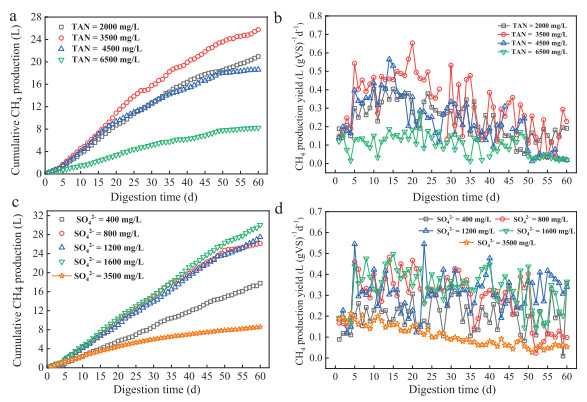
Figure 1.
Methane production (a, c) and variations of methane production yield (b, d) of anaerobic digestion reactors under different ammonia/sulfate concentrations.
Inhibition mechanisms of ammonia and sulfate in high-solids anaerobic digesters for food waste treatment: Microbial community and element distributions responses
Likui Feng , Weixin Zhao , Yu Liu , Yan Chen , Shufei He , Jing Ding , Qingliang Zhao , Liangliang Wei

With the continuous urbanization and the improvement of the living standards of China, the solid waste generation has been reported with an unprecedented increase in recent years [1]. Generally, organic solid waste occupies approximate 50% of the bulk municipal solid waste, in which the food waste, as a major component of the organic solid waste, contributes as high as 30% of the municipal solid waste in China [2,3]. Improper treatment of the food waste may potentially cause serious environmental pollutions such as soil and underground water pollution [3]. Tremendous approaches, such as direct feeding to pigs, composting, landfilling, etc., have been widely applied for the food waste treatment [4,5]. However, some serious environmental issues, high disposal cost and huge demanding of the land space seriously restricted their application [6]. Therefore, there is a pressing need to explore alternative methods of food waste for resourceful and security processing.
Anaerobic digestion (AD), an environment-friendly technology, has recently been developed to treat organic waste due to its environmental benefits, such as renewable energy production, soil fertilizer generation and small land requirements [7,8]. However, the AD systems always suffer from instable operation during the high solid waste treatment with the total solid (TS) concentration > 15%, especially for food waste with high proteins [9]. Additionally, the recent work of Serna-Maza et al. [10] stated that the abundant existence of protein with the food waste would negatively lead to a huge releasing of the ammonia nitrogen (NH4+-N) during its degradation, and undoubtedly high levels of ammonia will restrain the AD process and influence the performance of gas production. Many approaches including air stripping, bioaugmentation and ammonia-binding have been proposed to counteract ammonia inhibition in AD process [9,11]. However, high operational cost and technical challenges associated with these approaches further hinder their full-scale practical application [12].
Tremendous researches have been focused on the clarifying the threshold values of ammonia inhibition with different substrates. For example, Poirier et al. [13] found that an ammonia concentration of 19 g/L would suppress the AD of biowaste, while 800-1400 mg/L ammonia in swine slurry would inhibit the methane (CH4) production [14]. It was also reported that AD of food waste would be severely inhibited by free ammonia (FAN) at 300 mg/L [9]. However, few studies have reported the time and concentrations of ammonia inhibition occurrence in the horizontal flow anaerobic digester or investigated the inhibition mechanism based on real concentrations. Besides ammonia, sulfate (SO42−) has been regarded as another main limited factor influencing the gas production in AD process, despite the sulfur accounts only 0.2%-0.6% of the total mass of the food waste [15]. Sulfate reducing bacteria (SRB) in AD system take precedence in using carbon for electron donors than methanogenic archaea, thus SRB can compete with syntrophic bacteriam for propionate, butyrate and other short-chain volatile fatty acids [16]. Additionally, hydrogen sulfide (H2S) produced from SRB can also restrain methanogens [17]. Therefore, understanding the negative effect of sulfate onto the anaerobic digester is essential for enhancing the performance of AD systems.
Investigation of the transformation of elements of N and C in different operation phases of AD systems not only can clarify the degradation characteristics of the different pollutants, but also can improve the performance of AD systems from the view of elemental cycles. Li et al. [18] found that conversion efficiency of C and nutrient in liquid/solid stream of swine manure substrate, during anaerobic digestion, were higher than that using cattle manure and straws. Alcántara et al. [19] quantified the mass balance of C, N and P in the integrated process of microalgae cultivation coupled with AD to achieve stable operation and reused organics and nutrients in supernatants/residues according to the specific distribution of C and N. However, seldom works have been reported the distribution of C and N in AD process under the condition of ammonia/sulfate inhibition, which will provide new sights into an improved understanding of AD with food waste substrates.
The main objectives of this work were to: (1) Evaluate the methane productivity of horizontal flow anaerobic digester, using food waste as substrates, and determine the time and concentrations of inhibition occurrence; (2) explore the inhibition mechanisms of ammonia/sulfate in AD systems from the perspective of C and N transformation; and (3) investigate the microbial community distribution trend under different ammonia/sulfate conditions for further understanding the inhibition mechanism.
The schematic diagram of horizontal flow anaerobic digester was made from polymethyl methacrylate. The concrete design was described in Text S1 (Supporting information). Bottle anaerobic digestion reactors were conducted using anaerobic digestion methanogenic potential measurement systems (MC-BMP-Ⅱ). The system was mainly consisted of three parts: water bath, detector and 500-mL flasks. First, 200 mL substrates of food waste were added into flaks, and sealed with lids, which connected with the detector via pipes and special line. Then, those digesters were placed in the bath water under a constant temperature of 37 ± 1 ℃. And the detector would record the accumulative methane production and rate every minute.
Artificial food waste, main ingredients similar to the food waste in China, were prepared and applied as the substrates of the AD reactors according to Rajagopal et al. (Table S1 in Supporting information) [20]. Specifically, those configurated food were crushed an average diameter of 3 mm using a vegetable grinder before feeding. The inoculated sludge with the values of TS (3.14%), volatile solid (VS, 2.43%) and pH (7.24) in the horizontal flow anaerobic digestion reactor was taken from Dalian Dongtai Xiajiahe sludge treatment plant. The inoculated sludge (TS = 14.42%, VS = 7.51%, pH 7.92) of the bottle systems was taken from the horizontal flow anaerobic digester. To consume the internal organic matter, all collected sludge was placed in the laboratory at room temperature under anaerobic environment for 3 days (d). The other parameters of inoculative sludge were listed in Table S2 (Supporting information).
The horizontal flow digester was operated under a semi-continuous mode, with an organic loading was 5.87 kg VS m−3 d−1. The daily feeding/discharge food waste was about 220 g, and 50 mL samples were taken every 2 d to determine the common parameters (Text S2 in Supporting information). Bottle anaerobic digestion tests were designed and operated under four different ammonia nitrogen (NH4+-N) (2000, 3500, 4500 and 6500 mg/L) and five sulfate (400, 800, 1200, 1600 and 3500 mg/L) concentrations, based on the results of horizontal flow anaerobic digester. The different reactors, with varied NH4+-N concentrations, were defined as TAN2000, TAN3500, TAN4500 and TAN6500. Similarly, the AD systems with different SO42− concentrations were labeled as S1 to S5 correspondingly. Semi-continuous operation was applied for the anaerobic systems, with the daily feeding/discharged substrates of 8.3 g via the pipes. The TS of all reactors was kept under a constant value of 15%. All the AD reactors were continuously operated for 60 d, with an average organic loading of 8.85 kg VS m−3 d−1. The samples were taken and measured every two days.
The volume of CH4 was measured by the drainage method, and then, the contents of collected CH4 were determined by the gas detectors (MS600 CH4 detector) every day. Digestate sludge (25 mL) obtained from the reactors were separately centrifuged at 10000 r/min for 20 min. The collected supernatants were filtered using 0.45 µm membrane and stored at 4 ℃. Chemical oxygen demand (COD), soluble COD (SCOD) and total ammonium nitrogen (TAN) within the supernatants were analyzed according to the APHA Standard Method [21]. SO42− concentrations were measured using ion chromatography (Integrion HPIC). The protein content was detected via the Lowry method, whereas volatile fatty acids (VFAs) were measured by gas chromatography (Agilent 6890). The total alkalinity (TA) was conducted with total alkalinity kits. The pH value was determined by a pH meter. Specially, FAN was calculated according to the study of Peng et al. [9]. Additionally, methane, acetic acid, propionic acid, butyric acid and pentyl acid were expressed as the quality of COD, and calculated in Text S3 (Supporting information).
In this study, the biomass within the AD systems of the horizontal flow digester, TAN2000, TAN3500, TAN6500 and S1-S4 were selected for a purpose of microbial community analyzing. The methods of extraction and analysis were conducted based on the study of Peng et al. [9] in Text S4 (Supporting information).
Fig. S1 (Supporting information) shows the variations of parameters in the horizontal flow digester during the whole running period. From 0 d to 18 d, the maximum methane production was observed on 2nd day with 6.86 L, and then sharply decreased to 0 L. Meanwhile, the TAN concentration waved obviously with the maximum concentrations of 2373.95 mg/L, which was mainly caused by the bioconversion of protein existing in food waste into amnio acids and then ammonia [22].
Rajagopal et al. [23] have reported that ammonia inhibition might happen as a result of TAN levels up to 1500-7000 mg/L. Combined the methane production, the horizontal flow digester was considered to suffer from ammonia inhibition as evidenced by its high TAN concentration. To overcome ammonia inhibition, biochar was added into the digester from 18 d to 36 d due to its diverse physicochemical characteristics, which have been confirmed to effectively adsorb inhibitor, and relieve acidification [24]. Obviously, the addition of biochar sharply decreased TAN (1993.15 mg/L) and total VFA (TVFA, 675.48 mg/L) concentrations due to the strong adsorption of biochar. However, the TAN concentration was still higher than the threshold of ammonia concentration related to ammonia inhibition [23], which caused methane production close to 0 L.
According the previous work of Karlsson and Ejlertsson [25], totally 6 mL of HCl (5 mol/L) was added into the horizontal flow digester, during the operational periods of 36-54 d, to shift the NH4+/NH3 ratio of the digester. With the addition of HCl, the TAN concentration showed an obvious declined trend, and the concentrations (944.21 mg/L) was in the acceptable range for AD as reported in previous studies [23,26]. The result indicated the addition of HCl promoted the ionization equilibrium between NH4+ and NH3, which further alleviated ammonia inhibition. Karlsson and Ejlertsson [25] found that the HCl addition decreased the acetate, propionate, isovalerate and other fatty acids, promoted the balance between acidogenesis and methanogenesis. The relatively stable TVFA concentration in Fig. S1 during this period suggested that the dynamic balance between acids formation and consumption has been reached. Nevertheless, the collected methane was not improved with the addition of HCl, implying that ammonia inhibition was not the main factor in this period.
Based on the study of Park et al. [27], dilution of feeding streams was applied to adjust TS from 21.7% to 14.1%, which successfully run the digester. The methane production increased obviously with maximum volume of 21.69 L. However, the methane production displayed a large fluctuation trend, accompanying with the big waves of TAN, TVFA and SO42− concentrations. On 80th day, the SO42− concentration reached 3653.8 mg/L, while the methane production was only 9.94 L. With the reduction of SO42−, the methane production gradually increased. Additionally, the TAN concentration changed fiercely, which seriously influenced the methane production. Hence, there is a pressing need to explore the inhibition mechanism based on the results of the horizontal flow digester.
To further investigate the inhibition mechanism, the bottle digesters under different ammonia/sulfate concentrations were conducted, and the methane production and yields are shown in Fig. 1. Specifically, the TAN3500 system exhibited a relative higher substrates degradation rate, as well as the highest methane volume of 25.74 L (average methane yield 0.32 L (g VS)−1 d−1 among the four tests. A similar biogas production trend has been also observed for the TAN2000 and TAN4500 reactors, with an accumulative methane production of 20.96 L and 18.61 L (corresponding to methane yield were 0.23 and 0.22 L (g VS)−1 d−1, respectively. However, the results were much lower than the previous study, of which the methane production was 0.35-0.45 L (g VS)−1 d−1 with FW as substrates [5], which might result from the operational parameter. Notably, the pH value of the four different reactors changed slightly with a range of 7.8-8.2 (Fig. S2 in Supporting information), owing to high alkalinity caused by the abundant existence of NH4+-N, which was benefited for the biogas production and the operation of AD systems [11]. However, the further increase of the TAN (6500 mg/L) caused progressively lower methane production, which could be proven by the complete inhibition of biogas production with volume of 8.23 L, declining 68.03% as compared to the TAN3500. The inhibition value was much higher than the previous study [13,14], which reported that the inhibitory concentration values were 19.0 g/L in AD of biowaste and 1400 mg/L of swine slurry. The big discrepancy suggested that different substrates and operational parameters would significantly affect the performance of AD. Additionally, the fluctuated CH4 production yields as well as the lower TVFA and alkalinity (TA) ratio (TVFA/TA < 0.4) (Fig. S2) [28] in TAN3500 and TAN4500 implied that an unstable operation occurred in the latter period (Text S5 in Supporting information).
Methane productivity of the AD reactors under different initial concentrations of sulfate are listed in Figs. 1c and d. Generally, the S2, S3 and S4 reactors, performed similarly and exhibited a stable increasing of methane production during the whole operation, with a final CH4 volume of 26.15 L, 27.50 L and 30.05 L, respectively. By contrast, a relatively lower methane production of 8.67 L was observed for S5, indicating that a high sulfate existence inhibited the AD process. The S2, S3 and S4 reactors displayed higher methane yield of 0.29, 0.31 and 0.33 L (g VS)−1 d−1 during the whole digestion, 37.38%, 29.06% and 41.00% higher than that of S1 reactors, suggesting that a significant promotion of methane production could be achieved via sulfate concentration increasing in a certain degree. Thus, we can speculate that the methanogens and SRB within the digesters exhibited a good synergistic relationship for methane production under a sulfate concentration of 800-1600 mg/L. Those bacteria would be inclined to generate the volatile fatty acids (VFAs) via fermentation to adapt environment, whereas an opposite trend of SRB (kept high alkalinity status of the reactors) via activity increasing of the monovalent cation/proton antiporters for capture protons (constant alkaline pH value in Fig. S2), might be the main reason for the enhancement of the biogas generation [29].
However, with the continuous increasing of sulfate concentrations, the competition between methanogens and SRB in carbon source led to a low volatile acid consumption and proton capturing, ultimately an unstable operation of AD reactors [16]. Additionally, H2S produced from SRB also exhibited a significant inhibitory effect on methanogenic species, which hindered the methane generation process [17]. Furthermore, the TVFA/TA value of the S5 reactors was greater than 0.4 (Fig. S2) since the 10th day digestion, clarifying that the lowest CH4 production in S5 was attributed to the unstable running.
Concentration distribution of VFAs, the main substrates for methanogens [30], under different ammonia concentrations are listed in Fig. 2. Briefly, the TVFA concentrations of TAN2000 and TAN3500 systems fluctuated slightly during the initial 42 d operation, indicating that these systems were under a dynamic balance among the hydrolysis, acidification and methanation. Interestingly, the TVFA kept in a low level in TAN2000 system, revealing that the low CH4 productivity was mainly caused by inefficient hydrolyzation [5]. By contrast, the VFAs increased sharply in TAN3500, as evidenced by a relatively higher VFAs concentration of 6727.28 mg/L after 53 d digestion, suggesting that the methanation was inhibited under such ammonia condition. The accumulation of propionic acid, which could not be utilized directly by methanogens [30] in TVFAs (76.61%) demonstrated that the slow conversion of propionic acid to acetic acid might be responsible for the inhibition. Concentration of the TVFAs in TAN4500 and TAN6500 systems reached its maximum value after 26 d continuous digestion, and then a large amount of TVFAs accumulated due to the insufficient utilization.
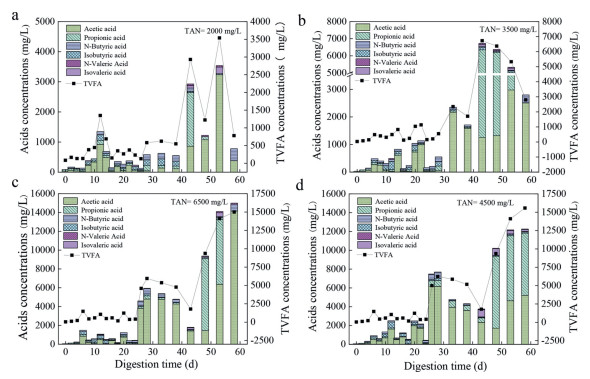
During the subsequential 41-60 d digestion, a relatively higher TVFAs values of 11880 and 15572 mg/L were observed in the TAN2000 and TAN3500 systems. Yang et al. [30] reported that the inhibition would occur once the bulk concentration of VFAs reached 4000 mg/L in the anaerobic digesters. Hence, the obvious accumulation of TVFAs accompanied with the increase of propionic acid, manifested the slow degradation of propionic acid was the limiting factor for AD process, similar to the observation of Hussain and Dubey [31]. Moreover, the concentration of the butyric and valeric acids was quite low during the whole operational periods, regardless of TAN2000 and TAN3500, implying that these acids could be rapidly consumed during the digestion process, consistent with the report of Li et al. [32]. Peng et al. [9] reported that accumulated acetate caused feedback inhibition at the acetogenesis stage, which led to considerable accumulation of propionate, valerate, and other long-chain fatty acids. The similar phenomenon in TAN6500 further indicated the complete inhibition of ammonia would occur once the concentration of ammonia reached 6500 mg/L.
For the anaerobic digesters with different sulfate additive, the generation and accumulation trend of the TVFAs were shown in Fig. S3 (Supporting information). Insufficient organics supplying caused the low CH4 productivity in S1 reactors, accompanying with low VFAs accumulation. Inversely, the AD process of S5 was inhibited at the beginning of the digestion, as evidenced by huge amount of TVFAs accumulation, indicating that high sulfate would affect the methanation process. Li et al. [32] revealed that the acetic acid was the primary factor for acid inhibition during the food waste degradation. The similar phenomenon with the higher acetic acid (above 5000 mg/L) in this study further demonstrated that sulfate inhibition occurred via accumulating acetic acid. Additionally, previous study revealed that high propionic acid would exert toxicity to microorganisms via diffusion of unionized propionic acid into microbial cells [33]. Therefore, a large amount of propionic acid at the latter period of digestion also contributed a lot to sulfate inhibition in S5 reactors (Text S6 in Supporting information).
The percentage distribution of the carbon within the different phases are listed in Table S3 (Supporting information). Experimental results demonstrated that majority of the clement C were existed as the solid phase (around 58.64%) within the AD reactors, regardless of the operational characteristics. Additionally, about 20.87%-26.67% of the element C were as the liquid phase in all TAN systems, clearly demonstrating that the ammonia concentration had no significant effect on the dissolution process of carbon elements from solid to liquid phase. Above results were consistent with the study of Li et al. [18], who found that the element C remained in solid form was over 60.3%. However, the efficiency of C's conversion from the liquid phase into CH4 (2.68%-7.08%) was a bit lower than that of 9.5%-15.8% observed by Li et al. [18]. The different substrates might cause this big discrepancy. By contrast, a relative lower percentage distribution of CH4-Gas in TAN6500 (2.68%) systems meant that higher ammonia inhibited microbial activities and further affected the methanation process [22]. Meanwhile, a higher CH4 conservation rate of TAN3500 (CH4-Gas = 7.08%) than TAN4500 (CH4-Gas = 4.79%) revealed the negative impact on AD process would occur when ammonia concentration reached 4500 mg/L. Rajagopal et al. [23] stated that the TAN concentrations of 1500-7000 mg/L would seriously inhibit the process of methanogenesis, much widely than the observation of the high solid digestion in this work, might be related to the differences in nature of inocula, substrates, and environmental conditions [34].
To intuitively investigate the element C transformation from liquid to gas phase, the contents of acetic acid, propionic acid and CH4 within the AD reactors were unified to COD (g), and then SCOD was applied in Fig. S4 (Supporting information). The contents of SCOD in all systems exhibited a noteworthy increment during the initial 22 d operation, revealing that the majority of the organics within the food waste was gradually degraded into soluble organic matters. During this period, the bio-conservation rate of CODmethane were much higher than that of CODacetic acid and CODpropionic acid in TAN2000 and TAN3500 systems, which reflected that the methanation was not a limiting factor under such ammonia concentrations. It is noteworthy that the parameters of CODmethane, CODacetic acid and CODpropionic acid all exhibited a significant declining within the TAN4500 and TAN6500, while the SCOD value increased and reached 31.56 g/L and 30.72 g/L, respectively, which implied that the inhibition occurred in the organic carbonation pathways both for hydrolysis and methanation. For the subsequential stage operation (from 22th day to 60th day), despite the CODacetic acid and CODpropionic acid contents were higher than that CODmethane in TAN2000 and TAN3500 systems, a relative higher CODmethane conservation (> 6.3%) revealed an efficient CH4 productivity. Notably, the CODpropionic acid contents increased sharply in TAN4500 and TAN6500 systems during digestion periods (> 7.0 g/L). Yang et al. [30] pointed that propionic acid accumulation could restrict the AD process and even jeopardize their performance; thus, the accumulative propionic acid was responsible for lower CH4 productivity. Similar to TAN systems, majority of the element C remained in solid phase in the AD reactors of S1-S5 (> 60.3%), followed by liquid phase (below 13.7%). Meanwhile, the variations of SCOD, CODmethane, CODacetic acid and CODpropionic acid under different sulfate concentrations were displayed in Fig. S5 and discussed in Text S7 (Supporting information).
For element N, ammonia concentration distribution within the substrates had exhibited no significant effect on the solid- and liquid-phase distribution of nitrogen, as evidenced by the insignificant variation of the percentage distribution of TN-solid (75.80%-80.43%) or TN-liquid (average 12.61%-16.62%) within the TAN systems (Table S4 in Supporting information). Similar to that of the element C, majority of the nitrogen were existed as solid phase, consistent with the observation of Li et al. [35], who reported that the distributions of solid-related and liquid-related TN were 79.3% and 18.1%, respectively, for the cattle mature substrates. As shown in Fig. 3a, the concentrations of the soluble protein were all exhibited the same distribution trend that increased during the initial stage operation and then continuously decreased during the subsequential stage. Above results demonstrated that the hydrolyzation of the solid protein from food waste into the liquid phase in the initial stage operation, as well as the deterioration of the microbial cell wall and the releasing of the intracellular materials led to the concentration rising of the soluble protein [36].
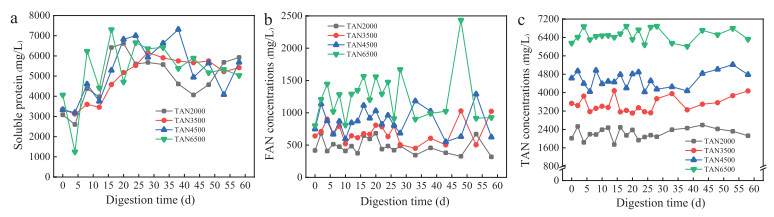
The FAN has been regarded as the main reason for the suppression of the AD systems due to its high permeability for bacterial cell membrane [6]. As shown in Fig. 3b, the most significant accumulation of FAN has been detected in TAN6500 system, followed by TAN4500 and TAN3500, whereas that of the TAN2000 was the lowest; and the accumulation trend became more significant with the increasing of the digestion time. Rajagopal et al. [23] revealed that abundant existence of the FAN within the AD system not only inhibited the growth of the methanogenic archaea, but also toxic to the acetogenic bacteria related to propionate degradation. That might be the main reason why TAN6500 system were inhibited by FAN (801.54 mg/L) at initial period. Previous studies stated that decreased pH could alleviate FAN inhibition via changing of the ionization equilibrium between NH4+ and NH3 [7,9]. However, the stable pH and large fluctuation of FAN concentrations in TAN4500 at the latter period implied that these digesters began to be in inhibited state, which was consistent with the decreasing methane production rates in Fig. 1b.
Concentration of the TAN of each system displayed an obvious gradient with the artificially regulation of ammonia, with average concentrations of 2125, 4074, 4784 and 6322 mg/L in TAN2000, TAN3500, TAN4500 and TAN6500 during the steady-state operation of the AD systems (Fig. 3c). Sun et al. [26] revealed that TAN inhibition firstly led to the concentration rising of the acetate and subsequently fell once the propionate accumulated. Similar results were also observed in TAN4500 and TAN6500 systems (Figs. 1c and d), indicating the occurrence of TAN inhibitory in above two AD systems. It was worthwhile to point that an obvious lag phase was observed for TAN4500, which revealed a real TAN inhibiton might occur at 4500-4784 mg/L. The result was consistent with the reported inhibitory concentration of TAN at 1500-7000 mg/L in AD systems. The similar phenomenon was also observed in sulfate systems (Fig. S6 in Supporting information) and were discussed in Text S8 (Supporting information).
The diversity and richness of microbial community were listed in Table S4 and discussed in Text S9 (Supporting information). The results manifested that high ammonia/sulfate showed toxicity to archaea and bacteria, which influenced the diversity and richness of microbial community [37]. To further investigate the microbial structures, the circuses of bacteria at genus level in different digestion systems are listed in Fig. 4a. Methanosarcina, Methanobacterium, and Methanoculleus were the predominant archaea within the digesters. Gonzalez et al. [14] pointed that Methanosarcina, belonging to aceticlastic methanogens, could utilize acetate for methane production. Therefore, the highest relative abundance of Methanosarcina in all systems manifested that aceticlastic methanogenesis was the major methanogenic pathway in digesters of food waste regardless of TAN or sulfate systems. The results were consistent with Yi et al. [38], who reported that Na+ existed in food waste was the potential key factor determining the pathway of methane production. Methanosarcina were not significantly inhibited by the Na+ level during the AD process [38]. Notably, as compared to other TAN systems, the percentage of Methanosarcina decreased obviously in TAN6500, implying that such a high concentration of ammonia in the digesters inhibited the aceticlastic methanogenesis pathways, which caused a significant TVFAs accumulation and thus a declining of CH4 production. In other words, the decreasing percentage of Methanosarcina led to the lower conversion efficiency of organics from the liquid phase into CH4 (2.68%) in TAN6500. Similar phenomenon of Methanobacterium, related to hydrogenotrophic methanogens, was also observed in TAN6500 [14], suggesting that high ammonia exhibited a negative effect on Methanobacterium. However, Methanoculleus, which also belongs to hydrogenotrophic methanogens, increased significantly in TAN6500. Amha et al. [39] found that Methanoculleus exhibited a higher resilience or adaptation potential to high fats, oils and grease, which could be used to explain this phenomenon in TAN6500. From above, we could speculate that high ammonia concentration facilitated a shift in methane-producing pathways from aceticlastic methanogenesis to hydrogenotrophic methanogens. Notably, the relative abundance of Methanosarcina decreased with the increasing sulfate concentrations, indicating that the activity of Methanosarcina was closely influenced by the sulfate concentrations. Contrarily, the relative abundance of Methanobacterium (hydrogenotrophic methanogens) increased with the increasing sulfate concentration in S1-S4 systems, demonstrating that high sulfate concentrations could improve H2-consuming methanogens activity [40]. And the horizontal flow AD reactor was discussed in Text S10 (Supporting information).
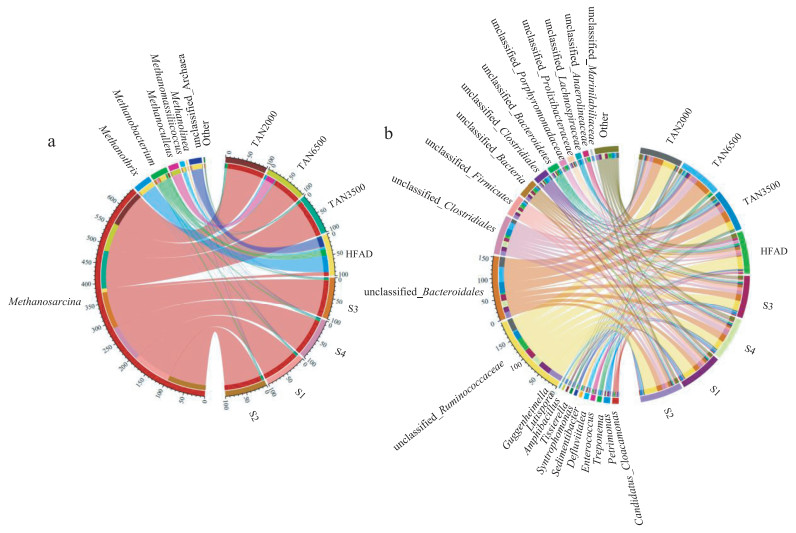
Firmicutes (43.78%-63.17%), Bacteroidetes (24.20%-33.305%), Cloacimonetes (0.44%-2.42%), Spirochaetes (0.39%-4.67%) and Chloroflexi (0.33%-1.89%) were the dominant phylum in all AD systems (Fig. S7 in Supporting information). Several previous studies have proven that the dominance of Firmicutes, Bacteroidetes and Proteobacteria in AD reactors played a key role in hydrolysis and acetogenesis [38,41]. Hence, the high proportion of hydrolytic bacteria in all AD systems, regardless of the existence of ammonia/sulfate, further revealed the excellent transformation of C from solid phase to liquid phase (20.87%-26.67% in TAN systems, and 7.33%-16.65% in sulfate systems). As for genus level bacteria (Fig. 4b), Treponema as a presumptive homoacetogen could utilize H2 and CO2 to generate acetate, and worked synergistically with aceticlastic methanogens [42]. Therefore, their percentage distribution decreasing in TAN6500 closely related to the ammonia inhibition or the decrease of Methanosarcina. Notably, the unclassified bacteria accounted more than 70% of total bacteria in all systems. Despite the function of unclassified bacteria could not identified directly, it could be speculated the functions from the family level. For example, the genus bacteria within family Ruminococcaceae were capable of hydrolyzing and fermenting the cellulose and hemicellulose related carbohydrates [43,44]. Hence, the higher relative abundance of unclassified_Ruminococcaceae contributed to the hydrolysis and fermentation of the organics within the food waste digesters. Similarly, unclassified_Clostridiales might involve in the hydrolysis and acidogenesis process of the AD systems, because the family Clostridiales were known to produce cellulases, lipases, proteases, and other extracellular enzymes [9]. Notably, these unclassified bacteria differed obviously among TAN systems. For instance, lowest relative abundance of unclassified_Ruminococcaceae in TAN6500 systems was observed, suggesting high ammonia concentrations could inhibit this type hydrolysis, while supported unclassified_Bacteroidales activity due to its higher abundance. Additionally, higher relative abundance of unclassified_Lachnospiraceae were obtained in TAN3500 and TAN6500 meant the higher hydrolysis rates of protein [42]. The bacterial dynamics changed slightly at genus level in sulfate systems owing to the similar community structures (Fig. 4b).
In summary, inhibition of ammonia and sulfate on the digestion of food waste were mainly caused by VFAs accumulation, especially for propionic acid. Ammonia inhibition was presented with 6500 mg/L, while sulfate inhibition occurred with the range of 1600-3500 mg/L. Element analysis indicated that the higher rate of C dissolution than conversion caused SCODpropionic acid accumulation. From microbial community structures, aceticlastic methanogenesis was the major methanogenic pathway for methane production. High TAN (6322 mg/L) levels showed toxicity on the Methanosarcina in TAN systems. Unclassified bacteria within Firmicutes and Bacteroidetes phyla played an important role in substrate hydrolysis.
The authors report no declarations of completing interests.
This research was financed by the National Key Research and Development Program of China (No. 2018YFC1900902), the State Key Laboratory of Urban Water Resource and Environment (No. 2020TS01), the Heilongjiang Nature Science Foundation (No. YQ2020E022) and the National Nature Science Foundation of China (No. 51878213).
Supplementary material associated with this article can be found, in the online version, at doi:
Q. Wu, H. Li, X. Hu, et al., Chin. Chem. Lett. 31(2020) 2825–2830. doi: 10.1016/j.cclet.2020.06.029
X. Dai, N. Duan, B. Dong, et al., Waste Manage. 3(2013) 308–316.
T. Luo, H. Huang, Z. Mei, et al., Chin. Chem. Lett. 30(2019) 1219–1223. doi: 10.1016/j.cclet.2019.03.018
T. Chen, Y. Jin, F. Liu, et al., J. Environ. Manage. 106(2012) 17–21. doi: 10.1016/j.jenvman.2012.03.045
F. Shen, H. Yuan, Y. Pang, et al., Bioresour. Technol. 144(2013) 80–85. doi: 10.1016/j.biortech.2013.06.099
Y. Ren, M. Yu, C. Wu, et al., Bioresour. Technol. 247(2019) 1069–1076.
L. Li, X. Peng, X. Wang, et al., Bioresour. Technol. 248(2018) 20–28. doi: 10.1016/j.biortech.2017.07.012
O. Kuczman, M.V.D. Gueri, S.N.M. De Souza, et al., J. Clean. Prod. 196(2018) 382–389. doi: 10.1016/j.jclepro.2018.05.282
X. Peng, S. Zhang, L. Li, et al., Bioresour. Technol. 262(2018) 148–158. doi: 10.1016/j.biortech.2018.04.076
A. Serna-Maza, S. Heaven, C.J. Banks, Bioresour. Technol. 152(2014) 307–315. doi: 10.1016/j.biortech.2013.10.093
H. Nie, H.F. Jacobi, K. Strach, et al., Bioresour. Technol. 178(2015) 238–246. doi: 10.1016/j.biortech.2014.09.029
A. Mahdy, I.A. Fotidis, E. Mancini, et al., Bioresour. Technol. 225(2016) 272–278.
S. Poirier, E. Desmond-Le Quéméner, C. Madigou, T. Bouchez, O. Chapleur, Bioresour. Technol. 207(2016) 92–101. doi: 10.1016/j.biortech.2016.01.124
C. Gonzalez-Fernandez, P.A. Garcia-Encina, Biomass Bioenergy 33(2009) 1065–1069. doi: 10.1016/j.biombioe.2009.03.008
H. Fisgativa, A. Tremier, P. Dabert, Waste Manage. 50(2016) 264–274. doi: 10.1016/j.wasman.2016.01.041
Z. Cetecioglu, J. Dolfing, J. Taylor, et al., Water Res. 155(2019) 444–454. doi: 10.1016/j.watres.2019.02.038
C. O'Reilly, E. Colleran, FEMS Microbiol. Ecol. 56(2006) 141–153. doi: 10.1111/j.1574-6941.2006.00066.x
H. Li, F. Tan, L. Ke, et al., Chem. Eng. J. 287(2016) 329–336. doi: 10.1016/j.cej.2015.11.003
C. Alcántara, P.A. García-Encina, R. Muñoz, Chem. Eng. J. 221(2013) 238–246. doi: 10.1016/j.cej.2013.01.100
R. Rajagopal, J.W. Lim, Y. Mao, et al., Sci. Total Environ. 443(2013) 877–886. doi: 10.1016/j.scitotenv.2012.11.016
Standard APHA, Methods for the Examinations of Water and Wastewater, 21st ed., APHA. AWWA and WEF DC, Washington, 2005.
J. Procházka, P. Dolej, J. Máca, et al., Appl. Microbiol. Biot. 93(2012) 439–447. doi: 10.1007/s00253-011-3625-4
R. Rajagopal, D.I. Massé, G. Singh, Bioresour. Technol. 143(2013) 632–641. doi: 10.1016/j.biortech.2013.06.030
W. Zhao, H. Yang, S. He, Q. Zhao, L. Wei, Bioresour. Technol. 341(2021) 125797. doi: 10.1016/j.biortech.2021.125797
A. Karlsson, J. Ejlertsson, Biochem. Eng. J. 61(2012) 43–48. doi: 10.1016/j.bej.2011.12.003
C. Sun, W. Cao, C.J. Banks, et al., Bioresour. Technol. 218(2016) 1215–1223. doi: 10.1016/j.biortech.2016.07.082
J.H. Park, G. Kumar, Y.M. Yun, et al., Bioresour. Technol. 248(2018) 134–140. doi: 10.1016/j.biortech.2017.07.025
E. Lee, P. Bittencourt, L. Casimir, et al., Waste Manage. 95(2019) 432–439. doi: 10.1016/j.wasman.2019.06.033
Z. Qian, T. Hao, H.R. Mackey, et al., Water Res. 150(2019) 162–181. doi: 10.1016/j.watres.2018.11.018
Y. Yang, Q. Chen, J. Guo, et al., Water Res. 87(2015) 112–118. doi: 10.1016/j.watres.2015.09.012
A. Hussain, S.K. Dubey, Appl. Water Sci. 7(31) 535-542.
Y. Li, Y. Jin, A. Borrion, et al., Bioresour. Technol. 244(2017) 213–224. doi: 10.1016/j.biortech.2017.07.006
L. Wei, Y. Ren, F. Zhu, et al., J. Water Process. Eng. 255(2021) 117668.
I. Rocamora, S.T. Wagland, R. Villa, et al., Bioresour. Technol. 299(2020) 122681. doi: 10.1016/j.biortech.2019.122681
Q. Li, Y. Li, Q. Wei, et al., Bioresour. Technol. 185(2015) 308–315. doi: 10.1016/j.biortech.2015.03.019
H. Du, F. Li, Chemosphere 168(2017) 1022–1031. doi: 10.1016/j.chemosphere.2016.10.108
L. Feng, S. He, L. Wei, et al., Environ. Res. 200(2021) 111415. doi: 10.1016/j.envres.2021.111415
J. Yi, B. Dong, Y. Xue, et al., J. Microbiol. Biotechn. 24(2014) 270–279. doi: 10.4014/jmb.1306.06067
Y.M. Amha, P. Sinha, J. Lagman, et al., Water Res. 123(2017) 277–289. doi: 10.1016/j.watres.2017.06.065
N.D. Jonge, S. Davidsson, J. Jansen, et al., Waste Manage. 117(2020) 124–135. doi: 10.1016/j.wasman.2020.07.047
S.M. Shaban, I. Aiad, M.M. El-Sukkary, et al., Chin. Chem. Lett. 26(2015) 1415–1420. doi: 10.1016/j.cclet.2015.06.006
C. Dennehy, P.G. Lawlor, M.S. Mccabe, et al., Waste Manage. 71(2018) 532–541. doi: 10.1016/j.wasman.2017.10.047
L. Zhou, Y. Yan, X. Pei, et al., Chin. Chem. Lett. 33(2022) 428–433. doi: 10.1016/j.cclet.2021.06.055
L. Lin, Z. Yu, Y. Li, Bioresour. Technol. 241(2017) 1027–1035. doi: 10.1016/j.biortech.2017.06.011

Figure 1 Methane production (a, c) and variations of methane production yield (b, d) of anaerobic digestion reactors under different ammonia/sulfate concentrations.

Figure 2 (a-d) Changes of VFAs concentration during the whole 60 d digestion under different concentrations of ammonia nitrogen additive in food waste.

Figure 3 The variations of soluble protein (a), FAN (b) and TAN (c) concentrations under different ammonia conditions.

 扫一扫看文章
扫一扫看文章

扫一扫关注我们
 DownLoad:
DownLoad:
 下载:
下载:

 下载:
下载: