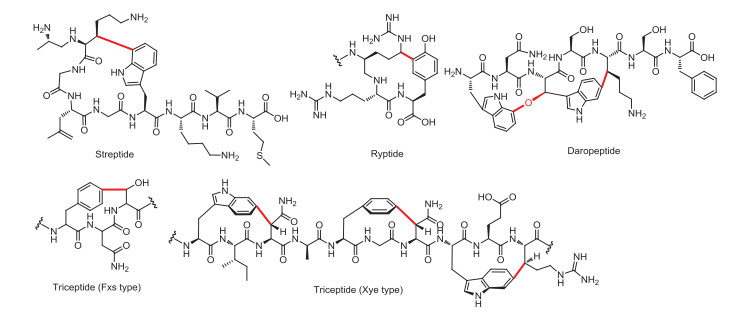
Figure 1.
Chemical structure of selected rSAM-dependent RiPPs. The crosslinks introduced by rSAM enzymes are highlighted in red.

Ribosomally synthesized and post-translationally modified peptides (RiPPs) are a growing superfamily of peptide natural products that are found in all domains of life [1, 2]. These compounds are derived from a gene-encoded precursor peptide and are produced by a series of posttranslational modifications. Owing to the increasingly expanding genome sequences and the vast diversity of the RiPP superfamily, mining of these natural products provided to be fruitful for discovering novel biochemistries as well as compounds with unique structures [1, 2]. Of particular interests in RiPP biosynthesis are radical S-adenosylmethionine (rSAM) enzymes, one of the largest enzyme superfamilies thus far containing more than 700,000 members [3, 4]. rSAM enzymes are extensively involved in RiPP biosynthesis and catalyze strikingly diverse reactions [5-9]. The rSAM-dependent RiPPs include sactipeptide (α-thioether linkage) [10-14], ranthipeptide (β or γ-thioether linkage) [15-17], streptide (C-C crosslink between Lys and Trp) [18], rotapeptide (C-O crosslink between Thr and Gln) [19], ryptide (C-C crosslink between Arg and Tyr) [20], daropeptide (C-O-C crosslink) [21-23], and poly-cyclopropylglycine-containing peptides [24], among others (Fig. 1). In these reactions, the rSAM enzymes utilize a strictly conserved [4Fe-4S] cluster to reductively cleave S-adenosylmethionine (SAM), and the resulting 5′-deoxyadenosyl (dAdo) radical then abstracts a hydrogen atom from the substrate to initiate a variety of oxidation reactions [25, 26].
A growing group of rSAM enzymes involved in RiPP biosynthesis is three-residue cyclophane-forming enzymes (3-CyFEs), which catalyze the crosslinking between an aromatic residue (Ω1) and a non-aromatic residue (X3) in a Ω1-X2-X3 motif to produce a cyclophane moiety (Fig. 1), and the mature RiPPs are designated as triceptide (three-residue in cyclophane peptides) [27]. 3-CyFEs are abundant in diverse bacterial phyla and can be further classified into different clades based on the characteristic motif of the substrate, such as Xye, Grr and Fxs [27, 28]. The Xye subfamily of triceptides is named because their biosynthetic gene clusters are mainly found in X and E. These compounds contain three cyclophane rings, including two Trp-derived rings in the N- and C-termini, and a Phe-derived ring in the middle (Fig. 1) [27]. The precursor peptide XyeA is roughly 50 amino acids long, consisting of a C-terminal core peptide composed of 12–15 amino acids, and an N-terminal leader peptide ended with a characteristic Gly-Gly motif, which is predicted to be the proteolytic cleavage site to remove the leader peptide [29, 30].
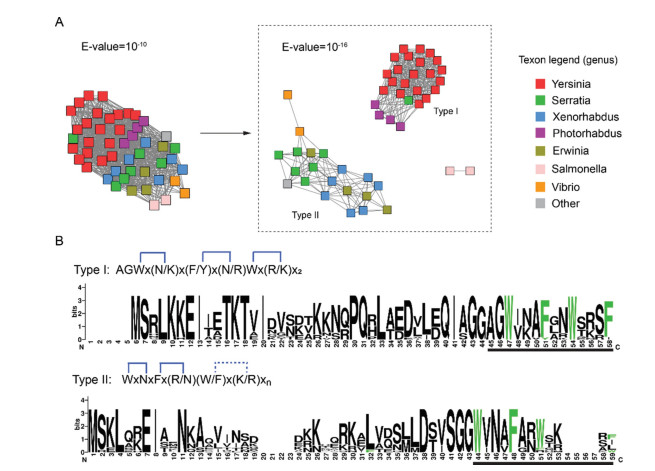
To explore the structural diversity of the Xye-type triceptide and their biosynthetic chemistry, a genome-wide search was carried out in the NCBI database for the biosynthetic gene clusters (BGCs) of the Xye-type triceptide. We performed multiple rounds of position-specific iterated (PSI)-BLAST search, and then a combination of EFI genome neighborhood tool (EFI-GNT) analysis [31] and rapid ORF description & evaluation online (RODEO) analysis [32]. These analyzes revealed 48 unique precursor peptides, which are characteristic of the Xye-type precursors and distinct from those of other triceptide subfamilies such as Grr and Fxs. The identified BGCs are all from γ-proteobacteria, suggesting these RiPPs are likely class-specific. Sequence similarity network (SSN) analysis of the pooled XyeA sequences showed that, in contrast to the hypervariable Fxs precursors shown in our previous analysis [28], Xye precursors are relatively conserved in sequence and can be grouped into two types with a relatively stringent criterion (i.e., E = 10−16) (Fig. 2A). The two types of XyeAs are highly divergent in leader peptide region but share a common LX4LX4GG motif (X denotes any other residues) (Fig. 2B), which is a recognition sequence of peptidase-containing ATP-binding transporter [29]. Type I XyeAs are mainly from the genus Yersinia and Phtorhabdus, and the mature products likely contain an extra N-terminal Ala-Gly motif, which is not present in the product of type Ⅱ XyeA that are mainly from Serratia, Xenorhabdus, Erwinia and Vibrio. Unlike the conserved C-terminal Ser-Phe motif in the type Ⅰ XyeA, the C-terminal sequences in the type Ⅱ precursors are more variable (Fig. 2B). Besides the precursor peptide XyeA and the 3-CyFE XyeB, the Xye BGCs also encodes a HlyD family secretion protein (XyeC), an ABC transporter fused with an N-terminal C39 peptidase domain (XyeD), and an aspartyl protease (XyeE) (Fig. 3A).
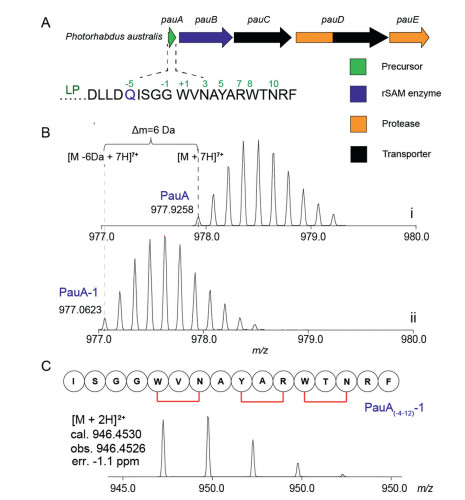
We focus on a type Ⅱ Xye BGC from Photorhabdus australis. To validate the enzyme activity of 3-CyFE encoded by this gene cluster, the precursor peptide PauA was coexpressed with the rSAM enzyme PauB in E. coli. In this analysis, PauA was expressed as a fusion protein, which contains an N-terminal hexa-histidine (His6) tag, a following maltose-binding protein (MBP), and a TEV protease cleavage site that separates MBP from PauA in the C-terminus [33]. To facilitate downstream analysis and characterization, Gln at the -5 position in the leader peptide was mutated to Lys for tryptic digestion (according to the RiPP nomenclature [1, 2], here the core peptide is labeled with positive numbers starting from the first residue of the predicted core sequence, and the leader peptide with negative numbers in a C-to-N order) (Fig. 3A). The resulting fused pauA construct was then coexpressed with pauB in E. coli, and the modified PauA peptide was purified by Ni-affinity chromatography followed by TEV protease digestion to remove the N-terminal His6 and MBP tags. Liquid chromatography with high-resolution mass spectrometry (LC-HRMS) analysis showed the resulting PauA is 6 Da less ([M + 7H]7+ = 977.06) compared to the unmodified PauA obtained by expressing PauA alone ([M + 7H]7+ = 977.93) (Fig. 3B), suggesting three cyclophane crosslinks were formed in PauA. The resulting peptide was then digested by trypsin and analyzed by LC-HRMS, which revealed the expected peptide fragment PauA(-4-12) containing three cyclophane crosslinks (Fig. 3C). Detailed HR-MS/MS clearly showed the three crosslinks are formed, respectively between Trp1 and Asn3, Tyr5 and Arg7, and Trp8 and Asn10, and this result is consistent with the expected 3-residue cyclophane structures of the Xye-type triceptides (Fig. S1 in Supporting information). We hereafter refer to the fully modified PauA (which contains three cyclophane rings) as 1, and the tryptic fragment was accordingly termed PauA(-4-12)-1.
Sequence analysis of the putative Xye precursor peptide showed that the to-be-cyclized X3 residues in the Ω1-X2-X3 motif is relatively conserved, which appear as Asn, Lys or Arg (Fig. 2B). To determine whether the enzyme can also act on other residues, we, respectively changed Asn3, Arg7, and Asn10 to Ser, a natural substrate of the Fxs type 3-CyFEs [27, 28]. The three PauA mutants (i.e., N3S, R7S, N10S) were then co-expressed with PauB, and the resulting peptides were purified, digested by TEV protease, and analyzed in a way similar that of PauA with the wild type core as discussed above. LC-HRMS analysis of the resulting peptides showed that all the three Ser-based mutants were fully modified to the corresponding -6 Da products (Fig. 4A), and this observation is consistent with the formation of the complete set of three cyclophane crosslinks in the three mutants. HR-MS/MS analysis of the corresponding tryptic fragments of the three mutants clearly revealed the crosslink between newly-introduced Ser residues and the corresponding Ω1 residue, besides the other two cyclophane rings (Figs. S2–S4 in Supporting information). We also constructed three Ala-based mutants (i.e., N3A, R7A, N10A), and each of these mutants was coexpressed with PauB followed by purification and proteolytic digestion. Subsequent LC-HRMS and HR-MS/MS analysis revealed that, similar to those of the Ser-based mutants, complete sets of cyclophane crosslinks were formed for all the three Ala-based mutants (Fig. 4B and Figs. S5–S7 in Supporting information). These results indicate that both Ser and Ala are efficient substrates of PauB for cyclophane formation in triceptide biosynthesis.
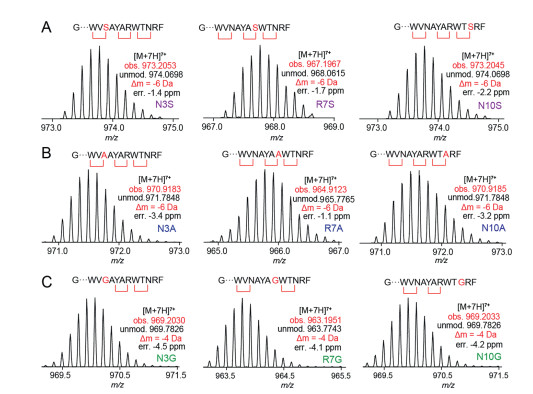
We next set out to test whether biosynthesis of the three cyclophane rings are independent of each other. Because the crosslinks introduced by 3-CyFEs are formed via the β-carbon of X3, we reasoned that changing X3 to Gly (which does not have a β-carbon) would abolish cyclophane formation at the mutation site. This mutagenesis would also impede the formation of other cycylophanes if formation of one cyclophane ring is neccessary for the the latter. We hence generated three Gly-based PauA mutants (i.e., N3G, R7G, N10G) and analyzed these peptides after coexpression with PauB. LC-HRMS analysis showed that only two cyclophanes were formed for all the Gly-based mutants. Following HR-MSMS analysis of the corresponding tryptic fragments of the mutants showed no crosslink is formed at the mutation site, whereas other two cyclophanes were installed succesfully (Fig. 4C and Figs. S8–S10 in Supporting information). These observations indicate that cyclophane produced by 3-CyFE can only be formed via the β carbon of the side chain. Although three cyclophane rings are produced in the precurosr peptide, formation of each crosslink is independent of other two rings.
To further investigate the PauB-catalyzed cyclophane formation, we purified PauB as an N-terminal His6 tagged form, and reconstituted the [4Fe-4S] cluster under a strict anaerobic condition, in a way similar to our previous analysis with other radical SAM enzymes [34-37]. Quantification analysis showed that each enzyme contains 8.2 ± 0.4 iron and 9.1 ± 0.5 labile sulfide, suggesting PauB harbors two [4Fe-4S] clusters. This observation is consistent with the fact that 3-CyFEs contains a C-terminal SPASM/twitch domain, which binds additional [4Fe-4S] clusters, with roles suggestive of peptide binding and/or electron transfer [38, 39]. We then treated the reconsituted PauB with sodium dithionite (DTH), a strong reductant that has been commonly used in converting [4Fe-4S] from the inactive +2 to the active +1 state [26]. Incubation of the reconstituted PauB with DTH and SAM resulted in apparent production of 5′-deoxyadenosine (dAdoH) (Fig. S11 in Supporting information), indicating that PauB is indeed a rSAM protein. In contrast to many other rSAM enzymes that do not require external reductant for activity [40-45], DTH appears to be strictly essential for enzyme activity (Fig. S11). The assay was then performed by incubation of PauA with PauB in the presence of SAM and DTH under strictly anaerobic condition. LC-HRMS analysis of the reaction mixture clearly revealed production of the fully modified PauA (PauA-1) carrying three cyclophanes (Fig. 5A), which was further validated by HR-MSMS analysis (Fig. S12 in Supporting information). We did not observed any signals corresponding to the non-cyclized fragment of the -6 Da product (Fig. S12), suggesting that PauB specifically catalyzed cyclophane formation on PauA, distinct from the remarkable catalytic promiscuity of the Fxs-type 3-CyFE SjiB observed in our previous analysis [28].
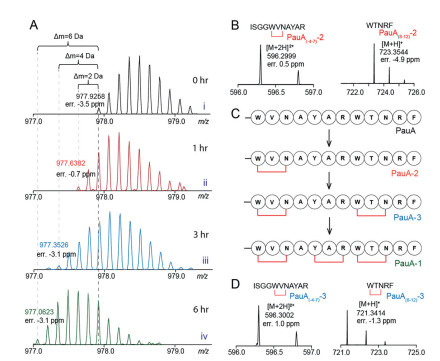
We next interrogated the reaction order in the formation of the three cyclophane rings. To this end, we performed the reaction and monitored the time course of in vitro PauA modification. This analysis showed the main product in 1 h reaction carries only one cyclophane ring (hereafter reffered as to PauA-2) (Fig. 5A, trace ii). The product carrying two cyclophane rings (i.e. PauA-3) culminated around 3 h (Fig. 5A, trace iii), whereas the fully modified product PauA-1 appeared as the major product after 6 h reaction (Fig. 5A, trace iv). These observation strongly indicates that the PauB-catalyzed multiple cyclophane production on PauA is a distributive process, an observation consistent with most (if not all) RiPP biosynthesis enzymes [46].
We then treated the PauA product from 1 h reaction by trypsin and analyzed it by LC-HRMS. Because cyclophane formation between Tyr5 and Arg7 would completely block trypsin digestion, as shown for PauA(-4-12)-1, it is expected that an intact fragment PauA(-4-12) carrying a -2 Da modification would be found if the first cyclophane was formed between Tyr5 and Arg7. However, such a PauA(-4-12) fragment was not observed in our analysis. Instead, the fragment PauA(-4-7) carrying a -2 Da modification (i.e. PauA(-4-7)-2) and the unmodified PauA(8-12) were observed in the assay (Fig. 5B), which has been further validated by HR-MSMS analysis (Figs. S13 and S14 in Supporting information). In contrast, the modified PauA(8-12) fragment carrying a -2 Da modification was barely observed in the reaction. These results clearly demonstrated that the N-terminal cyclophane produced from Trp1 and Asn3 is the first ring produced in PauB reaction (Fig. 5C).
LC-HRMS analysis of the 3 h reaction product pre-treated with trypsin revealed two fragments PauA(-4-7)-3 (which is same to PauA(-4-7)-2) and the PauA(8-12)-3. This two fragments both carry a -2 Da modification (Fig. 5D), which have been further validated by HR-MSMS analysis (Figs. S15 and S16 in Supporting information). In contrast, the intact PauA(-4-12) carrying a -4 Da modification was not found in the assay. These results indicates that the second crosslink catalyzed by PauB is the C-terminal cycphophane produced between Trp8 and Asn11, and the middle cycophane is produced lastly in PauB catalysis (Fig. 5C).
In summary, we show the BGCs of the Xye-type triceptides are present in a variety of gamma-proteobacteria. Although the motifs involved in cylcophane formation have characterstic features, and only three residues (i.e. Asn, Lys and Arg) are found at the X3 position, other residues such as Ser and Ala can also be recognized and modified for cylophane formtion. Although Gly at the X3 position cannot be modified, blocking cyclophane formation by the Gly-based mutagenesis does not have any observable impacts on the production of other two cyclophanes, suggesting that the three cyclophanes in the Xye system are produced independently. We also reconsituted the in vitro activity of PauB, showing it is fully active to install the complete set of cyclophane rings on PauA. Moreover, we showed in PauB catalysis, the N-terminal cyclophane is produced firstly, which is followed by the formation of the C-terminal ring, whereas the middle cyclophane formed from Phe5 and Arg7 is produced in the last step (Fig. 5C). Our study paves the way for future biosynthetic and engineering study of triceptides, and also contributes to the rapidly expanding knowledge the rSAM superfamily enzymes and the rSAM-dependent RiPP natural products.
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
This work is supported by grants from the National Key Research and Development Program (Nos. 2018YFA0900402 and 2021YFA0910501), from the National Natural Science Foundation of China (Nos. 21822703, 21921003, and 32070050), from the funding of Innovative research team of high-level local universities in Shanghai and a key laboratory program of the Education Commission of Shanghai Municipality (No. ZDSYS14005), and from West Light Foundation of the Chinese Academy of Sciences (No. xbzg-zdsys-202105).
Supplementary material associated with this article can be found, in the online version, at doi:10.1016/j.cclet.2022.06.012.
P.G. Arnison, M.J. Bibb, G. Bierbaum, et al., Nat. Prod. Rep. 30 (2013) 108-160. doi: 10.1039/C2NP20085F
M. Montalbán-López, T.A. Scott, S. Ramesh, et al., Nat. Prod. Rep. 38 (2021) 130-239. doi: 10.1039/d0np00027b
R. Zallot, N. Oberg, J.A. Gerlt, Biochemistry 58 (2019) 4169-4182. doi: 10.1021/acs.biochem.9b00735
N. Oberg, T.W. Precord, D.A. Mitchell, J.A. Gerlt, ACS Bio Med Chem. Au 2 (2022) 22-35. doi: 10.1021/acsbiomedchemau.1c00048
D.H. Haft, M.K. Basu, J. Bacteriol. 193 (2011) 2745-2755. doi: 10.1128/JB.00040-11
N. Mahanta, G.A. Hudson, D.A. Mitchell, Biochemistry 56 (2017) 5229-5244. doi: 10.1021/acs.biochem.7b00771
A. Benjdia, C. Balty, O. Berteau, Front. Chem. 5 (2017) 87. doi: 10.3389/fchem.2017.00087
K.A. Clark, L.B. Bushin, M.R. Seyedsayamdost, ACS Bio Med Chem Au 2 (2022) 328–339. doi: 10.1021/acsbiomedchemau.2c00004
A. Mendauletova, A. Kostenko, Y. Lien, J. Latham, ACS Bio Med Chem Au 2 (2022) 53-59. doi: 10.1021/acsbiomedchemau.1c00045
T. Mo, X. Ji, W. Yuan, et al., Angew. Chem. Int. Ed. 58 (2019) 18793-18797. doi: 10.1002/anie.201908490
L.B. Bushin, B.C. Covington, B.E. Rued, M.J. Federle, M.R. Seyedsayamdost, J. Am. Chem. Soc. 142 (2020) 16265–16275. doi: 10.1021/jacs.0c05546
S. Chiumento, C. Roblin, S. Kieffer-Jaquinod, et al., Sci. Adv. 5 (2019) eaaw9969. doi: 10.1126/sciadv.aaw9969
C. Balty, A. Guillot, L. Fradale, et al., J. Biol. Chem. 294 (2019) 14512-14525. doi: 10.1074/jbc.ra119.009416
C. Roblin, S. Chiumento, O. Bornet, et al., Proc. Natl. Acad. Sci. USA. 117 (2020) 19168-19177. doi: 10.1073/pnas.2004045117
G.A. Hudson, B.J. Burkhart, A.J. DiCaprio, et al., J. Am. Chem. Soc. 141 (2019) 8228-8238. doi: 10.1021/jacs.9b01519
Y. Chen, Y. Yang, X. Ji, et al., J. Biotechnol. (2020) e2000136.
A. Caruso, L.B. Bushin, K.A. Clark, R.J. Martinie, M.R. Seyedsayamdost, J. Am. Chem. Soc. 141 (2019) 990-997. doi: 10.1021/jacs.8b11060
K.R. Schramma, L.B. Bushin, M.R. Seyedsayamdost, Nat. Chem. 7 (2015) 431-437. doi: 10.1038/nchem.2237
K.A. Clark, L.B. Bushin, M.R. Seyedsayamdost, J. Am. Chem. Soc. 141 (2019) 10610-10615. doi: 10.1021/jacs.9b05151
A. Caruso, R.J. Martinie, L.B. Bushin, M.R. Seyedsayamdost, J. Am. Chem. Soc. 141 (2019) 16610-16614. doi: 10.1021/jacs.9b09210
Y. Imai, K.J. Meyer, A. Iinishi, et al., Nature 576 (2019) 459-464. doi: 10.1038/s41586-019-1791-1
S. Groß, F. Panter, D. Pogorevc, et al., Chem. Sci. 12 (2021) 11882-11893. doi: 10.1039/d1sc02725e
S. Guo, S. Wang, S. Ma, et al., Nat. Commun. 13 (2022) 2361. doi: 10.1038/s41467-022-30084-2
A. Kostenko, Y. Lien, A. Mendauletova, et al., J. Biol. Chem. 298 (2022) 101881. doi: 10.1016/j.jbc.2022.101881
H.J. Sofia, G. Chen, B.G. Hetzler, J.F. Reyes-Spindola, N.E. Miller, Nucleic Acid. Res. 29 (2001) 1097-1106. doi: 10.1093/nar/29.5.1097
W.E. Broderick, B.M. Hoffman, J.B. Broderick, Acc. Chem. Res. 51 (2018) 2611-2619. doi: 10.1021/acs.accounts.8b00356
T.Q.N. Nguyen, Y.W. Tooh, R. Sugiyama, et al., Nat. Chem. 12 (2020) 1042–1053. doi: 10.1038/s41557-020-0519-z
S. Ma, H. Chen, H. Li, et al., Angew. Chem. Int. Ed. 60 (2021) 19957-19964. doi: 10.1002/anie.202107192
S.C. Bobeica, S.H. Dong, L. Huo, et al., eLife 8 (2019) e42305. doi: 10.7554/eLife.42305
T.J. Oman, W.A. van der Donk, Nat. Chem. Biol. 6 (2010) 9-18. doi: 10.1038/nchembio.286
J.A. Gerlt, J.T. Bouvier, D.B. Davidson, et al., Biochim. Biophys. Acta 1854 (2015) 1019-1037. doi: 10.1016/j.bbapap.2015.04.015
J.I. Tietz, C.J. Schwalen, P.S. Patel, et al., Nat. Chem. Biol. 13 (2017) 470-478. doi: 10.1038/nchembio.2319
M. Lv, X. Ji, J. Zhao, et al., J. Am. Chem. Soc. 138 (2016) 6427-6435. doi: 10.1021/jacs.6b02221
Y. Zhong, X. Ji, Chin. J. Chem. 38 (2020) 218-219. doi: 10.1002/cjoc.201900480
Y. Zhong, X. Ji, Q. Zhang, Chin. J. Chem. 38 (2020) 39-42. doi: 10.1002/cjoc.201900399
Y. Yin, X. Ji, Q. Zhang, Chin. J. Chem. 39 (2021) 2417-2421. doi: 10.1002/cjoc.202100304
W. Ji, X. Ji, Q. Zhang, et al., Angew. Chem. Int. Ed. 59 (2020) 8880-8884. doi: 10.1002/anie.202000812
T.A. Grell, P.J. Goldman, C.L. Drennan, J. Biol. Chem. 290 (2015) 3964-3971. doi: 10.1074/jbc.R114.581249
N.D. Lanz, S.J. Booker, Biochim. Biophys. Acta 1853 (2015) 1316-1334. doi: 10.1016/j.bbamcr.2015.01.002
M.W. Ruszczycky, S.H. Choi, H.W. Liu, J. Am. Chem. Soc. 132 (2010) 2359-2369. doi: 10.1021/ja909451a
X. Ji, T. Mo, W.Q. Liu, et al., Angew. Chem. Int. Ed. 58 (2019) 6235-6238. doi: 10.1002/anie.201814708
J. Cheng, W. Ji, S. Ma, et al., Angew. Chem. Int. Ed. 60 (2021) 7570-7575. doi: 10.1002/anie.202015177
J. Cheng, W. Ding, Q. Zhang, Chin. J. Chem. 40 (2022) 1053-1058. doi: 10.1002/cjoc.202200032
J. Zhao, W. Ji, X. Ji, Q. Zhang, Chin. J. Chem. 38 (2020) 959-962. doi: 10.1002/cjoc.202000119
R. Wu, W. Ding, Q. Zhang, Chin. J. Chem. 38 (2020) 959–962. doi: 10.1002/cjoc.202000119
H. Lee, W.A. van der Donk, Annu. Rev. Biochem. 91 (2022) 269–294. doi: 10.1146/annurev-biochem-032620-104956

Figure 1 Chemical structure of selected rSAM-dependent RiPPs. The crosslinks introduced by rSAM enzymes are highlighted in red.

Figure 2 Sequence diversity of the Xye-type tricepeptide. (A) Sequence similarity network (SSN) of 48 XyeA sequences. (B) Sequence logos of the two XyeA subgroups.

Figure 3 Triceptide biosynthesis in Photorhabdus australis. (A) The pau BGC encodes a precursor peptide PauA, a 3-CyFE PauB, a HlyD family secretion protein PauC, an ABC transporter fused with a C39 peptidase PauD, and an aspartyl protease PauE. (B) HR-MS characterization of (i) PauA expressed alone and (ii) PauA modified by PauB in vivo. PauA with -6 Da modification (PauA-1) is a result of the formation of three cyclophane rings. (C) HR-MS spectrum of the tryptic fragment PauA(-4-12), which has three cyclophane rings formed between W1 and N3, Y5 and R7, and W8 and N10, respectively. LP denotes leader peptide.

Figure 4 HR-MS analysis of PauA mutants modified by PauB, showing the HR-MS spectra of (A) three Ser-based mutants, (B) three Ala-based mutants, and (C) three Gly-based mutants. The molecular weights were calculated based on the unmodified products, and -6 Da and -4 Da correspond to the formation of three and two cyclophane rings, respectively.

Figure 5 In vitro reconstitution of cyclophane formation by PauB. (A) Time-dependent formation of the three cyclophane rings on PauA. (B) The HR-MS spectra of the tryptic fragments of PauA-2 obtained by treating the 1 h reaction mixture with trypsin. (C) The proposed reaction order of the PauB-catalyzed cyclophane formation on PauA. (D) The HR-MS spectra of the tryptic fragments the PauA-3 obtained by treating the 3 h reaction mixture with trypsin.

 扫一扫看文章
扫一扫看文章

扫一扫关注我们
 DownLoad:
DownLoad:
 下载:
下载:
 下载:
下载: