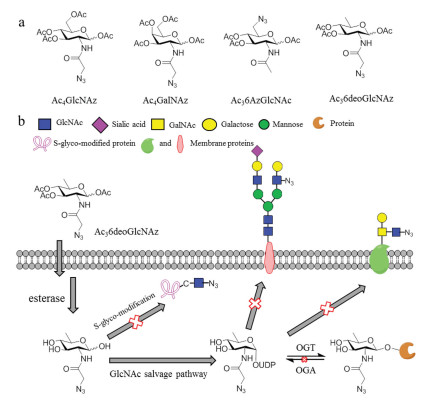
Figure 1.
MCRs for O-GlcNAcylation. (a) MCRs that have been reported and the newly synthesized MCR of Ac36deoGlcNAz; (b) The characterization of Ac36deoGlcNAz.
Ac36deoGlcNAz could selectively label O-GlcNAc modified proteins with minimal S-glyco-modification
Lu Zheng , Wei Cao , Biao Dou , Xueke Zeng , Mingya Cao , Jiajia Wang , Xia Li

O-Linked β-N-acetylglucosamine (O-GlcNAc) modification is a dynamic post-translation modification, which attaches a monosaccharide GlcNAc to the residues of serine and threonine in the nucleocytoplasmic proteins [1]. The process of this modification is controlled by a pair of enzymes: O-GlcNAc transferase (OGT) [2, 3] and β-D-N-acetylglucosaminidase (OGA) [4]. OGT is responsible for the addition of UDP-GlcNAc to Ser/Thr residues and OGA catalyzes the hydrolysis of O-GlcNAc from proteins. Protein O-GlcNAcylation is ubiquitous involving in regulation of the protein translation, signal transduction, gene expression, cell cycle regulation [5-9]. An increasing amount of evidence shows that aberrant O-GlcNAc modification contributes to the pathological processes of human diseases, including cancer, diabete and neurodegenerative disease [10-13]. Based on these important links, more detailed efforts are required to comprehensively elucidate the precise function and molecular mechanisms of O-GlcNAc in disease, which will further reveal the role of O-GlcNAc as a potential target in clinical therapy.
Characterization of O-GlcNAcylated proteins of interest is often the first step before performing functional assays. The challenges in detection and analysis of O-GlcNAcylation attribute to low abundance, stoichiometric modification and reversible regulatory, which have been partially addressed over the last two decades with the exploitation of metabolic and chemoenzymatic labeling approaches in combination with tandem mass spectrometry techniques. The platform based on metabolic chemical reporters (MCRs) has emerged as a useful tool for probing O-GlcNAcylation by chemically incorporating unnatural chemo-selective handles. This method has been successfully developed in living systems largely due to the substrate promiscuity of enzymes in the GlcNAc and/or GalNAc salvage pathway, UDP-glucose 4-epimerase/UDP-galactose 4-epimerase (GALE) and OGT [14]. Once inside the cells, the MCR is processed by the corresponding enzymes to provide glycans bearing the chemical handle that will react with fluorescent dyes or biotin through bioorthogonal reaction for visualization or affinity purification. Tetraacetylated N-azidoacetyl-glucosamine (Ac4GlcNAz) is the first MCR reported for metabolic labeling of intracellular O-GlcNAcylation. Unfortunately, the rate-limiting enzyme of UDP-N-acteylglucosamine pyrophosphorylase AGX1/2 limits the labeling efficiency of Ac4GlcNAz. Another classic MCR is Ac4GalNAz that results in robust labeling of O-GlcNAcylated proteins, however, it could also incorporate into mucin O-glycosylation on the cell surface [15]. These observations raising the necessity to develop novel MCRs that are selective for one kind of interested glycan labeling. We and others have successfully developed series of MCRs for selectively labeling O-GlcNAcylated proteins. For example, in our group, peracetylated 4-deoxy-GlcNAz was rationally designed to promote the selectivity by removal of hydroxyl group at C4 given it was essential for the α/β-1, 4-linkage in the N/O-glycan on the cell surface [16]. Moreover, subtle modification at the 6-position of GlcNAc, Ac36AzGlcNAc, also demonstrated high selective for O-GlcNAc modification (Fig. 1a) [17]. Notably, the good degree of tolerance of enzymes in salvage pathway and the OGT promiscuity toward the unnatural MCRs have been applied in identification of O-GlcNAcylated proteins in various cell lines, these successes encourage us to develop more and new MCRs based on rational hypothesis and design.
We previously synthesized a set of glycopeptides with the method that incubation of full-length hOGT and chemoenzymatically synthesized UDP-GlcNAc derivatives. The results indicated that UDP-6deo-GlcNAc was a better substrate for hOGT than UDP-6-azido-GlcNAc, the glycopeptides yields were 63.1% and 23.8%, respectively [18]. These discoveries implied that a MCR of Ac36deoGlcNAc incorporated a bioorthogonal handle will be a more efficient probe for O-GlcNAcylation than 6-azido-GlcNAc. Herein, we describe the synthesis and characterization of Ac36deoGlcNAz as a MCR for O-GlcNAcylation. Our results demonstrated that Ac36deoGlcNAz could robustly label intracellular proteins rather than cell-surface glycans by western blot and flow cytometry analysis. In addition, this labeling was strongly regulated by OGT, while the resultant O-GlcNAc residues could not be removed by OGA. Interestingly, enzyme of N-acetylhexosamine 1-kinase (NahK) from Bifidobacterium longum has been reported for the formation of GlcNAc-1-phosphate [19], the recombinant plasmid of which was transfected and overexpressed in 293T cell could predominately enhance the labeling. Notably, both the results of western blot in vitro and proteomics supported the idea that Ac36deoGlcNAz could significantly reduce S-glyco-modification (Fig. 1b). Taken together, it is suggested that MCR of Ac36deoGlcNAz we developed here is a robust and selective probe and could be applied to O-GlcNAc proteomic identification.
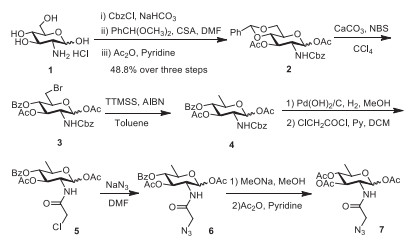
As shown in Scheme 1, Ac36deoGlcNAz was successfully synthesized over ten steps in a totally yield of 4.6%. Briefly, glucosamine hydrochloride was selectively protected with benzyl carbonochloridate (CBz), which was further fully protected in two steps to provide 4, 6-O-benzylidene-1, 3-O-diacetylated-N-CBz-Glcosamine 2. This intermediate 2 was then subjected to selectively benzylidene opening to furnish a bromide contained intermediate that ready to afford the 6-deoxy product 4. After reduction, the CBz protecting group was elaborately removed in the presence of Pd(OH)2/C to generate free -NH2, which was further reacted with chloroacetyl chloride and then converted to azide-substituted product 6. Zemplen deacetylation reaction conducted with sodium methoxide in methanol provided free sugar, which was fully esterified in the presence of anhydride to provide the desired products of Ac36deoGlcNAz 7.
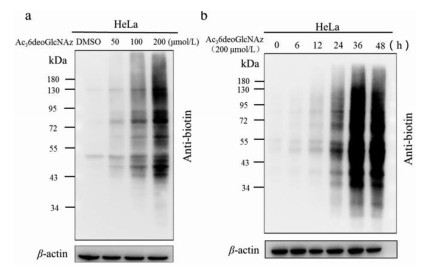
With Ac36deoGlcNAz in hand, we first evaluated its cytotoxicity by treating 293T and HeLa cells at various concentrations for 48 h, which was generally regarded as an experiential time for efficient labeling (Fig. S1 in Supporting information). There was no significant cytotoxicity observed for Ac36deoGlcNAz even at concentrations up to 200 µmol/L in both cultured cells, which demonstrated that the newly developed MCR of Ac36deoGlcNAz was safe to further investigate. We next set out to determine if it could label proteins in mammalian cells by incubation HeLa cells with various concentrations of Ac36deoGlcNAz for 24 h, followed by click reaction with Biotin-PEG4-Alkyne and analysis of the lysates by Western blot. Ac36deoGlcNAz exhibited higher incorporation with the increasing concentration of MCR and 200 µmol/L Ac36deoGlcNAz could achieve efficient label effects with no cytotoxicity (Fig. 2a). Using this as a reference, incubation of HeLa cells with 200 µmol/L Ac36deoGlcNAz for different lengths of times indicated that significantly higher incorporation in a time-dependent manner and highest at 36 h, no more obvious enhancement even incubation with more time (Fig. 2b). Eventually, Ac36deoGlcNAz could effectively label cell proteins at 200 µmol/L for 36 h. In addition, to exclude this labeling only occurs in HeLa cells, Ac36deoGlcNAz was further performed in a panel of cell lines including 293T, B16, 4T1, A549, H1299 and MDA-MB-231 cells. In all tested cell lines, Ac36deoGlcNAz exhibited a diversity of the pattern and intensity of modified proteins, raising the possibility that a proper MCR should be exquisitely selected when labeling in specific cell lines (Fig. S2 in Supporting information).
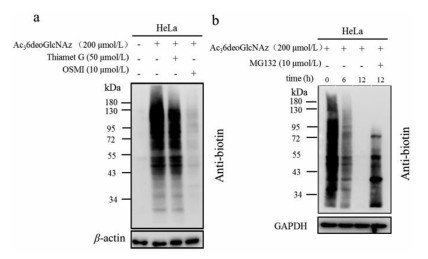
Since Ac36deoGlcNAz was rationally designed for O-GlcNAc modifications, we subsequently tested if this incorporation was affected by the enzymes of OGT and OGA that regulate the O-GlcNAcylation in cells. Therefore, HeLa cells were treated with OGT or OGA inhibitors in the presence of 200 µmol/L Ac36deoGlcNAz for 36 h, followed with click reaction. Using western blot analysis, we found that OGT-inhibitor treatment resulted in significant loss in labeling signal, confirming that the majority of Ac36deoGlcNAz labeling was the result of OGT activity. While OGA-inhibitor contributed minimal to the labeling intensity, which demonstrated that OGA was not responsible for the removal of the protein incorporation and further revealed the rigorous substrate of OGA (Fig. 3a). These results were highly in accordance with our previous glycopeptide results [18]. To determine the turnover of protein medication of Ac36deoGlcNAz, pulse-chase experiment was employed. Specifically, HeLa cells were treated with 200 µmol/L Ac36deoGlcNAz for 36 h and then the cells were washed and fresh media was added. Cells were harvested at indicated time lengths and subjected to click reaction. Western blot analysis showed that a steady loss of protein labeling over the course of 12 h. To confirm this loss of signal was not related with OGA, 10 µmol/L proteasome inhibitor MG132 was added in the culture medium and an obviously stabilized signal was observed, showing the conclusion that 6deoGlcNAz-modified proteins were preferentially degraded by the proteasome (Fig. 3b).
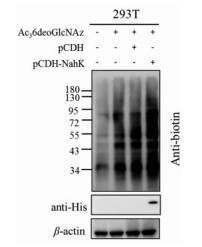
Matthew's group has disclosed that 6AzGlcNAc is a specific MCR for O-GlcNAcylation through direct phosphorylation of its 1-hydroxy by the enzyme of phosphoacetyl glucosamine mutase, because it cannot be transformed into the corresponding UDP-sugar by salvage pathway. Therefore, the labeling efficiency of 6-site medication GlcNAc was slightly lower than the classic Ac4GlcNAz. Based on this hypothesis, it is believed that Ac36deoGlcNAz could also effectively label in the same way. Moreover, our previous results revealed that 6-hydroxy was not necessary for OGT activity [18] and generation of 6deoGlcNAc-1-P in moderate yield (75%) in comparison to natural GlcNAc (90%) [19]. To promote the labeling efficiency of Ac36deoGlcNAz, we constructed a recombinant pCDH plasmid containing NahK gene from bacterial (Fig. S3 in Supporting information), the enzyme responsible for converting 6deoGlcNAz to 6deoGlcNAz-1-P [19], which was transfected into 293T cells in expectation of expressing NahK protein to increase the level of 6deoGlcNAz-1-P. After incubation of Ac36deoGlcNAz with transfected 293T cells for 36 h, and with a pCDH as control, cells were collected and subjected to click reaction. Western blot analysis exhibited that NahK was overexpressed and resulted in improved protein labeling of 6deoGlcNAz (Fig. 4). In short, 6deoGlcNAz labeling was increased in 293T cells with overexpressed NahK leading to higher conversion to 6deoGlcNAz-1-P, and then converted to UDP-6deoGlcNAz by the enzyme AGX1.
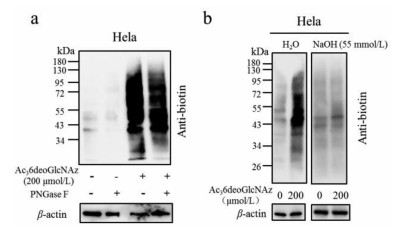
The O-acetyl groups facilitate the membrane permeability of MCRs which can be removed by endogenous esterase enzymes to generate a free sugar for the synthesis of UDP-sugar through the associated salvage pathway. Acetyl protected groups greatly improve the cellular uptake allowing for robust labeling at low concentration. This practiced strategy was challenged since the Chen lab discovered that per-O-acetylated MCRs resulted in abundance of S-glyco-modification proteins in the cells causing interference with proteomic identification [20]. The detailed mechanism was also well investigated, suggesting the need of developing novel MCRs with high efficiency and more importantly, without inducing artificial S-glyco-modification [21]. To detect the specificity of Ac36deoGlcNAz, cell lysates were incubated with varied unnatural monosaccharides. After 2 h at 37 ℃, the lysates were reacted with biotin-alkyne, followed with western blot analysis. As two controls, Ac4GlcNAz and Ac4GalNAz showed significant labeling, while Ac36deoGlcNAz resulted in negligible signal (Fig. 5a). When the cell lysates pretreated with iodoacetamide to block cysteine residues, and then followed with the protocol above, the results demonstrates that all the signals are abolished in the three MCRs treatment (Fig. 5b). Taken together, these results exhibited that Ac36deoGlcNAz was a robust MCR and do not induce non-specific S-glyco-modification.

To further determine if Ac36deoGlcNAz labeling was mainly N- or O-linked to proteins, HeLa cells were treated with 200 µmol/L Ac36deoGlcNAz for 36 h and the collected lysates were conducted with click reaction, followed with separation of proteins by sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred the proteins to a polyvinylidene fluoride (PVDF) membrane in duplicate. One membrane was subjected to H2O and another to NaOH at 50 ℃ for 16 h. After analysis with HRP-linked streptavidin, significant labeling signal loss of β-elimination was observed after treatment with NaOH, which suggested the indeed labeled proteins most likely through of O-linkages (Fig. 6a). Moreover, the cell lysates treated with Ac36deoGlcNAz were incubated with either H2O or PNGase-F before click reaction. The western blot results indicated that Ac36deoGlcNAz did not incorporate into N-linked glycans (Fig. 6b). Finally, to further detect the location of Ac36deoGlcNAz labeling, we set out to culture the HeLa cells with Ac4GalNAz, Ac4ManNAz and Ac36deoGlcNAz (200 µmol/L) for 36 h and Ac4GlcNAc as a negative control. After conjugation with DBCO-488 fluorescence, flow cytometry analysis was employed to confirm the labeling location of MCRs. No labeling over background was observed with Ac36deoGlcNAz, while significant labeling signals were observed with Ac4GalNAz and Ac4ManNAz (Fig. S4 in Supporting information). Taken together, the results indicated that Ac36deoGlcNAz predominately labeled intracellular O-GlcANc modifications.
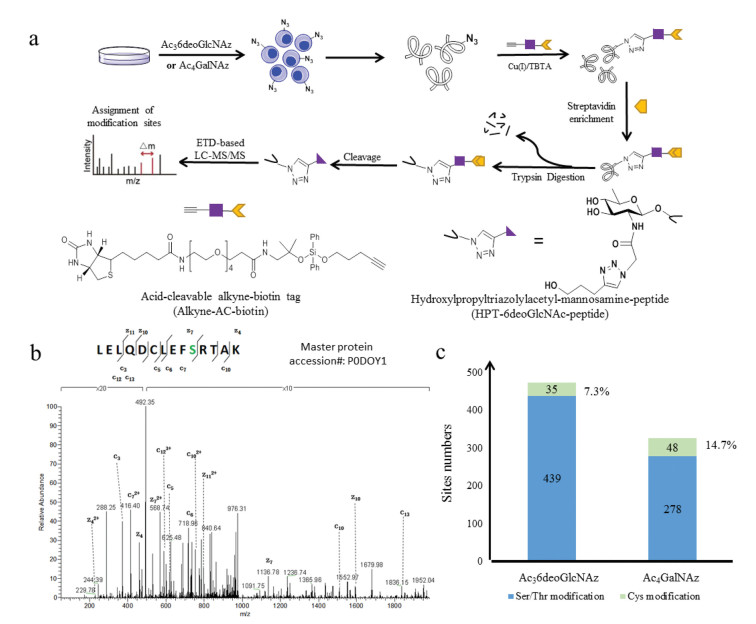
Finally, Ac36deoGlcNAz was used in B16 cells for proteomic identification of O-GlcNAcylated proteins and the whole workflow was illustrated in Fig. 7a. Briefly, B16 cells were treated in triplicate with 200 µmol/L Ac36deoGlcNAz, Ac4GalNAz or Ac4GlcNAc as control for 36 h. The lysates were reacted with biotin-AC-alkyne, followed by streptavidin enrichment and on-bead trypsin digestion. After removal the unmodified peptides, the desired glycopeptides were released and collected for LC-MS/MS analysis. Enriched proteins were characterized for O-GlcNAc labeling as following threshold criteria: (1) Proteins should be identified by at least 1 unique peptide in each of the three data sets; (2) A total of 3 spectral counts in the sum of three replicate data sets. (3) The sum of spectral counts of the MCR-treated samples must be 3-times greater than those in the control group. Based on these criteria, we totally identified 474 and 326 modification sites in the form of 6deoGlcNAz and GalNAz with high confidence, respectively, and 192 O-GlcNAc sites were overlapped. Moreover, most O-GlcNAcylated proteins that already characterized were also detected by Ac36deoGlcNAz and Ac4GalNAz, such as pyruvate kinase PKM [5, 20], glyceraldehyde-3-phosphate dehydrogenase [15, 22] and chromatin assembly factor 1 subunit A [23]. Besides that, Ac36deoGlcNAz proteomic analysis results detected a peptide corresponding to residues 641-654 of DNA (cytosine-5)-methyltransferase 3C (Master protein accession #: P0DOY1) was detected (Fig. 7b), as well as proteins of serine/threonine-protein kinase TAO1 and transcription factor SOX-11 that were the first time identified with O-GlcNAcylation. Of note, 35 and 48 cysteine residues were detected in each, and the ratios of S-glyco-modification were 7.3% and 14.7%, respectively, which suggested Ac36deoGlcNAz induced minimal side-modification compared to Ac4GalNAz (Fig. 7c). These results demonstrated that Ac36deoGlcNAz was an effective and specific MCR for O-GlcNAcylation detection.
In conclusion, we develop a novel MCR of Ac36deoGlcNAz which could effectively label O-GlcNAcylated proteins through GlcNAc salvage pathway in a similar mechanism with Ac36AzGlcNAc [17]. The forming of UDP-6deoGlcNAz can be accepted by OGT but not OGA, which is consistent with our previous results [18]. Furthermore, heterologous expression of NahK in 293T cells could improve the labeling intensity. Both the results of PNGase-F treatment and flow cytometry confirm that Ac36deoGlcNAz does not widely label cell-surface glycoproteins but instead of intracellular proteins. Finally, incubation of Ac36deoGlcNAz with cell lysates could effectively label O-GlcNAcylated proteins without inducing unexpected artificial S-glyco-modification. All the results above exhibit that Ac36deoGlcNAz is an efficient and specific MCR for labeling protein O-GlcNAcylation with no artificial S-glyco-modification, which could be widely applied in the proteomic identification of O-GlcNAc modified proteins.
This manuscript is approved by all authors for publication and there are no conflicts to declare.
This work was supported by the National Natural Science Foundation of China (Nos. 81901622, 31770987, 81971497 and 21907022); Key R & D and Promotion Special Project in Henan Province (Nos. 212102310185, 212102310899); Key Scientific Research Projects in Henan Colleges and Universities (No. 20A350001); Joint Construction Project of Henan Medical Science and Technology Project (No. LHGJ20210566).
Supplementary material associated with this article can be found, in the online version, at doi:
J. Ma, C. Wu, G.W. Hart, Chem. Rev.121 (2021) 1513-1581. doi: 10.1021/acs.chemrev.0c00884
R.S. Haltiwanger, G.D. Holt, G.W. Hart, J. Biol. Chem. 265 (1990) 2563-2568. doi: 10.1016/S0021-9258(19)39838-2
J.A. Hanover, S. Yu, W.B. Lubas, et al., Arch. Biochem. Biophys. 409 (2003) 287-297. doi: 10.1016/S0003-9861(02)00578-7
D.L. Dong, G.W. Hart, J. Biol. Chem. 269 (1994) 19321-19330. doi: 10.1016/S0021-9258(17)32170-1
Y. Wang, J. Liu, X. Jin, et al., Proc. Natl. Acad. Sci. U. S. A. 114 (2017) 13732-13737. doi: 10.1073/pnas.1704145115
V. Fisi, E. Katai, J. Orban, S. Dossena, A. Miseta, T. Nagy, Molecules 23 (2018) 1275. doi: 10.3390/molecules23061275
H. Nie, H. Ju, J. Fan, et al., Nat. Commun. 11 (2020) 36. doi: 10.1038/s41467-019-13601-8
Q. Zhu, X. Cheng, Y. Cheng, et al., Proc. Natl. Acad. Sci. U. S. A. 117 (2020) 7755-7763. doi: 10.1073/pnas.1915582117
Y. Tian, Q. Zhu, Z. Sun, et al., Angew. Chem. Int. Ed. 60 (2021) 26128-26135. doi: 10.1002/anie.202110053
P.S. Banerjee, O. Lagerlof, G.W. Hart, Mol. Aspects Med. 51 (2016) 1-15. doi: 10.1016/j.mam.2016.05.005
J.A. Hanover, W. Chen, M.R. Bond, J. Bioenerg. Biomembr. 50 (2018) 155-173. doi: 10.1007/s10863-018-9751-2
Y. Chen, X. Zhao, H. Wu, Arterioscler. Thromb. Vasc. Biol. 39 (2019) 1911-1924. doi: 10.1161/atvbaha.119.312192
W.B. Dias, G.W. Hart, Mol. Biosyst. 3 (2007) 766-772. doi: 10.1039/b704905f
S. Cecioni, D.J. Vocadlo, Curr. Opin. Chem. Biol. 17 (2013) 719-28. doi: 10.1016/j.cbpa.2013.06.030
M. Boyce, I.S. Carrico, A.S. Ganguli, et al., Proc. Natl. Acad. Sci. U. S. A. 108 (2011) 3141-3146. doi: 10.1073/pnas.1010045108
J. Li, J. Wang, L. Wen, et al., ACS Chem. Biol. 11 (2016) 3002-3006. doi: 10.1021/acschembio.6b00678
K.N. Chuh, B.W. Zaro, F. Piller, J. Am. Chem. Soc. 136 (2014) 12283-95. doi: 10.1021/ja504063c
S. Li, J. Wang, L. Zang, et al., Front. Chem. 6 (2018) 646.
L. Cai, W. Guan, M. Kitaoka, et al., Chem. Commun. (20) (2009) 2944-1946. doi: 10.1039/b904853g
W. Qin, K. Qin, X. Fan, et al., Angew. Chem. Int. Ed. 57 (2018) 1817-1820. doi: 10.1002/anie.201711710
Y. Hao, X.Q. Fan, Y.J. Shi, et al., Nat. Commun. 10 (2019) 4065. doi: 10.1038/s41467-019-11942-y
J.F. Alfaro, C.X. Gong, M.E. Monroe, J.T. Aldrich, T.R. Clauss, Proc. Natl. Acad. Sci. U. S. A. 109 (2012) 7280-7285. doi: 10.1073/pnas.1200425109
Z.H. Wang, N.D. Udeshi, C. Slawson, et al., Sci. Signal. 3 (2010) ra2.

Figure 1 MCRs for O-GlcNAcylation. (a) MCRs that have been reported and the newly synthesized MCR of Ac36deoGlcNAz; (b) The characterization of Ac36deoGlcNAz.

Figure 2 Characterization of newly synthesized MCR of Ac36deoGlcNAz. (a) Ac36deoGlcNAz labeling is concentration dependent. HeLa cells were incubated with Ac36deoGlcNAz at various concentrations for 24 h, then subjected to CuAAC and Western blot analysis; (b) Ac36deoGlcNAz labeling is time dependent. HeLa cells were treated with Ac36deoGlcNAz 200 µmol/L for different length of time, and then subjected to CuAAC and Western blot analysis.

Figure 3 Ac36deoGlcNAz is an O-GlcNAc reporter. (a) Ac36deoGlcNAz labeling is conducted by OGT. 200 µmol/L Ac36deoGlcNAz was metabolic labeling in the presence of OGT inhibitor (OSMI) or OGA inhibitor (Thiamet G), the lysates were subjected to CuAAC and Western blot analysis. (b) The signal of Ac36deoGlcNAz labeled proteins was decreased due to the protein degradation. HeLa cells were treated with 200 µmol/L Ac36deoGlcNAz for 36 h, at which time media was exchanged for fresh media. Cells were harvested after the indicated lengths of time, subjected to CuAAC, and analyzed by Western blot.

Figure 4 Overexpression of NahK enzyme in 293T cells could enhance Ac36deoGlcNAz labeling.

Figure 5 Ac36deoGlcNAz does not chemically modify cell lysates. The indicated concentrations of various MCRs unpretreated (a) or pretreated (b) with iodoacetamide were incubated with cell lysates before CuAAC with Biotin-PEG4-Alkyne and Western blot analysis.

Figure 6 Ac36deoGlcNAz is a MCR for intracellular proteins. (a) The signals labeled with Ac36deoGlcNAz were diminished after treatment with 55 mmol/L NaOH; (b) PNGase F could not reduce the labeling signal from Ac36deoGlcNAz.

Figure 7 Identification of GlcNAc-modification sites in Ac36deoGlcNAz and Ac4GalNAz-treated B16 cells. (a) Schematic illustration of selective enrichment and MS analysis of modified peptides; (b) ETD MS/MS spectrum of the DNA (cytosine-5)-methyltransferase 3C peptide with an O-GlcNAcylation site; (c) The numbers of O-GlcNAc sites modified by two MCRs in B16 cells.

 扫一扫看文章
扫一扫看文章

扫一扫关注我们
 DownLoad:
DownLoad:
 下载:
下载:

 下载:
下载: