Enzymatic pretreatment mitigates the dissemination of antibiotic resistance genes via regulating microbial populations and gene expressions during food waste fermentation
Citation:
Yang Wu, Wanying Hu, Haining Huang, Xiong Zheng, Lei Dong, Yinguang Chen. Enzymatic pretreatment mitigates the dissemination of antibiotic resistance genes via regulating microbial populations and gene expressions during food waste fermentation[J]. Chinese Chemical Letters,
2023, 34(8): 108058.
doi:
10.1016/j.cclet.2022.108058
Enzymatic pretreatment mitigates the dissemination of antibiotic resistance genes via regulating microbial populations and gene expressions during food waste fermentation
English
Enzymatic pretreatment mitigates the dissemination of antibiotic resistance genes via regulating microbial populations and gene expressions during food waste fermentation
State Key Laboratory of Pollution Control and Resource Reuse, School of Environmental Science and Engineering, Tongji University, Shanghai 200092, China
Received Date:
27 May 2022 Accepted Date:
10 December 2022 Revised Date:
21 November 2022 Available Online:
15 August 2023
Abstract:
Food waste (FW) has been recognized as essential reservoir for resource recovery via anaerobic fermentation, which could also bring the potential risk of antibiotic resistance genes (ARGs) dissemination. Although the structural deficiency of FW could be stimulated by enzymatic pretreatment to enhance fermentation efficiency, the influences of enzymatic pretreatment on ARGs fate and microbial metabolic pathways involved in ARGs dissemination have rarely been reported. This work proved that enzymatic pretreatment could effectively decrease the total abundance of ARGs (reduced by 13.8%-24.5%) during long-term FW fermentation. It was found that enzymatic pretreatment significantly reduced the ARGs belonging to the efflux pump, which might be ascribed to its ability to increase membrane permeability. Furthermore, enzymatic pretreatment was in favor of reducing microbial diversity and various potential ARGs host (e.g., Methanosarcina, Clostridium, Prevotella, Parabacteroides). Also, this pretreatment remarkably up-regulated the genetic expressions involved in ABC transporter (e.g., eryF and mntA) and down-regulated the genetic expressions that participated in DNA replication, two-component systems (e.g., uphA and cckA), and quorum sensing (e.g., rpfF and lsrG), thereby decreasing ARGs transmission. This study would expand the insight of the influences of pretreatment method on ARGs fate during FW fermentation, and offer practical guidance on the sustainable management of FW.
The wide utilization of antibiotics in daily life would result in the potential discharge to the environment, and subsequently cause the dissemination of antibiotic resistance genes (ARGs), one of the most important emerging contaminants with civil attention [1]. Notably, similar to sewage sludge and manure, food waste (FW) had gradually been recognized as the essential repository of ARGs [2], which might bring severe potential risks. Previous study had demonstrated that the relative abundance of ARGs in FW could reach 0.28–0.45 gene copies/16S rRNA copies and quickly transfer in the nearby environment [3]. Meanwhile, with fast advancement of industrialization and urbanization, the growth rate of FW production increased year by year, even over 10% in developing countries [4]. Hence, FW could pose the burdensome disposal problem, and the presence of ARGs in FW might aggravate the risk to human health and ecological safety.
Currently, anaerobic fermentation has been considered as a green and practical approach to treat FW for valorizing FW into high-value products (e.g., volatile fatty acids (VFAs)) [5]. The global VFAs demand was 18.5 million tonnes in 2020 with a 3% growth rate, and the market price of VFAs is estimated to be approximately 2000 €/tonnes currently [6,7]. Meanwhile, anaerobic fermentation has also been demonstrated to be able to effectively reduce ARGs dissemination, which would contribute to decrease the ecological risks [8]. For example, Tang et al. had proved that the targeted ARGs could be reduced 0.42–1.36 log units during the fermentation system [9]. Nevertheless, the organic substrates in FW were encapsulated by complex polymers (e.g., celluloses and proteins) [10], which had become the rate-limiting step for FW fermentation. Therefore, various pretreatment methods (e.g., ultrasonic, microwave, hydrothermal, enzyme) were conducted to enhance the efficiency of solubilization and hydrolysis. Compared with other methods, enzymatic pretreatment exhibited the advantages of high selectivity and low energy consumption [11]. However, to our best knowledge, the influence of enzymatic pretreatment on ARGs dissemination during long-term FW fermentation had been rarely reported, which required further explorations.
Moreover, the propagation of ARGs was dependent on the microorganisms, especially for ARGs hosts. For instance, Wang et al. had proved that the presence of graphene could greatly decrease the ARGs, including ermF, ermB, tetQ and tetX via reducing the abundance of Euryarchaeota, Thermotogae, Bacteroidetes, and Chloroflexi [12]. Hence, to comprehensively reveal the impact of enzymatic pretreatment on ARGs transmission during long-term fermentation, it was of great importance to track the dynamic variations of key anaerobe in the fermentation system, and further determine their specific role in ARGs proliferation. In addition, the achievement of microbially relevant functions depended on the expression of key genes, which also played an essential role in ARGs propagation. For example, Luo et al. had demonstrated the addition of surfactant up-regulates the genetic expressions of ABC transporter, resulting in the remarkable reduction (24%−46%) of total ARGs abundance [13]. However, the impact of enzymatic pretreatment on the critical metabolic pathways involved in ARGs dissemination had not yet been systematically illustrated.
The aim of this study is to unveil the influence of enzymatic pretreatment on ARGs transmission during long-term FW fermentation as well as the underlying mechanisms. Firstly, the variation of diverse ARGs and resistance mechanisms in reactors with different enzymatic pretreatment were determined. Then, the shifts of functional microbial profiles (including the potential ARGs hosts and genetic traits) responsible for ARGs dissemination are unveiled. Besides, the correlation of ARGs, microbial community, and metabolic pathways were characterized to further reveal the response mechanisms to enzymatic pretreatment. This study would expand the insight of pretreatment method on ARGs fate during FW fermentation process, and offer practical navigation on the sustainable management of FW.
The FW utilized in this study was obtained from the renewable energy utilization center in Shanghai, China. The oil and large undesirable particles had been firstly eliminated, and then stored at 4 ℃ for long-term experiments. The total and volatile suspended solids were 100.1 and 76.3 g/L, and the detailed information was exhibited in Table S1 (Supporting information). Meanwhile, waste activated sludge (WAS) was utilized as the inoculum, and it was obtained from a biological sewage treatment plant located in Shanghai, China. The obtained WAS was filtered to remove large particles, and stored at 4 ℃ before the experiments.
Since proteins and cellulose encased the FW in the form of stable biopolymer networks [14], the Viscozyme L as a representative of cellulase, and Flavourzyme as a representative of protease, were selected in this experiments and obtained from the Novozymes Biotechnology Co., Ltd. (Copenhagen, Denmark). The FW was firstly conducted by enzymatic pretreatment (cellulose with 1000 U/g TSS FW and protease with 10 LAPU/g TSS FW) at pH 4.8 and 50 ℃ for 12 h. The unit (U) represented the amount of enzyme required to hydrolyze cellulose to produce 1 mg glucose per hour, while LAPU (leucine aminopeptidase unit) represented the amount of enzyme required to hydrolyze 1 mmol of l-leucine-p-nitroaniline per minute. The optimal contents of cellulose and protease were based on the preliminary experiments [15].
After that, the liquid phase of pre-treated FW was obtained and added with Milli-Q water to the identical serum bottles (550 mL working volume) to maintain the initial total solid and fermentation volume as 60 g/L and 270 mL, respectively. Then, 30 mL of the concentrated WAS was dosed as the inoculum. The reactor with no enzymatic pretreatment and no fermentation pH control was selected as the control group (R0), while the reactor with enzymatic pretreatment and no fermentation pH control was named as R1. Meanwhile, since the pH during FW fermentation would be spontaneously decreased due to the generation of volatile fatty acids, the reactor with enzymatic pretreatment and fermentation pH regulation (pH 6) was set as R2 reactor. All the reactors were purged with high-purity (99.9%) N2 gas to maintain the anaerobic environment for 10 min. After being sealed with the rubber stoppers, all the bottles were placed in an air-bath shaker at 200 rpm and 35 ℃ for 48 d fermentation experiments. Moreover, the sludge retention time (SRT) was controlled for 6 d. Every 2 d, 100 mL of fermentation mixtures from different reactors were extracted, and 100 mL mixtures of FW, Milli-Q water, and WAS with the same proportions were correspondingly added. Then, all the reactors were purged with N2 and replaced in the air-bath shaker. The main product of FW fermentation is VFAs, including acetate, propionate, butyrate, and valerate. Among them, butyrate was the most abundant VFAs (36%−53%), followed by acetate (15%−28%). After 48 d experiments, the samples were extracted from the different reactors, and stored at −80 ℃ immediately for microbial analysis.
The TSS and VSS were detected referring to the standard method [16]. Meanwhile, the pH was detected by the pH meter (6010 M, Jenco, USA). Also, redundancy analysis (RDA) was applied by Canoco 5.0 software to disclose the correlation between ARGs, microbial populations, and metabolic pathways in response to enzymatic pretreatment. Moreover, the metagenomic sequencing was conducted via Illunima novaseq 6000 sequencing. The microbial DNA was extracted using the EZNA stool DNA Kit (Omega Bio-tek, Norcross, GA, USA). In briefly, 1 µg of genomic DNA was sheared by Covaris S220 Focused-ultrasonicator (Woburn, MA USA), and the sequencing libraries were prepared with a fragment length of approximately 450 bp. The taxonomy was assigned on customized kraken database, which included all genome sequences in NCBI RefSeq database. The microbial metabolic pathways and the corresponding genetic expressions were achieved by using the pathways mapped by genes annotated in the KEGG pathway database. The ARGs-OAP online pipeline for fast annotation and classification of ARG-like sequences from metagenomic data was used [17].
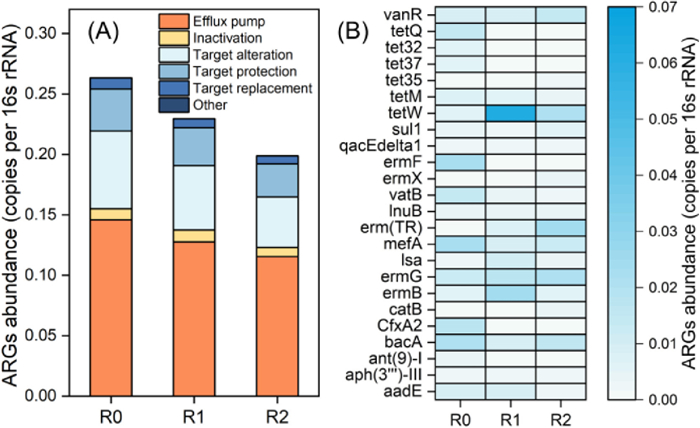
Enzymatic pretreatment was an efficient way to promote the depolymerization of FW and enhance the efficiency of solubilization and hydrolysis, which was beneficial for the VFAs production during anaerobic fermentation [11]. However, the effects of enzymatic pretreatment on the ARGs variation, which could bring potential ecological risk, had been rarely investigated. As exhibited in Fig. 1A, the total ARGs abundance in the reactor with no enzymatic pretreatment and no pH control (R0) was 0.2633 copies/16s rRNA. Nevertheless, it was reduced to 0.2296 copies/16s rRNA after enzymatic pretreatment (R1), and further decreased to 0.1987 copies/16s rRNA in the reactor with enzymatic pretreatment and acidic-pH operation (R2), which was approximately 24.5% remove efficiency. Obviously, the enzymatic pretreatment was favor to reduce the total ARGs during long-term FW fermentation, and the acidic-pH operation could further improve the promoting effect.
Figure 1
Figure 1.
Variation of total ARGs and resistance mechanisms (A) and specific ARGs (B) in different reactors during long-term FW fermentation with no enzymatic pretreatment and no pH control (R0), with enzymatic pretreatment and no pH control (R1), and with enzymatic pretreatment and acidic-pH operation (R2).
In terms of specific ARGs, the enzymatic pretreatment also exhibited distinct impacts. As depicted in Fig. 1B, the predominant ARGs, including mefA, ermF, bacA, CfxA2, vatB, tetQ, and tetW, etc., which were commonly determined in the FW [18]. Most of these ARGs were also observed to be decreased after enzymatic pretreatment, and further reduced with the fermentation pH control, except for tetW. For example, the abundance of mefA belonging to the MLS type reduced from 0.0222 copies/16s rRNA in R0 reactor to 0.01195 copies/16s rRNA in R1 reactor, and further reduced to 0.0099 copies/16s rRNA in R2 reactor. Meanwhile, in comparison with R0, the abundance of bacA belonging to the bacitracin type was decreased by 24.1 and 53.2% in R1 and R2 reactors, respectively. Moreover, the abundance of ermF (MLS type) and CfxA2 (beta-lactam type) was even reduced from 0.0218 to 0.0177 copies/16s rRNA in R0 to undetectable in both R2 and R3 reactor.
Moreover, further analysis demonstrated that the enzymatic pretreatment also exhibited the distinct impact on resistance mechanisms. As exhibited in Fig. 1A, the predominant resistance mechanism category of ARGs in fermentation systems was the efflux pump, which could remove the toxic substance and contribute to antibiotic resistance [19]. Its abundance was observed to be significantly decreased in the reactor with enzymatic pretreatment, especially with simultaneous pH adjustment (0.1458, 0.1276, and 0.1155 copies/16s rRNA in R0, R1, and R2 reactor). These results were also consistent with the variation of ARGs (e.g., mefA and ermF) belonging to this resistance mechanism. The previous publication had reported that protease was able to degrade the membrane protein to cell increase membrane permeability [20], which would be beneficial to reduce the ARGs affiliated to the efflux pump [13]. Generally, the enzymatic pretreatment was conducive to remove the ARGs during long-term fermentation, and the promoting impacts could be aggravated with the fermentation pH-regulation.
The microbial population had been accepted as the vital driver of ARGs proliferation [21]. Hence, the variation of microbial structure during long-term FW fermentation with different pretreatment was determined. The Shannon index related to the R0, R1, and R2 reactors were 4.20, 2.93, and 2.83, respectively. Thus, the decrement of microbial diversity induced by enzymatic pretreatment might be beneficial to enriching functional anaerobe, including the potential ARGs hosts. Further analysis found that Firmicutes (31.9%−42.4%), Bacteroidetes (1.51%−18.4%), Proteobacteria (10.19%−12.09%), and Actinobacteria (4.40%−33.4%) were the major phyla in the FW fermentation system (Fig. 2A), and were also frequently considered as the common ARGs hosts [22]. Nevertheless, their abundances were shifted remarkably under different pretreatment. For example, the relative abundance of Firmicutes was 42.4% in the reactor with no enzymatic pretreatment and no pH control (R0). Whereas, it was reduced to 34.8% in the reactor after enzymatic pretreatment (R1), and further decreased to 31.9% in the reactor with enzymatic pretreatment and acidic-pH operation (R2). Firmicutes had been considered as the essential microorganisms of hosts for MLS (mefA) and beta-lactam (CfxA2) [23]. Meanwhile, the relative abundance of Bacteroidetes, which were related to the proliferation of ARGs encoding ermF and tetQ [24], was 18.4%, 3.37%, and 1.51% in the R0, R1, and R2 reactors, respectively. Undoubtfully, their decrement in the reactor with enzymatic pretreatment, especially synchronous operation with fermentation pH regulation, contributed to the total ARGs reduction.
Figure 2
Figure 2.
Variations of microorganisms at phylum (A) and genus level (B) in different reactors during long-term FW fermentation with no enzymatic pretreatment and no pH control (R0), with enzymatic pretreatment and no pH control (R1), and with enzymatic pretreatment and acidic-pH operation (R2), and the Network analysis of the co-occurrence between ARGs and microorganisms (C).
In terms of the genus level (Fig. 2B), the main microorganisms in control reactor included Clostridiales sp. (7.80%), Methanosarcina (6.43%), Clostridium (5.75%), Clostridiaceae sp. (4.02%), Prevotella (3.06%), Parabacteroides (3.24%), and Bacteroides (2.87%), etc., which were frequently identified as the potential host of multiple antibiotic resistance types (especially MLS, bacitracin and beta-lactam) [25,26]. For instance, the relative abundance of Methanosarcina, which was characterized as the host microorganism of mefA and tetQ [12], was decreased from 6.43% in R0 reactor to 0.60% in R1 reactor, and further decreased to 0.38% in R2 reactor. Meanwhile, Clostridium genera were able to carry the various ARGs, especially aminoglycoside resistance genes [27], and its abundance was 5.75%, 1.25% and 1.05% in R0, R1, and R2 reactors, respectively. Moreover, the variation of Prevotella and Parabacteroides, which were also recognized as the potential ARGs hosts to deliver genes of ermF, vatB, and tetM [28,29], also exhibited the similar trend. In comparsion with R0 reactor, the relative abundance of Prevotella and Parabacteroides were reduced by 99.1% and 99.8% in R1 reactor while negligible in R2 reactor. Besides, Ruminococcaceae sp. was precited as the carrier of tetW [30], and its abundance was observed to be significantly enriched in the reactors with enzymatic pretreatment (0.40% in R0, 15.9% in R1, and 2.78% in R2). These results could also explain the remarkable elevation of tetW gene in R1 and R2 reactors.
Besides, the co-occurrence network analysis had been conducted to identify the potential ARGs hosts in fermentation systems. As demonstrated in Fig. 2C, mefA and tetQ were both positively correlated with Methanosarcina (P = 0.032 and 0.040) and Clostridiales sp. (P = 0.043 and 0.029), indicating these genera might be the potential host for mefA and tetQ, which exhibited agreement with the previous work [12]. Meanwhile, ermF and vatB were also observed to be positively linked with Parabacteroides (P = 0.001 and P = 0.022) and Prevotella (P = 0.004 and 0.020), which were also consistent with other study [28,29]. Moreover, CfxA2 was found to be positively related with Clostridium (P = 0.024), while bacA was positively correlated with Bacteria sp. (P = 0.004). Thus, the reduction of these microorganisms in the reactor with enzymatic pretreatment would promote the decrement of related ARGs. Generally, the shifts of microbial structure induced by enzymatic pretreatment, especially simultaneous acidic-pH operation, were favor to reduce most of ARGs during long-term FW fermentation.
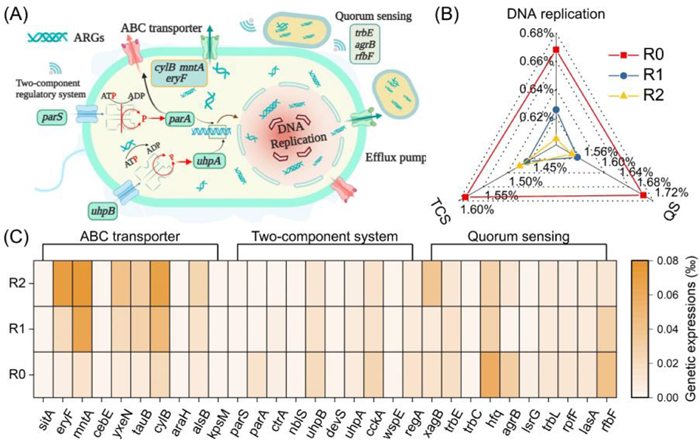
Apart from microbial population, the spread of resistance genes also relied on the regulation of microbial metabolism and the corresponding genetic expressions (Fig. 3A). Various metabolic pathways (e.g., DNA replication, ABC transporter, two-component regulatory system (TCS), and quorum sensing (QS)) had been recognized to be associated with microbial resistance to antibiotics [31,32]. Hence, the essential metabolic pathways responsible for ARGs transmission were annotated based on the KEGG databases. As depicted in Fig. 3B, the relative abundance of DNA replication, which participated in the generation and transfer of ARGs [19], was 0.67% in the reactor with no enzymatic pretreatment and no pH regulation (R0). However, after enzymatic pretreatment, it was reduced to 0.62% in the R1 reactor, and further decreased to 0.60% in the reactor with enzymatic pretreatment and acid-pH operation (R2). Hence, it could be hypothesized that the degradation of DNA replication induced by enzymatic pretreatment contributed to the reduced expression of resistance genes, resulting in decreasing their transmission during long-term FW fermentation.
Figure 3
Figure 3.
Schematic diagram of the ARGs transmission route (A), variation of critical metabolic pathways (B) and the corresponding genetic expressions (C) in different reactors during long-term FW fermentation with no enzymatic pretreatment and no pH control (R0), with enzymatic pretreatment and no pH control (R1), and with enzymatic pretreatment and acidic-pH operation (R2).
Meanwhile, the ABC transporter is able to discharge unfavorable substances (e.g., bacteriocins, antibiotics, and toxins) to the extracellular [33], and it could be considered as an indicator of membrane transfer efficiency [34], which was positively correlated with the membrane permeability. Its abundance increased from 2.73% in the R0 reactor to 2.90% and 2.98% in the R1 and R2 reactors, respectively (Fig. 3C). Correspondingly, most of the genes (e.g., eryF, mntA, yxen, tauB) that participated in ABC transporter was found to be significantly improved in the reactor with enzymatic pretreatment (31.0%−389.9%), especially simultaneous acid-pH operation (40.8%−980.8%). Hence, it could be inferred that the membrane permeability was enhanced by pH regulation. The over-expressed ABC transporter system induced by enzymatic pretreatment could promote the extracellular transfer of toxic substances, which was also consistent with the decrement of ARGs belonging to the efflux pump resistance mechanism.
Moreover, TCS can deliver cellular signals through the cytoplasm to the bacterial nucleolus and then regulate the corresponding genetic expression, leading to the mediation of drug resistance [35]. The relative abundance of TCS was 1.59% in R0 reactor, while it was reduced to 1.46% and 1.47% in R1 and R2 reactors, separately. Particularly, the essential genes that participated in TCS were observed to be down-regulated in the reactor with enzymatic pretreatment, especially with the simultaneous fermentation pH adjustment. For instance, compared with R0, the relative abundances of uphA and uphB, which induced bacterial resistance to various antibiotics through direct regulation of the efflux pump [36], were reduced by 21.4% and 19.3% in the R1 reactor, and further decreased by 33.7% and 37.2% in the R2 reactor, separately. Meanwhile, the relative abundance of cckA, which was component of the regulatory phosphorelay system and responsible for the cell growth, division, and intracellular survival [37], was 0.027‰, 0.016‰, and 0.015‰ in R0, R1, and R2 reactors, separately. Besides, the gene of crtA, wspE, and regA, which were related to antibiotic resistance transmission [13], also exhibited the similar trend. Obviously, the general reduction of genetic expressions related to TCS might be favor to decreasing the ARGs dissemination.
In addition, the QS system is a cell-to-cell communication mechanism, which relies on the production, detection and response of cellular signaling molecules. It could coordinate various cellular processes including plasmid transfer, genetic expressions, and antibiotic resistance, thereby controlling the ARG propagation process [38]. The relative abundance of QS was reduced from 1.69% in R0 reactor to 1.55% and 1.54% in R1 and R2 reactors, separately. Correspondingly, the variation of critical genetic expressions involved in QS system also exhibited the similar pattern as expected. For instance, the genetic expressions of rpfF, which was responsible for the intra or inter-species propagation of ARGs [39], was reduced by 48.6% and 51.7% in the R1 and R2 reactors, respectively. Meanwhile, the relative abundance of lsrG encoding (4S)−4‑hydroxy-5-phosphonooxypentane-2, 3‑dione isomerase, which could regulate the degradation of phospho-AI-2, was also decreased by 58.5% and 71.7% in the R1 and R2 reactors, separately. The previous publications demonstrated that the ARGs abundance was positively correlated with the expressions of QS degraded genes [39], which also exhibited agreement with this work. Besides, other genes involved in QS system (e.g., argB, lasA, trbC, trbL) were also observed to be down-regulated in the reactors with enzymatic pretreatment. Overall, enzymatic pretreatment, especially synchronous fermentation pH adjustment, promoted the up-regulation of ABC transporter and down-regulation of DNA replication, TCS, and QS, resulting in the reduction of ARGs during long-term FW fermentation.
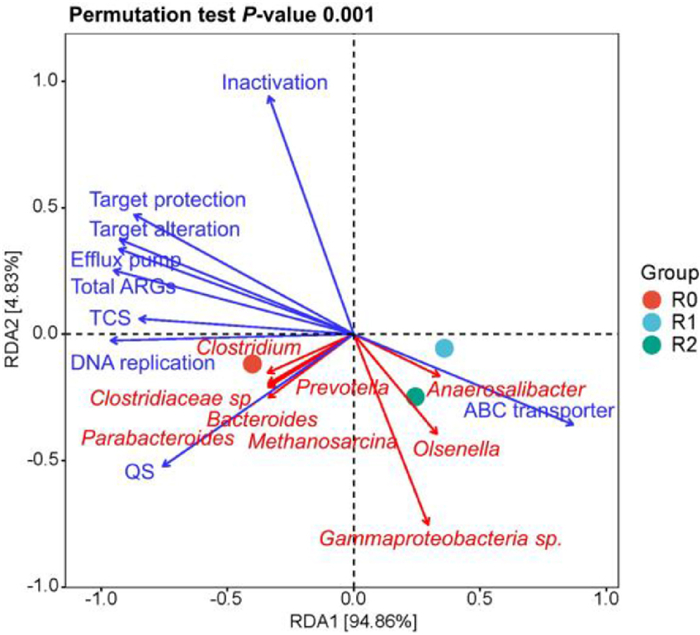
The previous publication reported that the ARGs dissemination was determined by various parameters (e.g., microorganisms and metabolic pathways) [13]. Hence, in this work, the RDA analysis was conducted to further disclose the correlations of ARGs, microbial community, and metabolic pathways in response to enzymatic pretreatment. As depicted in Fig. 4, the efflux pump was positively correlated with total ARGs abundance while ABC transporter with negatively linked with it, indicating the ABC transporter system accelerated the reduction of ARGs, especially ARGs belonging to the efflux pump, which was also exhibited agreement with the previous work [13]. Meanwhile, the ABC transporter also exhibited positive correlation with R1 and R2 reactor, also demonstrating that enzymatic pretreatment could effectively up-regulate the genetic expressions involved in ABC transporter, which might be ascribed to the improvement of membrane permeability by enzymatic pretreatment [40]. Moreover, the metabolic pathways (e.g., DNA replication, TCS, and QS) were also positively linked with the ARGs abundance. Thus, their down-regulations in the reactor with enzymatic pretreatment were favor to induce the low microbial kinetics of functional anaerobe for ARGs dissemination. Besides, various microorganisms including Bacteroides, Clostridium, Clostridiaceae sp., Methanosarcina, Prevotella, and Parabacteroides were found to be positively correlated with total ARGs abundance. Herein, the remarkable decrement of these ARGs hosts in the reactor with enzymatic pretreatment was responsible for the efficient reduction of ARGs during long-term FW fermentation.
Figure 4
Figure 4.
RDA analysis of the correlations of ARGs, microbial community, and metabolic pathways in different reactors during long-term FW fermentation with no enzymatic pretreatment and no pH control (R0), with enzymatic pretreatment and no pH control (R1), and with enzymatic pretreatment and acidic-pH operation (R2).
This work revealed that enzymatic pretreatment was beneficial to reduce the ARGs dissemination during long-term anaerobic fermentation of FW, and the promoting effect could be aggravated with the synchronous fermentation pH regulation. Enzymatic pretreatment significantly decreased the ARGs affiliated with the efflux pump, which might be ascribed to its ability to increase membrane permeability. Meanwhile, the microbial diversity and various potential ARGs host was observed to be reduced in response to enzymatic pretreatment. Moreover, metabolic pathways involved in ARGs proliferation was remarkably stimulated by enzymatic pretreatment, including the up-regulation the ABC transporter system and the down-regulation of DNA replication, TCS, and QS, resulting in the reduction of ARGs transmission. These findings provided new insights into the influences of pretreatment methods on ARGs fate during the fermentation process.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
This work was financially supported by the National Key Research and Development Program of China (No. 2019YFC1906302), the Shanghai Natural Science Foundation (No. 22ZR1466900) and the China Postdoctoral Science Foundation (No. 2021M692423).
Supplementary materials
Supplementary material associated with this article can be found, in the online version, at doi:10.1016/j.cclet.2022.108058.
Figure 1
Variation of total ARGs and resistance mechanisms (A) and specific ARGs (B) in different reactors during long-term FW fermentation with no enzymatic pretreatment and no pH control (R0), with enzymatic pretreatment and no pH control (R1), and with enzymatic pretreatment and acidic-pH operation (R2).
Figure 2
Variations of microorganisms at phylum (A) and genus level (B) in different reactors during long-term FW fermentation with no enzymatic pretreatment and no pH control (R0), with enzymatic pretreatment and no pH control (R1), and with enzymatic pretreatment and acidic-pH operation (R2), and the Network analysis of the co-occurrence between ARGs and microorganisms (C).
Figure 3
Schematic diagram of the ARGs transmission route (A), variation of critical metabolic pathways (B) and the corresponding genetic expressions (C) in different reactors during long-term FW fermentation with no enzymatic pretreatment and no pH control (R0), with enzymatic pretreatment and no pH control (R1), and with enzymatic pretreatment and acidic-pH operation (R2).
Figure 4
RDA analysis of the correlations of ARGs, microbial community, and metabolic pathways in different reactors during long-term FW fermentation with no enzymatic pretreatment and no pH control (R0), with enzymatic pretreatment and no pH control (R1), and with enzymatic pretreatment and acidic-pH operation (R2).

 DownLoad:
DownLoad:


 下载:
下载:





 下载:
下载:
