Pharmaceutical College, Guangxi Medical University, Nanning 530021, China
b.
National Center for International Biotargeting Theranostics, Guangxi Key Laboratory of Biotargeting Theranostics, Collaborative Innovation Center for Targeting Tumor Theranostics, Guangxi Medical University, Nanning 530021, China
c.
Department of Spine Surgery, The Second Xiangya Hospital of Central South University, Changsha 410011, China
wbxyeyy@csu.edu.cn (B. Wang). 1 These authors contributed equally to this work.
Received Date:
03 March 2022 Accepted Date:
15 June 2022 Revised Date:
14 June 2022 Available Online:
15 March 2023
Abstract:
Liposomes have been widely exploited as a drug delivery system in treating tumors because of their advantage to enhance anti-tumor efficacy and reduce side effects. In this study, the tumor-targeted 2-dodecyl-6-methoxycyclohexa-2, 5-diene-1, 4-dione (DMDD, i.e., Averrhoa carambola extractive) liposomes (HA/TN-DLP) were conducted and assessed. HA/TN-DLP showed controllable drug loading (up to 83%) with high stability. In vitro and in vivo studies showed good cell uptake behavior and high inhibition rate of breast cancer compared to free DMDD. HA/TN-DLP might be the suitable for DMDD due to its better advantages in delivery, penetrability, and targeting-tumor capability. For in vivo mouse model tests, HA/TN-DLP effectively inhibited tumor growth compared to free DMDD. Further analyses indicated that HA/TN-DLP inhibited the glycerophospholipid metabolism pathway by reducing the biosynthesis of phosphatidylcholine and 1-acyl-sn-glycero-3-phosphocholine through regulating the expressions of CEPT1 and LYPLA1, and inhibited tumor cell growth by regulating the PI3K/Akt and NF-κB signaling pathways. In conclusion, the obviously enhanced antitumor effect further demonstrated that HA/TN-DLP may be a promising tumor-targeting agent.
Breast cancer is one of the most common malignant tumors in women worldwide [1]. There are many methods to treat breast cancer, such as surgery, endocrine therapy, chemotherapy and radiation, but none of these methods can achieve a perfect therapeutic effect. Therefore, developing safe and effective neoadjuvant therapy drugs, screening drug targets, and elucidating the molecular mechanism of drug intervention in breast cancer are effective strategies for the clinical treatment of breast cancer [2].
A typical targeted drug delivery system usually consists of an anticancer drug and a targeting ligand, which can specifically bind to tumor markers that abundantly express in cancer cells [3, 4]. An ideal tumor marker for targeted therapy should be a membrane protein that is over-expressed on the surface of cancer cells, with relatively low expression in normal tissue. The CD44 molecule is an important surface marker molecule on the surface of breast cancer cells, which plays an important role in the occurrence, development and metastasis of breast cancer. The expression level of CD44 molecule is closely related to the malignancy and prognosis of breast cancer [5]. As the natural ligand of CD44 molecule, hyaluronic acid (HA) has a strong affinity with CD44 molecule. It achieves the purpose of targeted drug delivery through the receptor-ligand mechanism and is therefore commonly used in breast cancer targeted drug delivery studies [6, 7]. Liposome has been considered as a good drug delivery system due to its good biocompatibility and biodegradability and its benefit to improve the efficacy and reduce the toxicity of drugs. Liposomes can resolve the problem of poor water solubility regardless of the hydrophilic or hydrophobic drugs [8]. Taken together, preparing a liposome with HA might be able to improve the stability of the drug, and enhance the targeting-tumor potential as well.
In our previous studies, we have isolated an ingredient from the roots of Averrhoa carambola L. and identified it as 2-dodecyl-6-methoxycyclohexa-2, 5-diene-1, 4-dione (DMDD). We have found that DMDD could inhibit breast cancer cell proliferation and migration [9, 10]. which means it might be a promising agent for the treatment of breast cancer. Also, we found that DMDD had several defects, such as poor solubility, low bioavailability, and poor tumor-targeting ability, which should limit its application in treating breast cancer. To resolve this defect, we tried to synthesize the tumor-targeting DMDD liposomes containing TAT-NBD (TN: a fusion peptide that can selectively block NF-κB activating pathway) and HA fragments, assess the quality of liposomes, and evaluate the inhibitory effects of the DMDD liposomes on breast cancer.
DMDD was obtained from the root of Averrhoa carambola L. as previously described [11]. Common DMDD liposomes (DLP), liposomes contain TN and DMDD (TN-DLP) and liposomes contain HA, TN and DMDD (HA/TN-DLP) were prepared using the ethanol injection way [12], as shown in Fig. 1A. Briefly, the yolk lecithin/cholesterol/TN-PEG2000-DSPE/DMDD (100:14:1:14) were dissolved in ethanol as the oil phase. Under magnetic stirring, the oil phase was slowly dropped into the same volume of PBS at 52.5 ℃ for 30 min. Then the TN-DLP treated by miniature ultrasonic probe for 10 min at 100 W, and the TN-DLP was purified by dialysis for 24 h to remove unreacted substances. The preparation of DLP was the same as TN-DLP, except that TN-PEG2000-DSPE was not added to the oil phase. HA/TN-DLP was prepared by dispersing HA solution in TN-DLP at a HA/lipid mass ratio of 1:1 and then vortex for 30 min at 25 ℃. The 4T1 cells were used to establish the subcutaneous xenograft mouse model of breast cancer. And the liposomes of DLP, TN-DLP and HA/TN-DLP were treated to animals. The experiments were performed with approval from the Institutional Animal Care and Use Committee of Guangxi Medical University. The inhibition rate of tumor growth was calculated. Transcriptomic analysis was carried out to predict the potential targets and then the predictions would be verified by the following experiments.
Figure 1
Figure 1.
The preparation and characterization of DLP, TN-DLP and HA/TN-DLP. (A) The preparation of liposomes. (B) The size and TEM image of liposomes. (C) Uptake of 4T1 cells investigated by laser scanning confocal microscope. Blue and red signals indicate Hoechst 33, 342 and RHB. (D) Uptake of 4T1 cells investigated by fluorescent spectrophotometer. Data were presented as mean ± SEM.
HA is widely used in anticancer drug delivery since it is suitable for degradability, good bioactivity, and excellent cytocompatibility, nontoxic, and non-immunogenic; moreover, HA receptors are overexpressed on many tumor cells [13, 14]. Exploiting this ligand-receptor interaction, HA was used to target CD44-overexpressing cells, to improve anticancer therapies [15]. TN peptide, a fusion peptide of NF-κB essential modulator (NEMO)-binding domain peptide (NBD) and cell-penetrating peptide (TAT), can selectively block NF-κB activating pathway, resulting in tumor growth inhibition [16, 17]. In the present study, we prepared three DMDD liposomes: a common liposome (DLP), a novel TN-modified liposome (TN-DLP), and an anionic HA/TN-DLP. Generally, the quality of liposomes was assessed with the particle size, polydispersity index (PDI), zeta potentials and encapsulation efficiency (EE%). We found that the three liposomes had similar characteristics to some extent: (1) The particle size distribution was uniform and normalized to a single-peak distribution; (2) the PDI ranged from 0.1 to 0.3, and the surface liposome particle size distribution range was narrow; (3) the zeta potentials were all negatively charged, which means that the electrostatic repulsion can prevent the particles from accumulating and is beneficial for preserving liposomes; (4) the morphology observed by TEM revealed a relatively clear spherical structure and uniform size distribution and a fingerprint-like structure typical of a liposome; (5) all the liposomes had a high EE%, which was suitable for DMDD delivery (Fig. 1B and Table S1 in Supporting information). On the other hand, HA/TN-DLP showed significant advantages among the liposomes: (1) The examinations of uptake and penetrability showed that HA/TN-LP could remarkably promote cellular uptake (Figs. 1C and D); (2) the UPLC-qTof-MS with UNIFI software analysis was showed that DMDD in the liposome of HA/TN-DLP in the plasma and tumor tissue could be found; however, DMDD without the liposome could not be detected. The analysis demonstrated that HA/TN-DLP was beneficial to carrying DMDD into the bloodstream and targeting tumor tissues, it can be assumed that HA/TN-DLP had sustained-release and tumor targeting effects, which undoubtedly favor to the enhancing anti-tumor effect (Fig. S1 in Supporting information).
To further assess their advantage in enhancing the drug potency, the effects of the liposomes on breast cancer were investigated. The tumor volumes and the tumor weight in the DMDD liposomes groups were lower than the model group (Figs. 2A and B). Pathological examination showed that administration with DMDD liposomes caused different degrees of tumor necrosis: loose arrangement of tumor cells, cell membrane shrinkage, decreased volume, nuclear shrinkage, and fragmentation, chromatin aggregation (Fig. 2C). In addition, apoptosis is of great significance to tumor cell proliferation and tumor growth. The abnormal level of apoptosis is a biological characteristic that reflects the malignant degree of tumor cells. The lack or over-expression of apoptosis regulatory factors may lead to abnormal apoptosis, the rapid proliferation of tumor cells, and even cancer [18]. Bcl-2 is an anti-apoptotic protein and the imbalance of Bcl-2 is considered as a condition for the occurrence of apoptosis [19]. In this study, the TUNEL staining revealed that DMDD liposomes markedly increased the positive apoptotic cells, which might be attributed to the regulation of caspase-3 and Bcl-2 families (Figs. 2D and E). The data above suggested that the DMDD liposomes could inhibit the growth of breast cancer and promote tumor cell apoptosis. From these results, we also found that under the same administered dose of DMDD, HA/TN-DLP had better inhibitory effects than the other ones, which may be due to the targeted modification of HA and TN that could improve the delivery efficiency of DMDD and promote the uptake of cells, which undoubtedly favor to the enhancing anti-tumor effect of DMDD.
Figure 2
Figure 2.
DMDD liposomes suppress the growth of 4T1 cell xenografts. (A) The tumor volume curve after treatment. (B) The tumors. (C) HE staining of breast cancer tumor tissues (magnification: 400×). Yellow circles: apoptotic tumor. (D) Microscopic observation of TUNEL results in tumor tissues from breast cancer (magnification: 400×). (E, F) The effects of drugs on the protein and mRNA expressions of Bcl-2 in tumor tissues.
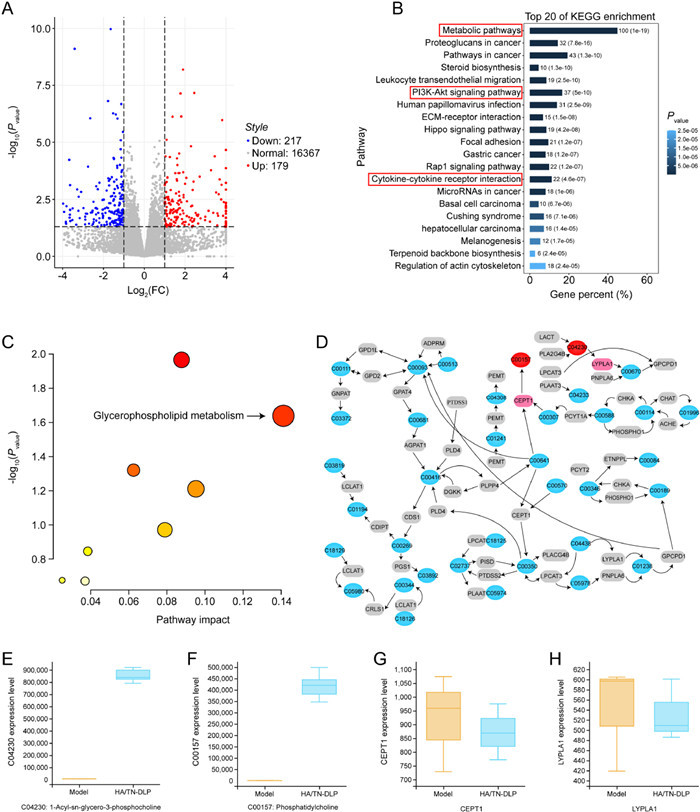
In the above study, we have clarified the inhibitory effect of HA/TN-DLP on breast cancer. To further understand the possible protective mechanisms, transcriptomics analysis was performed in the next study. In the current study, transcriptomic analysis was performed to predict the potential targets or signaling pathways, and then multiple examinations were carried out to verify the predictions. The transcriptomic analysis revealed that there were 396 differentially expressed genes (DEGs) between the model and HA/TN-DLP groups: 179 down-regulated genes and 217 up-regulated genes. In addition, the gene ontology (GO) and KEGG pathway analysis demonstrated that these DEGs could be enriched in the metabolic pathways, PI3K-Akt signaling pathway, and cytokine-cytokine receptor interaction, suggesting that HA/TN-DLP might affect these pathways and thereby ameliorate breast cancer to some extent (Figs. 3A and B).
Figure 3
Figure 3.
Integrated analysis of transcriptomics and metabolomics. (A) Volcano plot. (B) KEGG pathway analysis. (C) The comprehensive analysis of gene and metabolic profiles was carried out using MetaboAnalyst 5.0 database. (D) The glycerophospholipid metabolic pathway. (E-H) The glycerophospholipid metabolic pathway-related metabolites and target genes.
Based on the prediction of the transcriptomic analysis (HA/TN-DLP could target the metabolic pathways), we further tried to verify whether and how HA/TN-DLP affected metabolism. The metabolomics analysis showed a significant difference in the distribution of total ions between the model and HA/TN-DLP groups; there were 91 differential metabolites between them, mainly being enriched in 9 pathways. Considering P-value and impacting index (P < 0.05 and impact > 0.1), the glycerophospholipid metabolism pathway was regarded as the most important relevant pathway. Interestingly, we found that HA/TN-DLP regulated the glycerophospholipid metabolism pathway likely by modulating the synthes is of phosphatidylcholine (PC) and 1-acyl-sn-glycero-3-phosphocholine (LysoPC) (Fig. S3 in Supporting information). In addition, we want to know the targets that HA/TN-DLP regulated to understand the potential mechanism of HA/TN-DLP on this pathway. The integrative analysis of transcriptomics and metabolomics showed that HA/TN-DLP could affect several key metabolites and genes in the network of sphingolipid metabolism. As shown in Figs. 3C–H, the target metabolites included C00157 (phosphatidylcholine) and C04230 (1-acyl-sn-glycero-3-phosphocholine); and the target genes were CEPT1 and LYPLA1. Moreover, the quantitative analysis revealed that HA/TN-DLP could enhanced these target metabolites content and inhibit the target genes expression level, which suggested that HA/TN-DLP alleviated glycerophospholipid metabolism by regulating the expression of the target genes above. Phosphocholine (PC) is the key lipids of cell membrane, which are required to maintain cellular structure, provide energy, and are involved in cell signaling [20]. Manipulation of glycerophospholipid metabolism is related to signaling networks in the regulation of cell growth, proliferation, differentiation, survival, apoptosis, inflammation, motility, and membrane homeostasis [21, 22]. At the same time, Manipulation of glycerophospholipid metabolism can alter membrane composition and permeability which cause the development and progression of many diseases including a variety of cancers proliferation [23, 24]. Our experiments revealed that HA/TN-DLP inhibited breast cancer in mice by regulating glycerophospholipid metabolism.
As the transcriptomic analysis, the PI3K/Akt signaling pathway may also be involved in the regulatory mechanism of the liposomes. The activation of the PI3K/Akt signaling pathway plays a crucial role in the occurrence and development of a variety of cancer and participates in the biological processes of cell proliferation, differentiation, and apoptosis. When stimulated, the PI3K is activated, turning into PI3K phosphorylated phosphatidylinositol 3, 4-triphosphate (PIP3). PIP3 promotes Akt phosphorylation, thereby activating the target of mTOR or PTEN, which regulates multiple signal transduction related to apoptosis [25, 26]. Phosphorylated mTOR also awakens the downstream protein P70S6K to promote mRNA coding, translation and transcription, regulating cell growth and proliferation [27]. In this study, DMDD liposomes significantly inhibited the PI3K/Akt signaling pathway in tumor tissue, as evidenced by the significantly decreased phosphorylation of PI3K, Akt, and mTOR, as well as the mRNA levels of PI3K, Akt, mTOR, and P70S6K (Fig. S4 in Supporting information). These analyses above confirmed that HA/TN-DLP could inhibit the PI3K/Akt signaling pathway.
In addition, the transcriptomic analysis demonstrated that DMDD liposomes might affect cytokine-cytokine receptor interaction. It is well known that pro-inflammatory cytokines can induce the expansion of inflammatory reaction, leading to the NF-κB activation and thereby promoting the progression of metastatic tumors and malignant [17]. It has previously been reported that abnormal activation of NF-κB is closely associated with the malignancy of breast cancer, and blockade of the aberrantly activated NF-κB could trigger apoptosis in breast cancer cells [28, 29]. In the current study, DMDD liposomes caused a significant reduction in the levels of cytokines (IL-6, IL-12 and TNF-α) in the serum of mice and significantly inhibited the protein expression of p-NF-κB p65, MyD88, p-IKKα/β, and p-ⅠκBα. Similarly, qPCR results also showed that DMDD liposomes significantly reduced the expression of key genes in the NF-κB signaling pathway. These results indicated that DMDD liposomes treatment inhibited the NF-κB pathway, leading to the reduction of cytokines (Fig. S5 in Supporting information).
In summary, the tumor-targeting DMDD liposomes are successfully synthesized. The results confirmed that the optimized DMDD liposomes have 100 nm size and good DMDD loading efficiency. The UPLC-qTof-MS with UNIFI software analysis showed that HA/TN-DLP had sustained-release and tumor targeting effects, which undoubtedly favor to the enhancing anti-tumor effect. For in vivo studies, HA/TN-DLP displayed significantly inhibitive effect against breast cancer. Moreover, the transcriptomics predicts that HA/TN-DLP inhibits breast cancer likely by targeting the metabolic pathways, PI3K/Akt signaling pathway, and cytokine-cytokine receptor interaction (being regulated by the NF-κB signaling pathway), which are verified by the metabolomic analysis and the relevant assays (Fig. 4). Our findings demonstrate that HA/TN-DLP may be a promising tumor-targeting agent.
Figure 4
Figure 4.
Preparation and evaluation of DMDD liposome in ameliorating breast cancer: based on transcriptomics and metabolomics. (A) Synthetic scheme of DMDD liposome. (B) Schematic diagram of mice tumor-bearing and treatment time points and (C) the effects of DMDD liposomes on signaling pathways.
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
This study was funded by the Guangxi Key Laboratory of Biotargeting Theranostics (No. GXSWBX201804), the State Project for Essential Drug Research and Development (No. 2019ZX09301132), Guangxi Science and Technology Bases and Talent Special Project (No. AD17129062), Guangxi Key Research and Development Project (No. AB20117001), Guangxi First-class Discipline Project for Pharmaceutical Sciences (No. GXFCDP-PS-2018), Natural Science Foundation of China (Nos. 81760665, 81460205), Guangxi Science and Technology Base and Talent Project (No. AD17195085).
Supplementary materials
Supplementary material associated with this article can be found, in the online version, at doi:10.1016/j.cclet.2022.06.041.
[1]
X. Jiang, H. Tang, T. Chen, J. Gynecol. Oncol. 29 (2018) e7. doi: 10.3802/jgo.2018.29.e7
A. Norambuena, C. Metz, L. Vicuña, et al., J. Biol. Chem. 284 (2009) 12670–12679. doi: 10.1074/jbc.M808949200
[25]
R.L.B. Costa, H. Han, W.J. Gradishar, Breast Cancer Res. Treat. 169 (2018) 397–406. doi: 10.1007/s10549-018-4697-y
[26]
P. Rusquec, C. Blonz, J.S. Frenel, Ther. Adv. Med. Oncol. 12 (2020) 1758835920940939. doi: 10.1177/1758835920940939
[27]
J.C. Coffey, J. Wang, M.J.F. Smith, et al., J. Biol. Chem. 280 (2005) 20968–20977. doi: 10.1074/jbc.M414696200
[28]
X. Zhen, H. Choi, J. Kim, et al., Biomolecules 10 (2020) 245. doi: 10.3390/biom10020245
[29]
Z. Orlova, F. Pruefer, R.C. Oropeza, et al., Biochim. Biophys. Acta Mol. Cell Res. 1866 (2019) 598–611. doi: 10.1016/j.bbamcr.2019.01.002
Figure 1
The preparation and characterization of DLP, TN-DLP and HA/TN-DLP. (A) The preparation of liposomes. (B) The size and TEM image of liposomes. (C) Uptake of 4T1 cells investigated by laser scanning confocal microscope. Blue and red signals indicate Hoechst 33, 342 and RHB. (D) Uptake of 4T1 cells investigated by fluorescent spectrophotometer. Data were presented as mean ± SEM.
Figure 2
DMDD liposomes suppress the growth of 4T1 cell xenografts. (A) The tumor volume curve after treatment. (B) The tumors. (C) HE staining of breast cancer tumor tissues (magnification: 400×). Yellow circles: apoptotic tumor. (D) Microscopic observation of TUNEL results in tumor tissues from breast cancer (magnification: 400×). (E, F) The effects of drugs on the protein and mRNA expressions of Bcl-2 in tumor tissues.
Figure 3
Integrated analysis of transcriptomics and metabolomics. (A) Volcano plot. (B) KEGG pathway analysis. (C) The comprehensive analysis of gene and metabolic profiles was carried out using MetaboAnalyst 5.0 database. (D) The glycerophospholipid metabolic pathway. (E-H) The glycerophospholipid metabolic pathway-related metabolites and target genes.
Figure 4
Preparation and evaluation of DMDD liposome in ameliorating breast cancer: based on transcriptomics and metabolomics. (A) Synthetic scheme of DMDD liposome. (B) Schematic diagram of mice tumor-bearing and treatment time points and (C) the effects of DMDD liposomes on signaling pathways.


 DownLoad:
DownLoad:



 下载:
下载:





 下载:
下载:
