayed in the right bar indicate the Pearson correlation coefficient.
Figure 5
Relative alteration of the levels of DNA and RNA modifications in six mouse tissues in CSD group compared to control group. The average level of each RNA modification from six mouse tissues in CSD group was normalized to the control group. Dot in gray represents no significant difference; dot in green represents significant difference.

 DownLoad:
DownLoad:

 下载:
下载:






 下载:
下载:
