Citation:
Wang Zhangjie, Chi Lianli. Recent advances in mass spectrometry analysis of low molecular weight heparins[J]. Chinese Chemical Letters,
2018, 29(1): 11-18.
doi:
10.1016/j.cclet.2017.08.050
Recent advances in mass spectrometry analysis of low molecular weight heparins
Received Date:
13 June 2017 Accepted Date:
24 August 2017 Revised Date:
25 July 2017 Available Online:
22 January 2018
Abstract:
Carbohydrates are important biomolecules and promising novel drug candidates, but their structural complexity and heterogeneity hinder their study and application. Low molecular weight heparins (LMWHs) are widely used as anticoagulants in the clinic and are considered the most successful carbohydrate-based drugs. They consist of both natural structures inherited from parent heparin and modified structures derived from depolymerization reactions. Sophisticated analytical methods are in great demand to elucidate the fine structure of LMWHs. This review summarizes recent progress in mass spectrometry (MS) that facilitates the in-depth structural characterization of LMWHs. The strategies can be sorted into top-down approaches and bottom-up approaches. In top-down approaches, the intact oligosaccharides are analyzed directly by hyphenated MS techniques to reveal their distribution and composition. Bottom-up approaches provide complementary structural information by analyzing partially enzymatically digested fragments or exhaustively degraded disaccharide building blocks of LMWHs. Advances in the MS/MS sequencing of short oligosaccharides and bioinformatics tools are also reviewed. Multidimensional analysis by MS offers a rigorous inspection of LMWHs, which is indispensable for ensuring drug quality and expanding their applications.
Low molecular weight heparins (LMWHs) are a class of widely used anticoagulant drugs derived from the chemical or enzymatic depolymerization of porcine intestine heparin [1]. As carbohydrate-based drugs, LMWHs exhibit structural features distinct from conventional small molecule drugs and modern proteinbased biomacromolecule drugs. They are comprised of numerous oligosaccharide chains with different chain lengths, monosaccharide compositions, sulfo and N-acetyl substitutions, and sequences. The importance of the structural characterization of heparin and LMWHs was underestimated for a long period until the global heparin contamination crisis occurred in 2008 [2, 3]. The lessons learned are that traditional analytical tests, such as molecular weight (MW) analysis by gel permeation chromatography, monosaccharide composition analysis and bioactivity analysis, are not sufficient to reflect the fine structure of a polysaccharide drug and ensure its safety and efficacy [4, 5].
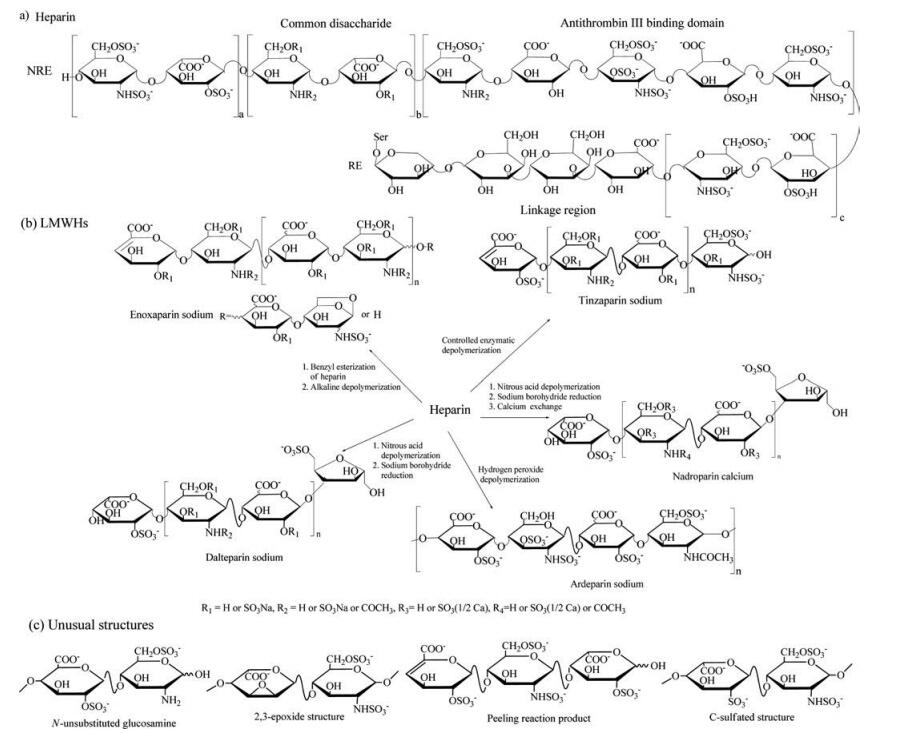
Heparin is constructed of repeating disaccharide units of a glucuronic acid residue (GlcA) or iduronic acid residue (IdoA) 1, 4-linked to a glucosamine residue (GlcN), with various substitution patterns of sulfation at the 2-O-position of the hexuronic acid residue (HexA), the 3-O, the 6-O and/or the N-position of GlcN, and N-acetylation at GlcN [6]. A unique pentasaccharide sequence of —GlcNS/Ac6S-GlcA-GlcNS3S6S-IdoA2S-GlcNS6S-and a yet to be completely elucidated octadecasaccharide sequence in heparin specifically activate ATⅢ to inhibit coagulation factor Xa and factor Ⅱa, respectively, and together execute the anticoagulant activity [7-9]. The reducing end (RE) of heparin is a linkage tetrasaccharide, —GlcA-Gal-Gal-Xyl-, through which it covalently links to the serine core of proteoglycan during its synthesis and elongation in the Golgi apparatus (Fig. 1a) [10]. The backbone structure of heparin is essentially retained in LMWHs, while the termini of newly created oligosaccharides are generally modified by chemical or enzymatic depolymerization reactions. The terminal structures are characteristic of each type of LMWH. As shown in Fig. 1b, enoxaparin sodium, produced by alkaline depolymerization of the benzyl ester derivative of heparin, possesses a 4-enopyranose uronate residue at the non-reducing end (NRE), and a minority portion (ranging from 15% to 25% of the oligosaccharide chains) has a 1, 6-anhydro structure at the RE [11]. Dalteparin sodium and nadroparin calcium are manufactured via nitrous acid degradation followed by sodium borohydride reduction. As a result, their RE is modified to a 6-O-sulpho-2, 5-anhydro-D-mannitol structure [12, 13]. Tinzaparin is the controlled enzymatically digested product of heparin, for which the NRE is an unsaturated uronic acid structure due to cleavage by heparinase from Flavobacterium heparinum [14, 15]. Ardeparin sodium is derived from the hydrogen peroxide degradation of heparin, with a saturated uronic acid residue at the NRE [16, 17]. In addition to these natural and modified structures, a variety of uncommon byproducts generated by side reactions have been reported. For example, odd-numbered oligosaccharides are generated by a peeling reaction that sequentially peels off monosaccharide residues from the RE [18]. N-Unsubstituted glucosamine residues result from the neutral loss of N-sulfo groups during the heparin and LMWH manufacturing process [19]. A 2, 3-epoxide structure is the 2-O-desulfated derivative of IdoA2S under the basic condition during heparin purification, which can be further converted to another unusual structure, a rearranged uronic acid residue containing a carbon-sulfur (C-S) bond (Fig. 1c) [20, 21]. Although these unusual structures are in very low abundance in LMWH products, they are important indicators of the process conditions.
图 1
图 1
Structures of heparin and LMWHs. (a) Structure of heparin. (b) Manufacturing process and characteristic structures of LMWHs. (c) Unusual structures in LMWHs derived by side reactions.
Figure 1.
Structures of heparin and LMWHs. (a) Structure of heparin. (b) Manufacturing process and characteristic structures of LMWHs. (c) Unusual structures in LMWHs derived by side reactions.
In recent years, extensive efforts from scientific research groups and pharmaceutical industries have been devoted to the development of sophisticated structural characterization approaches for LMWHs. Understanding the detailed structures is essential to improving the clinical use of LMWHs. Due to several advantages, including a more predictable dose-response relationship, less risk of bleeding, better bioavailability and a longer half-life, LMWHs are becoming more favored in the clinic over unfractionated heparin [22]. However, the therapeutic interchange between LMWHs in the prevention and treatment of thrombotic diseases is still controversial and needs further investigation into the properties of LMWHs produced using different methods [23]. LMWHs are also considered promising novel drug agents for the treatment of inflammatory disease, cancer and infectious disease, which requires the isolation and characterization of specific functional motifs in addition to the anticoagulant motif to elucidate the mechanism [24]. Since the approval of the first generic LMWH by the US Food and Drug Administration (FDA) in 2010, an urgent need for developing structural profiling methods has arisen for the comparison between innovator and generic LMWHs [25, 26]. Because it is impracticable to sequence and quantify all individual oligosaccharide chains in LMWHs, a framework consisting of the characterization of several key aspects with analytical advancements was presented by the FDA to allow an in-depth and comprehensive evaluation of LMWH products (Fig. 2). The heterogeneity of LMWHs can be determined by their overall physicochemical properties, high-resolution intact chain mapping, fragment mapping, disaccharide building blocks and the sequence of oligosaccharide species [27]. Traditional analytical methods, such as liquid chromatography (LC), gel electrophoresis, capillary electrophoresis (CE) and nuclear magnetic resonance (NMR), cannot provide precise structural information on the individual components in LMWHs [28-31]. Mass spectrometry (MS) techniques, especially electrospray ionization (ESI)-MS linked to LC or CE, are therefore indispensable in the characterization of LMWHs, especially for various chain mapping analyses and sequence analysis methods.
图 2
图 2
Integrated analytical techniques for evaluating the key structural properties of LMWHs.
Figure 2.
Integrated analytical techniques for evaluating the key structural properties of LMWHs.
This article reviews recent research advances in the MS analysis of LMWHs. The development and application of various LC-MS, CEMS and MS/MS approaches for intact chain mapping, fragment mapping, disaccharide building blocks and sequences of oligosaccharide species are summarized, and the advantages and challenges of the approaches are discussed.
LMWHs generally have a weight-averaged MW below 8000 Da. For example, the weight-average MW of enoxaparin is defined as between 3800 and 5000 Da, with no more than 18% of oligosaccharide chains having a MW above 8000 Da. Dalteparin has a characteristic weight-average MW of approximately 6000 Da, corresponding to a degree of depolymerization of (dp)18. With the help of improved ESI techniques, increased mass analyzer resolution and bioinformatics tools, the intact chains of glycosaminoglycans with sizes over dp40 have been successfully analyzed [32, 33], which satisfies the requirement for covering the majority of oligosaccharide components in LMWHs.
Intact chain mapping analysis provides information on the distribution and relative abundance of oligosaccharides with different sizes, monosaccharide compositions and substitutions. Conventional size exclusion chromatography (SEC) has low resolution and cannot demonstrate the compositions of LMWH chains. Strong anion exchange chromatography (SAX) and upgraded cetyltrimethylammonium-coated SAX (CTA-SAX) can efficiently separate highly sulfated LMWHs, but they cannot be connected to MS due to the high concentration of involatile salts used in the mobile phases [34, 35]. Ultraperformance size exclusion chromatography (UPSEC), reversed phase ion pairing chromatography (RPIP) and hydrophilic interaction chromatography (HILIC) methods have been developed to replace SAX and CTA-SAX in the separation of highly negatively charged carbohydrates, and many attempts using these ESI-MS compatible chromatographies to profile the fingerprints of LMWHs have been reported in recent years.
Zhang et al. have demonstrated the use of UPSEC coupled to a hybrid quadruple time-of-flight (Q-TOF) mass spectrometer to analyze enoxaparin sodium [36]. More than 70 components were identified and sorted into 6 groups: even-numbered oligosaccharides with unsaturated NRE, even-numbered oligosaccharides with saturated NRE, even-numbered oligosaccharides containing 1, 6-anhydro RE, odd-numbered oligosaccharides with saturated HexA at both termini, odd-numbered oligosaccharides with GlcN at both termini and odd-numbered oligosaccharides with unsaturated HexA at NRE. This method is simple and robust and reflects the structural changes of LMWHs in response to manufacturing processes, potentially revealing the contribution of individual components to the overall bioactivity of LMWHs. An online desalting device has been developed to minimize ammonium adducts after SEC separation and simplify the mass profiles, which allows the assignment of LMWH oligosaccharides with sizes up to dp30 [37].
Since SEC resolves molecules mainly by their size, a variety of RPIP methods have been developed to achieve better resolution, especially for oligosaccharides of the same size but with different substitution patterns. Linhardt's group first established a RPIP-MS method of using tributylamine (TBA) to separate heparin-derived oligosaccharides on a C18 column [38]. Doneanu, Chen and Gebler evaluated the performance of a series of linear and branched aliphatic amine ion pairing reagents, including n-propylamine, nbutylamine, n-pentylamine (PTA), n-hexylamine, n-octylamine, tripropylamine and TBA, in the separation of heparin-derived oligosaccharides [39]. PTA exhibited the best efficiency in separating charged oligosaccharides and enhancing mass response using the unique additive, 1, 1, 1, 3, 3, 3-hexafluoro-2-propanol (HFIP). PTA also offered positive charges, making the negatively charged oligosaccharides detectable in the positive ion mode. Characterization of tinzaparin sodium using this method was demonstrated, and the largest identifiable oligosaccharide using its accurate MW detection was a dp22. Our group further optimized the PTA RPIPMS method and applied it to the analysis of enoxaparin sodium and nadroparin calcium [40]. Some minor oligosaccharide components arising from slightly miscontrolled processing conditions were disclosed. These impurities add risk to LWMH drug safety, but are easily overlooked by other analytical approaches that are not as sensitive and specific as the RPIP-ESI-MS method. Two-dimensional chromatography combing SEC and RPIP has been attempted by Ouyang and coworkers. A multiple heart-cutting system that automatically collects the hearts of fractions from SEC then injects them into RPIP was used to show a complete picture of the structural complexity of LMWHs [41]. This approach provides information on both the size distributions and sequences of enoxaparin and nadroparin, which makes it invaluable in the quality control of LMWHs and the glycomics study of other glycosaminoglycans. RPIP methods resolve LMWH oligosaccharides better than any other MS-compatible chromatography, but the accumulation of ion-paring reagents will eventually cause the contamination of mass spectrometers. In addition, the orbitrap mass analyzer, which has become prevalent in the structural analysis for biomacromolecules, cannot tolerate the ion-pairing reagents.
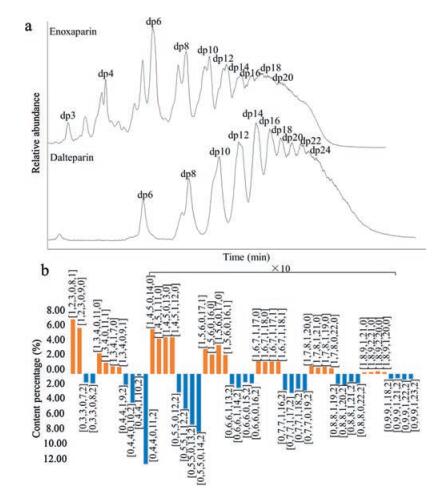
HILIC-MS is recognized as a promising new analytical platform for separating highly sulfated and very polar heparin oligosaccharides [42]. Ambient mobile phases with a low concentration of ammonium salts are needed, making HILIC universally adapted to any MS instrument without sacrificing ionization efficiency. The immobilized functional groups on the stationary phases of HILIC, including amides, diols and cross-linked diols, are suitable for retaining heparin and LMWH oligosaccharides. Li et al. established a top-down approach for the direct characterization of LMWHs using a cross-linked diol HILIC column coupled to a Fourier transform LTQ-orbitrap mass spectrometer [43]. The structures of enoxaparin oligosaccharides ranging from dp3 to dp28 were determined and designated to a formula to indicate the presence or absence of unsaturated NRE, the number of HexA, the number of GlcN, and the number of acetyl groups and sulfo groups. The minority components with a 1, 6-anhydro terminus at their RE were also identified. Quantitative comparison of enoxaparin sodium and its generic version were demonstrated to show their similarity and discrepancy. The HILIC-MS method is robust, reproducible and easy to use. It was applied to a large-scale comparison study of the innovator and generic versions of enoxaparin sodium marketed in the US and Europe [44]. The intact chain mapping analysis of two different LMWHs, enoxaparin sodium and dalteparin sodium, is presented in Fig. 3.
图 3
图 3
Intact chain mapping of LMWHs. (a) Total ion chromatograms of HILIC-ESIMS analysis of enoxaparin and dalteparin. (b) Distribution and composition of major oligosaccharide species. Oligosaccharides are presented as [NRE (0 for saturated NRE and 1 for △HexA), HexA, GlcN, Ac, SO3, RE (0 for GlcN, 1 for 1, 6-anhydro and 2 for 2, 5-anhydro-mannitol)].
Figure 3.
Intact chain mapping of LMWHs. (a) Total ion chromatograms of HILIC-ESIMS analysis of enoxaparin and dalteparin. (b) Distribution and composition of major oligosaccharide species. Oligosaccharides are presented as [NRE (0 for saturated NRE and 1 for △HexA), HexA, GlcN, Ac, SO3, RE (0 for GlcN, 1 for 1, 6-anhydro and 2 for 2, 5-anhydro-mannitol)].
CE, or more specifically, capillary zone electrophoresis (CZE), is an efficient way to separate charged analytes and is usually complementary to LC separations. The interface is the key to establishing a successful CE-MS method for characterizing LMWHs. Sun et al. explored the first use of a newly developed electrokinetic pump-based CE-MS interface for intact chain mapping analysis [45]. LMWH oligosaccharides were detected in the positive ion mode, assisted by the ammonium in the electroosmotic buffer. Later, Lin et al. applied a volatile methanolic ammonium acetate electrolyte and sheath fluid in the CE separation and linked it to negative ion ESI-MS [46]. Compared to LC-MS methods, the CE-MS methods are much more rapid and consume very small quantities of running buffer, making them potentially high-throughput, cost-effective means for chain mapping analysis of LMWHs. Further instrument development, method development and validation of CE-MS have bright prospects for fulfilling the demand for rapid and ultrasensitive analysis of complex LMWHs.
Intact chain mapping conducted by LC-MS or CE-MS analysis offers rich structural information about the chain distribution and composition of LMWHs. It not only identifies and quantifies the major and minor oligosaccharide components but also discloses trace amounts of unusual structures or impurities without bias. While intact chain mapping provides direct evaluation of the structural heterogeneity of LMWHs, additional analyses from other aspects are necessary to comprehensively profile their structures.
Fragmentmapping refers to the partial digestion of LMWHs using a single enzyme or controlled conditions to generate oligosaccharide fragments, followed by profiling of the digests using analytical approaches such as LC-MS. Heparinases, including heparinase Ⅰ (Hep Ⅰ, Enzyme Commission (EC) # 4.2.2.7), heparinase Ⅱ (Hep Ⅱ, no EC # assigned) and heparinase Ⅲ (Hep Ⅲ, EC # 4.2.2.8), are enzymes isolated from Flavobacterium heparinum that cleave the β-1, 4-glycosidic linkage between GlcN and HexA of heparin and LMWHs. During cleavage, a double bond is introduced at the C4-C5 of the HexA at the NRE of the newly generated oligosaccharide chains, which incorporates a chromophore with characteristic ultraviolet (UV) absorption at 232 nm [47]. The three kinds of heparinases each display unique specificity depending on the stereoisomerism of uronic acid and the sulfation patterns of the oligosaccharide chains. Hep Ⅰ and Ⅲ have restricted substrate specificities; for example, Hep Ⅰ prefers to cleave the glycosidic bond between GlcNS and IdoA2S, while the low sulfation domain, GlcNAc or GlcNS linked to GlcA, is the optimum substrate for Hep Ⅲ. Hep Ⅱ exhibits broad substrate specificity that tolerates O-sulfation of HexA and GlcA residues [48]. Partial digestion of heparin oligosaccharide by strategic use of a single heparinase or an enzyme combination can reduce structural complexity while retaining the specific sequence of interest, which allows reconstruction of the entire sequence of the original oligosaccharide chain with the help of a property-encoded scheme [49]. Since LMWHs are complex mixtures, fragment mapping represents a global signature of oligosaccharide sequences that constitutes all intact chains of the LMWHs.
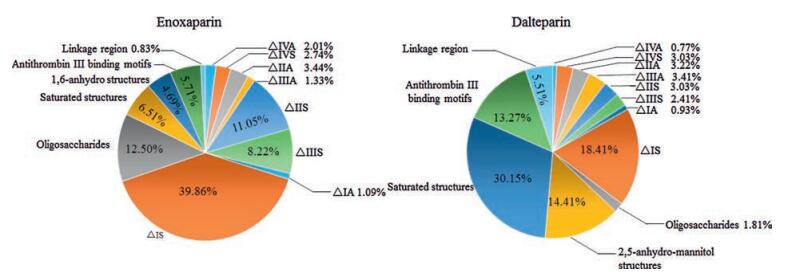
Li et al. has established a bottom-up approach utilizing Hep Ⅱ to digest enoxaparin sodium, followed by HILIC-ESI-MS analysis on an LTQ-orbitrap mass spectrometer [50]. Qualitative and quantitative analyses of more than 40 oligosaccharide fragments were achieved. The major fragment structures were unsaturated disaccharides, arising from the prominent repeating units of the backbone of LMWHs. Unsaturated 3-O-sulfated tetrasaccharides arising from antithrombin Ⅲ binding sites were identified rather than disaccharides, because these structures are resistant to Hep Ⅱ. Fragments containing the 1, 6-anhydro ring are from the original RE of a minority of enoxaparin chains. Some structural features of parent heparin, including saturated uronic acid oligosaccharides and linkage or oxidized linkage regions, are also found in the enzymatic digests (Fig. 4). This finding suggests that the bottom-up method is useful to evaluate the structure of LMWHs as determined from the depolymerization conditions as well as from the starting materials. We performed a more comprehensive fragment mapping study by using Hep Ⅰ, Ⅱ and Ⅲ to partially digest enoxaparin sodium and nadroparin calcium [51]. The digestion conditions were optimized to generate a rich pool of oligosaccharide fragments from disaccharides to decasaccharides, as visualized by polyacrylamide gel electrophoresis. A RPIP-ESI-MS method was performed on an ion trap (IT)-TOF hybrid mass spectrometer to plot the fingerprinting of enoxaparin and nadroparin digests by three kinds of heparinases. The fine structures of 94 oligosaccharide fragments from enoxaparin and 109 oligosaccharide fragments from nadroparin were identified, providing a multiplexed evaluation of the composition and sequence of LMWHs.
图 4
图 4
Comparison of Hep Ⅱ-digested fragments between enoxaparin and dalteparin by fragment mapping analysis.
Figure 4.
Comparison of Hep Ⅱ-digested fragments between enoxaparin and dalteparin by fragment mapping analysis.
Reoccurring disaccharide building blocks are the basic structural motifs that construct heparin polysaccharides. In addition to the eight common heparin disaccharides, LMWHs also contain modified disaccharides at one or both termini, arising from chemical or enzymatic depolymerization reactions during manufacturing. These modified disaccharides are characteristic to each type of LMWH. Some unusual components that result from inevitable side reactions may also exist at very low abundances. Thus, disaccharide building block analysis is a fundamental way to characterize the structure of LMWHs.
A cocktail of Hep Ⅰ, Ⅱ and Ⅲ is generally used to exhaustively digest LMWHs into disaccharide building blocks. Some uncommon structures may be resistant to heparinases and present as small oligosaccharides, such as trisaccharides and tetrasaccharides, in exhaustive enzymatic digests. The conventional disaccharide analysis of heparin is restricted to the eight common disaccharides [52]. A variety of analytical methods have been developed for LMWHs that aim to also cover modified disaccharides and unusual structures.
The SAX method has been used to profile the disaccharide building blocks of enoxaparin sodium, but some components arising from the NRE, such as saturated disaccharides derived from the original NRE of parent heparin, lack a chromophore and are not detectable by UV [53]. This is more problematic for dalteparin sodium and nadroparin calcium, which contain characteristic saturated NREs. Fluorescent molecules such as 2-aminoacridone (AMAC) have been used to introduce a fluorophore into heparin disaccharides and oligosaccharides [54]. This also changes the hydrophobicity of heparin disaccharides and oligosaccharides, making them better resolved on reversed phase chromatography. However, disaccharides and oligosaccharides with modified REs, such as the 1, 6-anhydro structure of enoxaparin and 2, 5-anhydro-D-mannitol of nadroparin calcium and dalteparin sodium, are not reactive during fluorescent labeling. We have used a diode array detector to monitor UV absorption at 232 nm for unsaturated uronic acid residues and at 255 nm for AMAC labels simultaneously [55]. A total of 15 basic building blocks for enoxaparin and 17 for dalteparin were identified and quantified using this method after exhaustive digestion and labeling with AMAC. Several hyphenated MS methods, including LC-MS and CE-MS, have recently been reported for profiling the disaccharide building blocks of LMWHs [36, 55-57]. These methods are limited to detecting 12 to 17 different building blocks due to insufficient separation resolution, detection sensitivity or specificity.
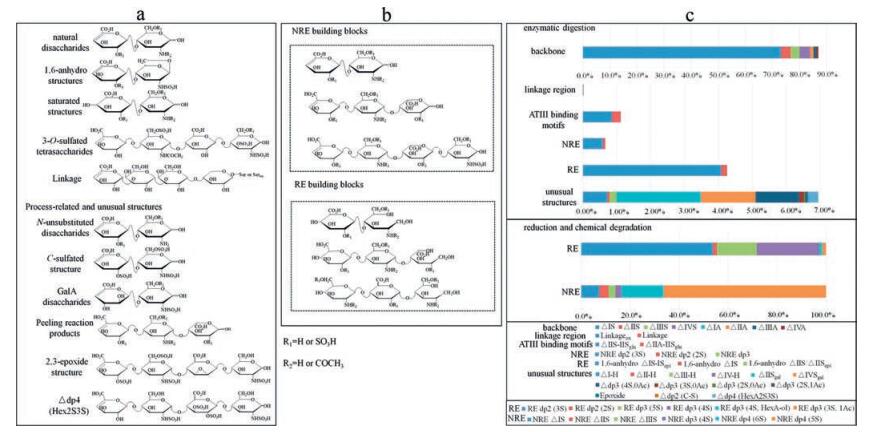
The most comprehensive analytical method covering all known enoxaparin building blocks has been established by combining the superior separation resolution of HILIC, the ultra-high resolution of the orbitrap mass spectrometer and the excellent sensitivity of the triple quadrupole mass spectrometer in multiple reaction monitoring (MRM) mode (Fig. 5) [58]. As many as 37 building blocks were identified, and 31 were successfully quantified from the enzymatic digests of enoxaparin sodium. Major disaccharides derived from the backbone and minor components from modified termini, active domains and side reactions were profiled to reflect the detailed structural characteristics and processing conditions of LMWHs. Some novel structures, including a disaccharide containing a C-S bond, a saturated trisaccharide containing two GlcN residues and a tetrasaccharide containing an unusual 3-O-sulfated HexA residue, were also discovered from enoxaparin sodium. Moreover, sodium borohydride reduction and oxidative degradation were used as supplements to the enzymatic digestion to differentiate the NRE, RE and backbone structures from one another. Additionally, 12 terminal building blocks were characterized as indicators to chemical depolymerization conditions during enoxaparin manufacturing. This result suggests that LMWHs are likely to be more structurally complicated than previously considered. The minor components and unusual structures are worth further investigation to elucidate their bioactivity and potential risks for LMWHs. We have also successively used the HILIC-MRM-MS method for basic building block analysis of dalteparin sodium and nadroparin calcium [59]. HILIC separation was optimized by using a maltose modified column to replace the diol column. A total of 36 basic building blocks were unambiguously identified and then quantified by MRM analysis. This pproach represents the most comprehensive disaccharide building block analysis method for LMWHs prepared using nitrous acid degradation.
图 5
图 5
Disaccharide building block analysis of enoxaparin. (a) Structures of exhaustively enzymatically digested basic building blocks. (b) Structures of reduced and chemically degraded terminal building blocks. (c) Composition of basic building blocks determined by LC-MRM-MS analysis of enoxaparin.
Figure 5.
Disaccharide building block analysis of enoxaparin. (a) Structures of exhaustively enzymatically digested basic building blocks. (b) Structures of reduced and chemically degraded terminal building blocks. (c) Composition of basic building blocks determined by LC-MRM-MS analysis of enoxaparin.
Disaccharide building blocks, or more accurately, basic building blocks, provide fundamental information on the structural properties of LMWHs. These building blocks can be sorted into different groups and reveal the essence of LMWHs from different aspects. For example, the common disaccharides, original heparin NREs and linkages reflect the structure of starting materials from which LMWHs are made, the 3-O-sulfated disaccharides and tetrasaccharides represent the functional domains for antithrombin Ⅲ binding and anticoagulant activity, the modified disaccharides and oligosaccharides are characteristic of each type of LMWH and indicate the depolymerization reaction conditions, and some unusual structures arising from undesired side reactions may affect the safety of LMWH drugs.
Heparin and other linear glycosaminoglycans are believed to have defined sequences, which are essential for their functions and bioactivities. During the depolymerization of LMWHs from heparin, the majority of antithrombin Ⅲ-binding pentasaccharide domains are retained, while the longer factor Ⅱa binding domains are more likely destroyed. This process makes the dose-response relationship of LMWHs more predictable, as the more complicated factor Ⅱa-involved coagulation mechanism is avoided [23]. The adverse effect of heparin and LMWHs known as heparin-induced thrombocytopenia and thrombosis is closely related to the sequences of oligosaccharides [60]. The ultimate characterization of LMWHs would be achieved by sequencing all oligosaccharide chains, but this is currently unpractical due to their extreme complexity and heterogeneity.
Alternatively, sequencing the short oligosaccharide pool of LMWHs provides sufficient evidence of the structural properties of LMWHs, because these short oligosaccharides likely undergo more cleavage reactions than longer species during the manufacturing process [27]. Ozug et al. have elucidated the tetrasaccharide pool in enoxaparin sodium using matrix-assisted laser desorption ionization (MALDI)-MS and NMR after purifying 15 oligosaccharide species with SEC and SAX [61]. The sequences and compositions of the predominant tetrasaccharides provide a sensitive, specific readout of the properties of enoxaparin and its physicochemical process conditions. However, the substantial number and high purity of samples required by NMR makes this approach very tedious.
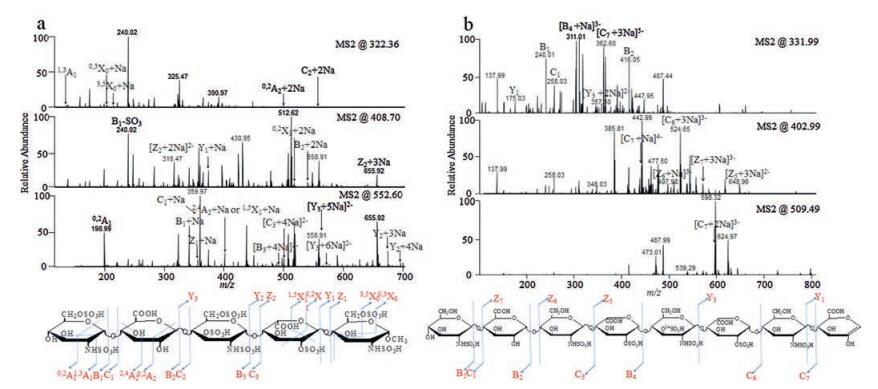
A tandem MS technique, also known as MS/MS, has been widely investigated as a powerful sequencing tool for heparin-derived oligosaccharides. Methods of ionization, including MALDI and ESI, as well as fragmentation methods, such as collision-induced dissociation (CID) and electron detachment dissociation (EDD), have been broadly explored [62-66]. The main challenge is to generate sufficient glycosidic bonds and cross-ring cleavage fragment ions while protecting the labile sulfo groups from neutral loss. Huang, Liu and Sharp developed a sequential permethylation, desulfation and trideuteroperacetylation approach to chemically replace fragile sulfo groups with acetyl groups containing stable isotopic labels. The original substitution positions of the sulfo groups were then determined by MS/MS. A heparan sulfate oligosaccharide mixture with saccharides as large as dodecasaccharides was sequenced [67, 68]. Amster's group established a direct CID-MS/MS sequencing method by adding sodium hydroxide to the spray solution to deprotonate all acidic groups, which stabilizes the sulfo groups during fragmentation [65, 66]. The synthetic heparin pentasaccharide with the sequence GlcNS6SGlcAGlcNS3S6SIdoA2SGlcNS6SOCH3 was used to mimic the antithrombin Ⅲ binding domain, which can be completely annotated (Fig. 6). The stereoisomerism of uronic acid residues was successfully distinguished by the presence of diagnostic ion 2, 4An for GlcA or its absence for IdoA. The sequencing of biochemically synthesized heparin oligosaccharides with CID-MS/MS was also demonstrated by different research groups [66, 69].
Although MS/MS has exhibited its power by sensitively and completely sequencing heparin oligosaccharides, the pool of short oligosaccharides must still be separated by LC to reduce the complexity, especially for isomers with different substitution patterns and stereoisomerism. However, HILIC is not good at resolving heparin oligosaccharide isomers. The ion paring reagents in RPIP are easily adducted to oligosaccharide molecules, which usually generates useless fragment ions. SAX offers satisfactory separation of oligosaccharides, but online or offline desalting is necessary before the eluent is introduced to the ionization interface. An efficient high-throughput SAX-ESI-MS/MS method for sequencing the tetrasaccharide pool of enoxaparin sodium and the hexasaccharide and octasaccharide pool of dalteparin sodium is currently under development in our group.
The demand for bioinformatics tools in the MS analysis of LMWHs involves the interpretation of complicated MS data and the processing of massive datasets from multiple LC-MS tests. Oligosaccharide ions are usually multiply charged and adducted with various numbers of sodium, ammonium and other neutral molecules, making the interpretation of ESI-MS data extremely difficult, especially for larger oligosaccharides.
We have previously developed a computer-assisted MS interpretation program to assign high-resolution mass spectra in intact chain mapping analysis [40]. A theoretical structure database containing all LMWH chains with possible compositions was generated. The monoisotopic m/z values corresponding to each oligosaccharide chain with different charge states and adduction forms were then calculated. The experimental m/z values were extracted and searched against the database for matches. Linhardt's group applied an opposite strategy to match experimental MWs to theoretical MWs for LMWHs. The raw data from high resolution mass spectra were deconvoluted using the open source software DeconTools. Next, the newly developed software package GlycReSoft was used to process the output of DeconTools to generate the identity and quantitative information for LMWH chains [43].
To evaluate the equivalence between multiple lots of LMWHs, the comparison tool GlycCompSoft was generated based on the programming language Microsoft ExcelTM Visual Basic for Applications. The lot-to-lot variability or biosimilarity between LMWHs from different manufacturers can be efficiently assessed [70]. Bioinformatics tools can also be used for the parent heparin and LMWH products. Principal component analysis was performed to process the data obtained by intact chain mapping, fragment mapping and disaccharide building block analysis, which enables the evaluation of the starting material and the manufacturing process based on the structural properties of LMWHs [71].
Interpretation of MS/MS spectra for heparin oligosaccharides is more challenging. A computational framework known as HS-SEQ was designed by Hu et al. to address the complex product ion patterns apparent in the negative electron transfer tandem MS analysis of heparan sulfate oligosaccharides. A divide-and-conquer strategy was used to assign the distributions of acetyl and sulfo groups on the oligosaccharide chains [72]. High-throughput identification software known as GAG-ID was developed by Chiu et al. to automatically interpret the LC-MS/MS dataset of the glycosaminoglycan mixture. Rapid, accurate sequencing of a moderately complex mixture of heparin tetrasaccharides was demonstrated [73]. The algorithms for processing larger, more complicated LMWH oligosaccharides are currently under development, and a more user-friendly version of software is anticipated.
Modern MS techniques offer sensitive and comprehensive tools for the structural characterization of LMWHs. Intact chain mapping by LC or CE linked to a high-resolution mass spectrometer demonstrates the distribution and composition of oligosaccharide chains in LMWHs. Fragment mapping of partially digested LMWHs with specific heparinase or controlled conditions provides an alternative way to reveal the alignment of reoccurring structural domains. Disaccharide building blocks of LMWHs, including common backbone disaccharides, characteristic terminal disaccharides, 3-O-sulfated structures and unusual structures, represent starting materials, depolymerization, active motifs and side reactions, respectively. The sequence of short oligosaccharide pool of LMWHs, which can be determined by MS/MS, reflects the chemical selectivity of depolymerization reactions and serves as a sensitive surrogate marker of manufacturing conditions. These MS-based analytical methods work together to define the structure of complicated and heterogeneous LMWHs from different standpoints.
Glycosaminoglycans are the most complex biomolecules, and the full determination of their sequences was once considered impossible. LMWHs are chemically modified drugs with increased structural complexity. Recent advances in MS, including ultra-high resolution mass analyzers, novel dissociation modes in MS/MS and new MS interfaces, facilitate the complete structural characterization of LMWHs. The package MS analysis of LMWHs, including intact chain mapping, fragment mapping, disaccharide building block analysis and MS/MS sequencing of short oligosaccharide pools, is indispensable to ensure drug quality and safety, assess lotto-lot variability and evaluate the equivalence between innovators' products and their generic versions. Associated with other rapidly developing techniques, such as bioinformatics and the biosynthesis of oligosaccharides, advances in MS project a promising future for better understanding glycosaminoglycans, elucidating their mechanisms, and discovering novel glycosaminoglycan-related biomarkers, drug targets and drug candidates.
Acknowledgment
We thank the support of the National Natural Science Foundation of China (No. 21472115).
H. Hu, Y. Huang, Y. Mao, et al., Mol. Cell. Proteomics 13(2014) 2490-2502. doi: 10.1074/mcp.M114.039560
[73]
Y. Chiu, R. Huang, R. Orlando, J.S. Sharp, Mol. Cell. Proteomics 14(2015) 1720-1730. doi: 10.1074/mcp.M114.045856
Figure 1
Structures of heparin and LMWHs. (a) Structure of heparin. (b) Manufacturing process and characteristic structures of LMWHs. (c) Unusual structures in LMWHs derived by side reactions.
Figure 3
Intact chain mapping of LMWHs. (a) Total ion chromatograms of HILIC-ESIMS analysis of enoxaparin and dalteparin. (b) Distribution and composition of major oligosaccharide species. Oligosaccharides are presented as [NRE (0 for saturated NRE and 1 for △HexA), HexA, GlcN, Ac, SO3, RE (0 for GlcN, 1 for 1, 6-anhydro and 2 for 2, 5-anhydro-mannitol)].
Figure 5
Disaccharide building block analysis of enoxaparin. (a) Structures of exhaustively enzymatically digested basic building blocks. (b) Structures of reduced and chemically degraded terminal building blocks. (c) Composition of basic building blocks determined by LC-MRM-MS analysis of enoxaparin.

 下载:
下载:


 下载:
下载:







 下载:
下载:
