Citation:
Yumeng Yang, Junfeng Xiang, Hongxia Sun, Hengwen Chen, Hong Zhang, Yalin Tang. New insights into the effect of molecular crowding environment induced by dimethyl sulfoxide on the conformation and stability of G-quadruplex[J]. Chinese Chemical Letters,
2023, 34(3): 107695.
doi:
10.1016/j.cclet.2022.07.038
New insights into the effect of molecular crowding environment induced by dimethyl sulfoxide on the conformation and stability of G-quadruplex
English
New insights into the effect of molecular crowding environment induced by dimethyl sulfoxide on the conformation and stability of G-quadruplex
Beijing National Laboratory for Molecular Sciences, State Key Laboratory for Structural Chemistry of Unstable and Stable Species, Institute of Chemistry, Chinese Academy of Sciences, Beijing 100190, China
b.
University of Chinese Academy of Sciences, Beijing 100049, China
c.
Center for Physiochemical Analysis & Measurement, Institute of Chemistry, Chinese Academy of Sciences, Beijing 100190, China
d.
Department of Cardiology, Guang'anmen Hospital, China Academy of Chinese Medical Sciences, Beijing 100053, China
* Corresponding authors. ** Corresponding author at: Beijing National Laboratory for Molecular Sciences
State Key Laboratory for Structural Chemistry of Unstable and Stable Species
Received Date:
23 May 2022 Accepted Date:
18 July 2022 Revised Date:
13 July 2022 Available Online:
15 March 2023
Abstract:
Various structures of G-quadruplex in biosystems play an important role in different diseases and are often regulated by a variety of molecular crowding environments induced by internal and even external factors (e.g., a solvent). Dimethyl sulfoxide (DMSO), a universal solvent, has been widely used in biological studies and for drug therapy, but little is known regarding its effect on G-quadruplex structure and stability. Here, we report the influence of molecular crowding environment induced by DMSO on the conformation and stability of G-quadruplex structure. We show that the G-quadruplex-forming sequences such as human telomeric sequence, which may have diverse conformations in different environments, tend to convert their topologies to parallel structures under the molecular crowding stimulated by DMSO. Moreover, DMSO can increase the stability of the parallel and antiparallel topologies, especially the parallel G-quadruplex sequence c-kit, but not the hybrid topologies. Further analysis of c-kit using the CD and NMR technique, combined with the unique structural characteristics of c-kit, reveals that the crowding, dehydration and interaction of DMSO are conductive to the formation and stability of the parallel G-quadruplex. The present study suggests that, DMSO, a common solvent used in DNA experiments, may have a nonnegligible influence on the structure and stability of G-quadruplex.
G-quadruplexes are a series of nucleic acids with unique structures, which have drawn great attention as their in vivo existence has been demonstrated and their potential roles in regulating cell growth and proliferation have been unveiled [1-6]. G-quadruplex formation is reported in promoters of different human proto-oncogenes, like c-kit, c-MYC, bcl-2, KRAS, VEGF, and so on [2,7-12]. G-quadruplexes are formed by guanine-rich sequences that fold into four-stranded constructs, where guanine bases make hydrogen bonds with each other to yield square planar structures [13-17]. G-quadruplexes may be intramolecular or intermolecular, and there are mainly three types of topologies: parallel, antiparallel and hybrid, classified by the orientation of their loops and G-tracts [18-22]. During the past two decades, many researches have revealed that the conformation of G-quadruplex can be affected dramatically by internal and even external environmental factors, such as cations, pH, temperature and crowding [23,24].
Real biosystems such as living cells are crowed with a wide range of biospecies, whose concentration is reported to be 50–400 g/L, whereas in most in vitro biochemical studies it is usually less than 1 g/L [23]. The real intracellular environment is extremely different from the diluted solution [25]. These different crowding environments can significantly alter the conformation and stability of G-quadruplex [14,24,26-33]. Furthermore, the altered G-quadruplex topology or conformation may possess different biological functions, despite having the same sequence [19,34]. Because of these features, it is necessary to investigate the state of G-quadruplex under real physiological conditions.
Toward this end, several co-solutes have been used to simulate molecular crowding environments to study the related biomolecular reactions [23,35]. In general, molecular crowding agents need to meet some criteria, for instance, these co-solutes should be almost inert with the target molecule and apt to dissolve in water [23,35,36]. Several large molecules like polyethylene glycol (PEG), polysaccharides, ficoll and dextrans were used to mimic molecular crowding conditions; other small molecules such as ethanol, glycols, amino acids, acetonitrile, betaine and trimethylamine N-oxide at high concentrations can also act as crowding/dehydrating agents, generating molecular crowding environment or decreasing water activity to mimic real cell conditions [26-33,36,37]. Among them, PEG or acetonitrile has been widely used in recent molecular crowding studies [36].
Dimethyl sulfoxide (DMSO), a commonly used organic polar aprotic solvent, is soluble in both aqueous and organic media. Because of its good amphipathicity and relatively low toxicity, DMSO has been extensively used as a universal solvent in biological studies and as a vehicle for drug therapy [38]. Moreover, DMSO has also been considered as a neutral co-solute in related molecular crowding researches [30,37]. However, there are few reports specifically focusing on DMSO, so the effect of DMSO on the conformational transition and stability of G-quadruplex is not fully understood.
In this work, the influence of molecular crowding induced by DMSO on the conformation and stability of G-quadruplex has been investigated using circular dichroism (CD) and NMR spectroscopy. We find that DMSO not only tends to convert G-quadruplexes into their parallel structures, but also, more importantly, selectively stabilizes the c-kit G-quadruplex. Further analysis of c-kit reveals that, besides the effect of hydration and steric crowding, the insidious interaction between DMSO and c-kit plays a significant role in the structural transformation and stabilization of the G-quadruplex. Our results provide a new insight into the influence of DMSO on the structure and stability of G-quadruplex, and promote the understanding of the DMSO effect.
The structural changes of the G-quadruplex formed in the human telomeric sequence d(AG3(TTAG3)3) (Hu22, Table S1 in Supporting information) under different DMSO concentration conditions were first examined with circular dichroism spectroscopy (for details, see Materials and Methods in Supporting information). Hu22 was chosen as a model because the formation and stabilization of the Hu22 G-quadruplex have been shown to inhibit the activity of telomerase, and the telomeric G-quadruplex has now become an attractive target for developing anticancer drugs [39,40]. Previous studies revealed that Hu22 can form in Na+ solution a basket-type antiparallel G-quadruplex, whereas its hybrid G-quadruplex structure can be detected in K+ solution [3,39,40]. Different structural changes caused by crowding have been reported based on this sequence [26,27,31]. However, the change behavior of the Hu22 G-quadruplex in DMSO-induced crowding environment remains poorly understood. Here, we made such a study.
As shown in Fig. 1, the CD spectra of Hu22 differ greatly under the crowding conditions induced by different volume ratios of DMSO in NaCl and KCl solutions, reflecting the distinct conformation alteration of the Hu22 G-quadruplex. It is known that positive peaks at 260 nm and 295 nm in CD spectra are characteristic of parallel and antiparallel G-quadruplex structures [5,41,42], respectively. In the case of a hybrid form, a positive CD band and a shoulder-positive CD band generally appear at 290 nm and about 270 nm, respectively [39,42]. Based on these knowledge and Fig. 1, the Hu22 G-quadruplex mainly exists in antiparallel (Fig. 1A, black line) and hybrid (Fig. 1B, black line) conformations in dilute NaCl and KCl solutions, respectively. In the presence of a low proportion of DMSO (50%, v/v), it can be seen that the conformation of the Hu22 sequence in the K+ environment gradually changes from hybrid to parallel, with a positive peak appearing at ~265 nm; whereas in the Na+ environment, only a small change in the CD spectra is produced, suggesting that Hu22 still maintains the antiparallel structure. This behavior is consistent with the previous reports about the influence of molecular crowding agent of PEG on the conformation of Hu22 [26,31]. When the volume ratio of DMSO is increased to 75%, a significant decrease of the CD signal at 290 nm and a large increase at ~265 nm are observed in both Na+ and K+ environments (Fig. 1), clearly indicating that Hu22 can be transformed into a parallel structure under the present condition. This finding has not been reported to our knowledge, probably due to the less crowded condition used previously. Besides, DMSO itself produced almost no CD signal at the wavelength of more than 255 nm (Fig. S1 in Supporting information). In a word, the above data suggest that the conformational transition of Hu22 to parallel-stranded G-quadruplex can take place in the molecular crowding environment generated by high concentration of DMSO, whether in Na+ or K+ solution.
Figure 1
Figure 1.
CD spectra of Hu22 under different crowding conditions. Hu22 was dissolved in pH 7.4 phosphate buffered saline (PBS) containing 100 mmol/L of NaCl (A) or KCl (B) with different volume ratios of DMSO. Spectra were recorded at ambient temperature of 25 ℃.
Encouraged by the above experimental results, many other G-quadruplexes with different structures (Table S1) were used to investigate their conformations in the crowded solution created by DMSO. As shown in Fig. 2 and Fig. S2 (Supporting information), the CD spectra of hybrid G-quadruplexes (B25, G3T3 and Hu24A) change dramatically in the crowding environment, and DMSO tends to switch their conformation from hybrid to parallel. However, for the typical antiparallel (SPB1 and TBA) and parallel (e.g., H6 and AB1) G-quadruplexes, no obviously change in their conformation is observed in the presence of DMSO, except that the CD signal intensity of some G-quadruplexes (e.g., SPB1 and c-kit) alters slightly. These results also demonstrate that the G-quadruplex-forming sequences with changeable conformations (G3T3, Hu22, Hu24A, B25, etc.) can convert their topologies to parallel forms under the molecular crowding stimulated by 50% DMSO. DMSO at a higher concentration of 75% produced a similar result for B25 (hybrid) and AB1 (parallel) with different representative G-quadruplex structures (Fig. S3 in Supporting information).
Figure 2
Figure 2.
CD spectra of B25 (A), G3T3 (B), SPB1 (C), TBA (D), H6 (E) and AB1 (F) under different crowding conditions. All these oligonucleotides were dissolved in pH 7.4 PBS containing 100 mmol /L KCl without (control) and with 50% (v/v) DMSO. All the spectra were recorded at ambient temperatures of 25 ℃.
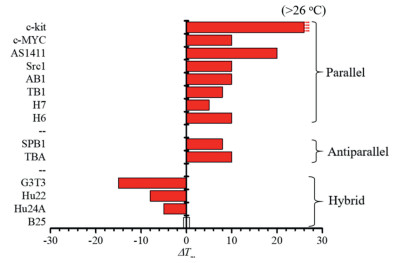
As mentioned above, the structural stability of G-quadruplexes is also affected by the crowding. To gain further insights into the effect of DMSO on the structural stability of G-quadruplexes, the melting curves of various oligonucleotides in pH 7.4 PBS containing 100 mmol/L KCl without (control) and with 50% (v/v) DMSO were measured as a function of temperature ranging from 25 ℃ to 90 ℃ by CD melting assays [32,33]. The melting temperature (Tm) of each G-quadruplex and its melting temperature difference (ΔTm, reflecting the stability change) with and without DMSO are shown in Fig. S4 (Supporting information), Table 1 and Fig. 3. Thereinto, the ΔTm values of parallel G-quadruplexes were obtained by measuring the characteristic CD signal at 262 nm, whereas those of hybrid and antiparallel structures were determined at 290 nm. These data indicate that DMSO exhibits different effects on the stability of the G-quadruplex structures.
Table 1
Table 1.
Stability and conformational changes of various G-quadruplexes under molecular crowding conditions stimulated by DMSO.
For parallel and antiparallel G-quadruplexes, their ΔTm values were increased to varying degrees. As depicted in Table 1 and Fig. 3, most of the G-quadruplexes (c-MYC, Src1, AB1, TB1, H6, H7, SPB1 and TBA) display a ΔTm increment of about 5–10 ℃, and some parallel G-quadruplex sequences such as c-kit and AS1411 show larger ΔTm increments (no less than 20 ℃). Interestingly, the parallel G-quadruplex sequence c-kit is observed to produce the most selective and dramatic ΔTm increment, with a ΔTm value greater than 26 ℃. These results suggest that the stability of the parallel and antiparallel G-quadruplexes, in particular the c-kit structure, can be increased under the crowded environment with 50% (v/v) DMSO.
For hybrid G-quadruplexes, however, their ΔTm values (Table 1, Fig. 3) were decreased, demonstrating that the hybrid structures cannot be stabilized by DMSO. Moreover, under the 50% DMSO crowding solution, the CD signal at 290 nm (indicative of the antiparallel form) of the hybrid structures diminished, while that at 260 nm (indicative of the parallel form) arose with the increase of temperature, implying that the conformation of the hybrid G-quadruplexes (e.g., G3T3, Hu22 and Hu24A) tends to change to their parallel forms; namely, as the temperature rises, the conversion trend will increase. This is consistent with the above results of conformational transition experiments (Fig. 2 and Fig. S2).
On the other hand, for the vast majority of reported cases, the biophysical effects of co-solutes (such as PEG) on G-quadruplex stability are mainly explained by the steric crowding and hydration effect [24,43,44]. However, these mechanisms cannot explain why DMSO can specifically stabilize the c-kit G-quadruplex structure. There may be other possible mechanisms underlying the effects of DMSO on c-kit.
To gain insight into the possible mechanisms, the ability of DMSO to stabilize mutant c-kit G-quadruplex was further evaluated by CD melting assays (Fig. 4 and Table 2). Moreover, PEG200 (polyethylene glycol of the average molecular weight 200), a famous co-solute to mimic steric crowding, was used as a contrast agent under the same experimental conditions. On the other hand, it is known that loops of G-quadruplex structures play an important role in the interaction between G-quadruplexes and ligands [45,46]. According to the literature, the c-kit G-quadruplex structure has a two-residue loop and a five-residue loop [47]. The oligonucleotides mutated at the above two loops, respectively (Table 2). CD spectra showed that these mutated DNA oligonucleotides formed the same parallel conformations as the c-kit G-quadruplex (Fig. S5 in Supporting information).
Figure 4
Figure 4.
CD melting curves of c-kit (A), c-kit-T (B), c-kit-A (C), c-kit-3T (D) and c-kit-4T (E). All the oligonucleotides were dissolved in pH 7.4 PBS containing 100 mmol/L KCl without (control) and with 50% (v/v) DMSO or PEG200. The temperature programming was from 25 ℃ to 90 ℃.
Just as shown in Fig. 4A, the CD melting behaviors of the c-kit G-quadruplex with DMSO and PEG200 are similar, the Tm of the c-kit G-quadruplex in the presence of 50% (v/v) DMSO or PEG200 is much higher than 90 ℃ and the ΔTm value is greater than 26 ℃ (Table 2 and Table S2 in Supporting information). However, for the c-kit with mutation at T (c-kit-T) and A (c-kit-A), DMSO and PEG200 exhibited entirely different experimental results (Figs. 4B and C and Table 2). CD melting results indicated that the stability of c-kit-T and c-kit-A in the presence of PEG200 was much higher than that in the presence of DMSO. PEG200 can efficiently stabilize the structure of the c-kit G-quadruplex with or without mutation. In contrast, in the presence of DMSO, the stability of c-kit-T (ΔTm = 18 ℃) and c-kit-A (ΔTm = 22 ℃) was much lower than that (ΔTm > 26 ℃) of c-kit. The above data demonstrated that the molecular mechanism of DMSO was different from that of PEG200 on stabilizing the G-quadruplex structure, which may be attributed to the different interactions of G-quadruplexes with the co-solutes, similar to the interactions between G-quadruplex's loops and ligands [45,46].
To explore the different interactions of the G-quadruplex's loops with the co-solutes, similar sequences with different loop lengths have been synthesized based on c-kit, including c-kit-3T and c-kit-4T (Table 2). As shown in Figs. 4D, E and Table 2, PEG200 displays good thermal stabilization towards c-kit-3T and c-kit-4T (ΔTm values are greater than 34 ℃ and 35 ℃, respectively); however, DMSO exhibits a mild effect on the stability of c-kit-3T and c-kit-4T (the ΔTm values of both are 24 ℃). Moreover, we also found that in the presence of DMSO, when the temperature of solution was higher than 65 ℃, the CD signal at 260 nm decreased sharply, suggesting that the c-kit-3T or c-kit-4T G-quadruplex was destabilized. Clearly, the thermal stabilization of DMSO and PEG200 is different greatly, and these results indicate that, in addition to molecular crowding and dehydration, other interactions such as the binding (see below NMR experiments) between DMSO and G-quadruplex may also influence the stability of the G-quadruplex in the crowded environment.
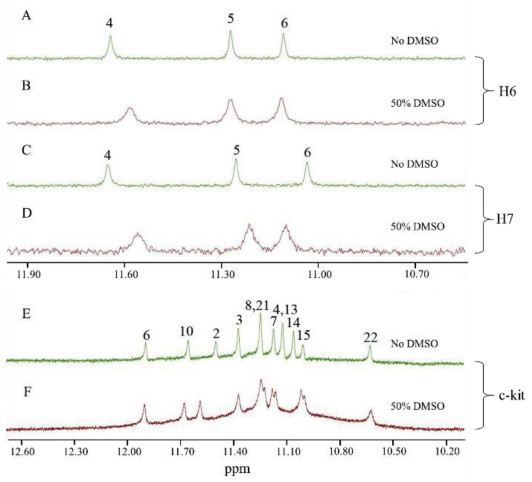
Finally, in order to further understand the effect of DMSO on G-quadruplexes, 1H NMR experiments were performed with H6, H7 (typical intermolecular parallel-stranded quadruplexes) and c-kit (a typical intramolecular parallel-stranded quadruplex), because these sequences exhibit well-resolved and assigned NMR signals in PBS containing K+ solution [47,48].
As shown in Fig. 5, H6 and H7 exhibit three sharp peaks of imino proton signals in the absence of DMSO, which can be assigned to G4, G5 and G6 protons (Figs. 5A and C) [48], respectively. However, addition of 50% (v/v) DMSO causes dramatic line-broadening of these signals (Figs. 5B and D), with less dispersion. It is known that adding DMSO to the dilutedsolution can strongly reduce the exchange between guanine imino protons with other active protons, thereby producing a solvation effect. In other words, the broadened guanine imino proton signals reflect a predominate role of the crowding. Moreover, by careful analysis, the G4 and G6 imino proton signals exhibited upfield and downfield shifts, respectively, especially for H7. The small, but distinct, NMR signal shifts of these guanine imino protons indicated that DMSO may bind to H7.
Figure 5
Figure 5.1H NMR spectra of G-quadruplexes (0.2 mmol/L each) in 100 mmol/L K+ solutions under different crowding conditions. (A) H6; (B) H6 with 50% (v/v) DMSO; (C) H7; (D) H7 with 50% (v/v) DMSO; (E) c-kit; (F) c-kit with 50% (v/v) DMSO.
As an intramolecular parallel G-quadruplex, c-kit shows 10 sharp peaks in the absence of DMSO (Fig. 5E), which correspond to the 12 imino protons; among them, the larger peaks at 11.14 and 11.25 ppm contain two protons each [47]. In contrast to the narrow and well-resolved NMR peaks in the diluted solution, c-kit displays not only broad but also slightly regrouped imino proton peaks under the crowding condition induced by 50% (v/v) DMSO (Fig. 5F). These observations suggest that the parallel c-kit G-quadruplex may form a tighter and more stable conformation under the dehydration and interaction effect of DMSO.
These NMR experimental results were not only in good agreement with the above CD melting experiments, but also more importantly, confirmed that DMSO could interact with G-quadruplex. Therefore, in the crowded environment created by DMSO, the molecular crowding, dehydration effects and interaction may be the main factors that influence the structural transformation and stability of the G-quadruplexes.
In summary, we have studied the influence of the molecular crowding created by DMSO (a universal solvent widely used in biological and pharmacological experiments) on the conformation and stability of G-quadruplexes. It is found that DMSO tends to convert G-quadruplex-forming sequences into their parallel structures. Moreover, DMSO can effectively stabilize the parallel forms (especially the c-kit G-quadruplex structure) as well as the antiparallel structures, but is hardly effective for the hybrid forms. CD melting and NMR data reveal that crowding, dehydration and interaction may be the key factors in the stability and transformation of G-quadruplex structures. So, in normal experiments with this solvent, the influence of DMSO on the structure and stability of G-quadruplex should not be ignored. The present study may be also helpful for better understanding of the DMSO effect on DNA.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
This research was supported by the National Natural Science Foundation of China (Nos. 21977099, 22077123 and 21977096) and China Academy of Chinese Medical Sciences Innovation Fund (No. CI2021A05011). The measurements of CD and NMR Spectroscopy were performed at the Center for Physicochemical Analysis and Measurements in ICCAS. The help from Jie Cui and Aijiao Guan was also acknowledged.
[1]
S. Balasubramanian, S. Neidle, Curr. Opin. Chem. Biol. 13 (2009) 345-353. doi: 10.1016/j.cbpa.2009.04.637
A.T. Phan, V. Kuryavyi, S. Burge, S. Neidle, D.J. Patel, J. Am. Chem. Soc. 129 (2007) 4386-4392. doi: 10.1021/ja068739h
[48]
O.Y. Fedoroff, M. Salazar, H.Y. Han, et al., Biochemistry 37 (1998) 12367-12374. doi: 10.1021/bi981330n
Figure 1
CD spectra of Hu22 under different crowding conditions. Hu22 was dissolved in pH 7.4 phosphate buffered saline (PBS) containing 100 mmol/L of NaCl (A) or KCl (B) with different volume ratios of DMSO. Spectra were recorded at ambient temperature of 25 ℃.
Figure 2
CD spectra of B25 (A), G3T3 (B), SPB1 (C), TBA (D), H6 (E) and AB1 (F) under different crowding conditions. All these oligonucleotides were dissolved in pH 7.4 PBS containing 100 mmol /L KCl without (control) and with 50% (v/v) DMSO. All the spectra were recorded at ambient temperatures of 25 ℃.
Figure 4
CD melting curves of c-kit (A), c-kit-T (B), c-kit-A (C), c-kit-3T (D) and c-kit-4T (E). All the oligonucleotides were dissolved in pH 7.4 PBS containing 100 mmol/L KCl without (control) and with 50% (v/v) DMSO or PEG200. The temperature programming was from 25 ℃ to 90 ℃.


 DownLoad:
DownLoad:





 下载:
下载:






 下载:
下载:
