引用本文:
孙梦雪, 于婵婵, 许敏, 冯雪艳, 姚立. DNA机械技术的研究进展[J]. 化学通报,
2020, 83(10): 866-874, 918.

Citation: Sun Mengxue, Yu Chanchan, Xu Min, Feng Xueyan, Yao Li. Advances in DNA Mechanical Technology[J]. Chemistry, 2020, 83(10): 866-874, 918.
Citation: Sun Mengxue, Yu Chanchan, Xu Min, Feng Xueyan, Yao Li. Advances in DNA Mechanical Technology[J]. Chemistry, 2020, 83(10): 866-874, 918.
DNA机械技术的研究进展
English
Advances in DNA Mechanical Technology
Abstract:
DNA mechanical technology has been widely concerned and deeply studied in recent years. Mechanical DNA devices have been enabled by the unique molecular structure and physical and chemical properties of DNA. More complex mechanical behavior or function can be achieved by designing more elaborate mechanisms. DNA mechanical technology has played important role in studies of the fundamental properties of the biomolecules that sense, transmit, and generate pN forces. Studies of the mechanics of these biomolecules are essential to revealing their functions. Here, we reviewed the latest progress in the study of DNA mechanical technology. First, the mechanical basis of DNA was briefly described. Especially several types of mechanical DNA devices were focused on. Finally, the challenges and prospects of DNA mechanical technology were discussed.
-
Key words:
- DNA mechanical technology
- / Molecular machine
- / Sensor
- / Molecular Interaction
-
生物大分子脱氧核糖核酸(DNA)作为活细胞中的遗传信息载体,其生物学功能包括存储和编码遗传信息。然而从材料科学和工程的角度来看,DNA独特的结构和性质使得它能够构建具有多种功能的装置。因此,DNA在分子生物学和材料科学与工程之间架起了桥梁,为新型纳米材料的合成铺平了道路。DNA可以由互补的DNA链通过非共价作用自组装,也可以通过酶催化的共价键连接。这些方法为设计和合成具有多种功能的DNA材料提供了巨大的可控性,也推动了一个我们称之为DNA机械技术的子领域的出现。在DNA机械技术领域,DNA被设计为可在纳米尺度上产生、传输和感知机械力。
自从Seeman在1982年[1]首次提出结构DNA纳米技术的概念以来,人们对制造可控制的和多功能的DNA纳米结构越来越感兴趣,DNA机械技术也受到了科学家们广泛的研究。生物学中有许多参与生物体各种生命活动的分子机器,由于尺寸很小,这种分子机器通常很难被制造出来。然而,通过对DNA特性的广泛研究,DNA机械技术能够实现功能纳米结构的构建。例如, DNA的分子识别能力已被广泛应用于纳米元件的结构导向,以产生纳米尺度精确控制的材料和器件[2~7]。
DNA机械技术在研究细胞感知、传递和产生皮牛(pN)力的蛋白质的基本特性方面发挥了重要作用[8]。这些蛋白质包括运动蛋白肌球蛋白、细胞骨架蛋白肌动蛋白和整合素受体,它们将细胞固定在周围环境中并感知机械力。这些生物分子的力学研究对于揭示它们的功能至关重要,通常我们可使用单分子力谱(SMFS)技术如光镊和磁镊对它们进行研究。然而,目前单分子力谱技术仍存在两个局限性:一是常规力谱的连续特性导致的低数据吞吐量;二是需要在目标分子和仪器之间通过长聚合物手柄建立连接[9]。这些手柄是高度灵活的,它们易受漂移和噪声的影响,特别是在与生理相关的小于10pN的力范围内。
为了避免上述问题,科学家利用DNA折纸技术创建由几个DNA螺旋捆绑在一起的刚性手柄,这些坚硬的手柄显示出较小的测量噪声,因此能够直接测量弱分子间键的机械力,例如相邻DNA碱基之间的叠加力[10]。在这种情况下,DNA机械技术正在取代传统的SMFS。其中利用DNA折纸技术[11]得到的各种纳米结构成为了DNA机械技术强有力的工具。基于DNA结构设计的可行性、多功能性和精确性,DNA折纸技术使得构建复杂的DNA机械装置成为可能,在单分子生物物理学、结构生物学和生物医学等领域具有重要意义[12~17]。
本文针对DNA的结构和性质,总结了近些年来DNA机械技术的研究进展。首先简要介绍DNA的力学基础,然后重点阐述几种DNA机械装置,并在此基础上进一步探究它们在生物研究和材料科学中的广泛应用,最后讨论了DNA机械技术的挑战与展望。
1. DNA力学基础
1.1 DNA结构和性质
DNA分子可以看作是由称为核苷酸的单体组成的聚合物。每个核苷酸有两个主要的组成部分:一个磷酸基和一个与碱基相连的糖基(脱氧核糖)。每个单体通过磷酸二酯键连接在一起。其中共有四种不同的单体,分别是四种不同的含氮碱基:腺嘌呤(A)、胸腺嘧啶(T)、鸟嘌呤(G)和胞嘧啶(C)。通常,DNA链的一端在糖环的5′碳上有一个磷酸基(定义为5′端),而另一端在糖环的3′碳上有一个羟基(定义为3′端)。DNA分子的第一个独特之处在于,它们可以是单链DNA(ssDNA),也可以是双链DNA(dsDNA)。为了形成dsDNA,两条互补的ssDNA链按照Watson-Crick碱基配对规则,腺嘌呤(A)与胸腺嘧啶(T)配对,胞嘧啶(C)与鸟嘌呤(G)配对,以反平行方向杂交成双螺旋结构。另一个独特之处在于,DNA双螺旋结构本质上是一种纳米尺度的物体。典型的B型DNA是右手双螺旋,直径约为2nm,碱基分离为0.34nm,螺旋周期为10个碱基对,螺距为3.4nm。这些参数能够帮助科学家在纳米尺度上精确地设计DNA结构。
DNA具有独特的性质,为设计和制造多功能纳米结构提供了优异的性能。从物理方面来说,DNA结构的刚性可以通过改变DNA中碱基对的数目来控制。虽然DNA双螺旋在生物系统如质粒和染色体中是一个灵活的聚合链,但这种灵活性并不适用于短的双链。dsDNA在大于其持久长度150bp(bp即碱基对,150bp约为50nm)时是非常灵活的,而dsDNA在短于其持久性长度时是刚性的。ssDNA的持久性长度约为1nm(对应1~3个碱基),大多数ssDNA是柔性的,可以很容易地形成环状和弯曲结构[18]。因此, 可以通过ssDNA和dsDNA的组合来设计所需DNA结构的柔性和刚性。例如,刚性的dsDNA结构可以由相对灵活的ssDNA链连接起来,形成具有理想拓扑结构的稳定装置[19]。DNA杂交可以在温度[20]、pH[21]和离子强度[22]等刺激下可逆地发生,dsDNA可以被分离成两个ssDNA(即“变性”),也可以重新组合成dsDNA(即“复性”)。利用这一可逆特性可以在某种条件下调控DNA的状态以实现机械功能。
从化学方面来说,DNA是水溶性的,因此DNA在水环境中可以通过多种化学反应进行修饰。5′端上的磷酸基和3′端上的羟基通常用于化学修饰。由于核酸酶的存在,DNA也可被生物降解,降解产物为核苷酸,是机体内自然发生的代谢物。因此DNA分子是无毒和生物相容的,这促进了其在体内外的应用[23]。此外,许多不同种类的酶为DNA装置提供了多功能性和可控性。例如,通过使用特定的酶,DNA可以被拉长、缩短,并在特定的位点连接。DNA纳米结构也可以通过简单的碱基配对与功能性核酸如核酸适配体[24]和分子信标[25]相互作用。
分子生物学为DNA的合成、操作和修饰提供了丰富的工具。例如,大块DNA可以从培养的生物体(如细菌或酵母)中纯化;带有所需序列的单个DNA片段可以保存在质粒中;较短的DNA序列(寡核苷酸)可以用化学方法从头合成。DNA也可以通过聚合酶链式反应(PCR)进行扩增,这使得DNA能够以较低并且在不断降低的成本购买到。除此之外,由于DNA可以与其他生物分子和金属纳米颗粒偶联,因此, DNA也可以用作模板来构建生物分子和纳米颗粒阵列[26]。
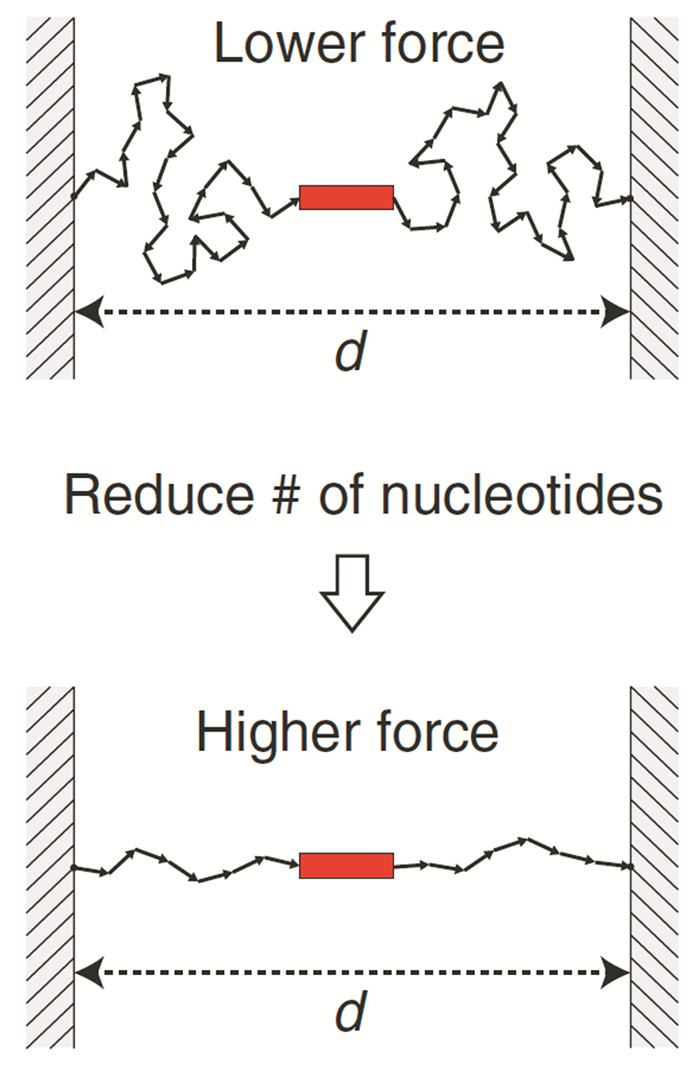
从生物方面来说,几乎所有的活细胞和组织都受到影响功能的机械力的作用,例如整合素受体在细胞骨架和细胞外基质之间双向传递力,细胞通过整合素受体来感知细胞外基质的机械特性。其中pN尺度的机械力在分子生物学、免疫学、再生医学等多个领域具有重要意义,而DNA机械技术特别适合测量和操控pN尺度的力。例如Nickels等[27]构建的DNA折纸钳是一种机械刚性支架,能够量化转录因子对DNA张力和染色体组织的敏感性。它利用ssDNA的熵弹簧特性,将目标分子的两端连接在一起,通过调整碱基数量改变ssDNA的长度,直接影响作用在研究系统上的熵力,从而对目标分子施加机械力(图 1)。然后用荧光显微镜监测目标分子,以揭示特定的力-函数关系。研究发现,当张力超过10pN时,转录因子TATA结合蛋白与DNA的关联被抑制,这为基因调控中机械力的重要性提供了证据。总之,没有任何其他材料能提供上述所有优势,这正是DNA能够构建多功能的、高度可操控的机械装置的重要原因。
图 1
1.2 DNA装置的构建
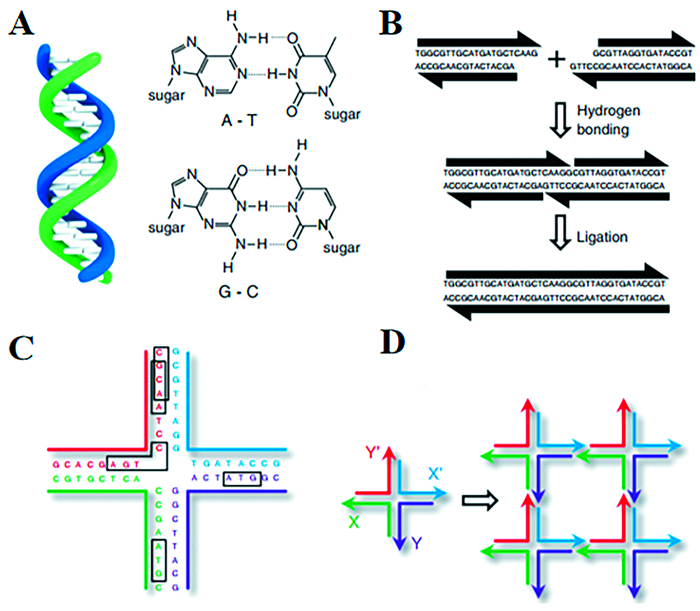
DNA材料的制备主要有两种组装过程:自组装和酶催化组装。最简单的DNA自组装方法是使用两个互补的ssDNA链自发杂交在一起形成一个dsDNA双螺旋(图 2(A))。这一过程是由几个非共价相互作用驱动的,包括氢键、静电、碱基堆积、疏水相互作用和范德华力,严格遵循Watson-Crick碱基配对规则。另一种自组装过程是利用dsDNA的粘性末端将两条dsDNA连接起来。在这种情况下,两条具有互补粘性末端的dsDNA链将杂交,并且缺口处可以被DNA连接酶封闭,从而形成一个单一的dsDNA(图 2(B))。
图 2
 图 2. DNA自组装的基本原理:(A)Watson-Crick碱基配对规则是指腺嘌呤(A)与胸腺嘧啶(T)配对,鸟嘌呤(G)与胞嘧啶(C)配对;(B)两个dsDNA可以通过粘性末端杂交连接,两个dsDNAs之间的缺口可以通过酶连接在一起,形成一个单一的dsDNA;(C)分支DNA基序必须具有不对称碱基对,才能稳定地作为DNA组装的组成部分;(D)分支DNA基序的粘性末端组装,左边显示一个带有四个粘性末端的分支DNA基序:X和X’互补,Y和Y’互补,四个分支DNA基序组合成一个四边形,如右图所示,外侧有额外的粘性末端,进一步使结构形成一个二维晶格Figure 2. Basic principles of DNA self-assembly: (A) The Watson-Crick base-paring rules mean that adenine (A) pairs with thymine (T) and guanine (G) pairs with cytosine (C); (B) Two double-stranded DNAs (dsDNA) can be connected by hybridization of sticky ends. The nick between the two dsDNAs can be chemically linked by enzyme ligation, resulting in a single dsDNA; (C) A branched DNA motif must have base-pair asymmetry to be stable enough to function as a component for DNA assembly; (D)Sticky-end assembly of branched DNA motifs. A branched DNA motif is shown on the left with four sticky ends: X complementary to X' and Y complementary to Y'. The four motifs assemble to form a quadrilateral, shown on the right, with further sticky ends on the outside, further allowing the structure to form a 2D lattice
图 2. DNA自组装的基本原理:(A)Watson-Crick碱基配对规则是指腺嘌呤(A)与胸腺嘧啶(T)配对,鸟嘌呤(G)与胞嘧啶(C)配对;(B)两个dsDNA可以通过粘性末端杂交连接,两个dsDNAs之间的缺口可以通过酶连接在一起,形成一个单一的dsDNA;(C)分支DNA基序必须具有不对称碱基对,才能稳定地作为DNA组装的组成部分;(D)分支DNA基序的粘性末端组装,左边显示一个带有四个粘性末端的分支DNA基序:X和X’互补,Y和Y’互补,四个分支DNA基序组合成一个四边形,如右图所示,外侧有额外的粘性末端,进一步使结构形成一个二维晶格Figure 2. Basic principles of DNA self-assembly: (A) The Watson-Crick base-paring rules mean that adenine (A) pairs with thymine (T) and guanine (G) pairs with cytosine (C); (B) Two double-stranded DNAs (dsDNA) can be connected by hybridization of sticky ends. The nick between the two dsDNAs can be chemically linked by enzyme ligation, resulting in a single dsDNA; (C) A branched DNA motif must have base-pair asymmetry to be stable enough to function as a component for DNA assembly; (D)Sticky-end assembly of branched DNA motifs. A branched DNA motif is shown on the left with four sticky ends: X complementary to X' and Y complementary to Y'. The four motifs assemble to form a quadrilateral, shown on the right, with further sticky ends on the outside, further allowing the structure to form a 2D lattice然而,这些线性DNA链不能组装成更复杂的纳米结构。因此,产生了分支DNA基序(branched DNA motifs),以扩大DNA装配的多样性。这样的分支序列在自然界中就可以找到,例如Holliday交叉(Holliday junction)是基因重组的中间产物[28, 29]。从结构来看,Holliday交叉的对称性导致了支点迁移,使得这个不稳定的基序不能用作构建稳定纳米结构的基本单元。为了解决这一问题,对各种不稳定的分支基序进行了综合设计和表征[30],例如稳定的不对称Holliday交叉(图 2(C))。因此,DNA进行自组装的基本前提是两端互补的粘性末端和不对称碱基配对的稳定支链DNA分子。图 2(D)给出了具有粘性末端的稳定支链DNA组装的过程:四个带有粘性末端的基元组装成一个四边形,外侧有额外的粘性末端,进一步形成二维晶格。然而,分支DNA交叉结构非常灵活,往往不能产生所需稳定的纳米结构。为了解决这个问题,Seeman等构建了一系列具有更强刚度的交叉结构,例如double-crossover(DX)[31~35]、triple-crossover(TX)[36~38]以及paranemic-crossover(PX)[39~41]。
DNA结构设计的另一个里程碑是Rothemund[42]提出的DNA折纸的概念,其增加了DNA组装的复杂性和多功能性。DNA折纸技术的基本思路是利用很多短DNA单链杂交的方式将一条长DNA单链折叠成特定的形状。这一技术吸引了来自化学、生物学、材料科学等不同领域科学家的广泛关注。短短几十年内,基于DNA折纸技术的研究已经取得了令人瞩目的发展。Rothemund利用折纸技术构建了令人眼花缭乱的纳米图形和纳米结构,包括五角星、笑脸等,充分展示了DNA折纸技术的优势。首先利用先进的软件设计出折纸结构的形状;其次设计交叉结构将DNA双螺旋结构固定在一起并按照特定方向排布,以此作为骨架;最后将骨架链和发挥“订书钉”作用的短链混合,在一定盐浓度下退火处理。此时数百条短链在骨架链上固定自己的位置,从而自组装形成多种多样的折纸结构。
随着研究的不断深入,利用DNA折纸技术得到的各种复杂纳米结构已经成为了DNA机械技术领域强有力的工具。Chao等[43]将机械连锁结构应用到DNA折纸中,组装了一个机械连锁的纳米肌肉。它能够模拟肌动蛋白和肌球蛋白在肌肉收缩和放松过程中的运动,它的长度可由加入燃料DNA链或反燃料DNA链调节,例如在添加燃料链(或反燃料链)后,纳米肌肉从约160nm延伸到310nm(或从310nm收缩到160nm)。通过粘性DNA链的功能化,纳米肌肉有望在今后的研究中用来控制分子的运动或产生力。
2. DNA机械装置
2.1 DNA机器人
机械装置的结构决定了机械运动的复杂性。通过设计出更加精巧的机械装置来产生更加复杂的机械行为或功能是DNA机械技术一个长期的目标。第一代DNA步行者可以在燃料链的驱动下移动[44]。2010年,Seeman等[45]报告了一个由三角形步行者和DNA折纸平台组成的系统。他们为系统提供了一个轨道和装载货物(金纳米颗粒)的站点。在添加燃料链(例如DNA链)后,三角形步行者将沿着轨道移动,并在站点装载预期的货物。额外的DNA链控制着步行者是否在每个站点装载货物。除了在线性轨道上,步行者也可以在沿规定轨道的二维平面上运动。所有的运动都是通过链置换反应进行的。该系统表明,通过DNA在结构和动力学上的可编程性,可以在纳米尺度上完成对分子运动的控制。
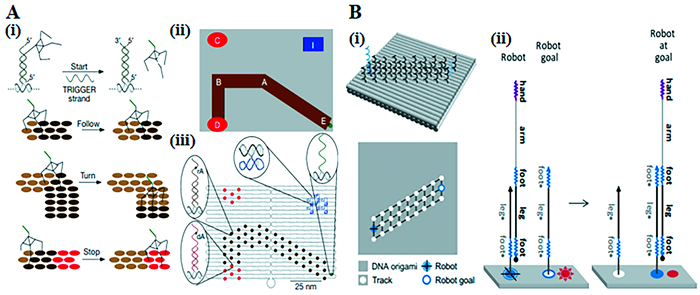
除了用链置换反应作为动力外,酶解法和DNA链杂交也可以驱动DNA机器人在轨道上的运动。Lund等[46]设计了一个可以执行启动、转弯和停止命令的DNA分子蜘蛛(图 3(A))。蜘蛛有一个链霉亲和素分子作为主体,三个脱氧核酶作为催化腿。蜘蛛在起始位置由触发链提供能量,并通过裂解衬底链行走,并驱动蜘蛛向剩余的衬底区域移动。一旦遇到无法裂解的链,蜘蛛的运动就停止了。此外,轨道可以被分割成一个网络,为机器人提供多条移动路线[47]。如果DNA机器人修饰有对光敏感的分子,它的运动就可以被光触发[48]。相对于在单个DNA折纸单元上对运动进行编程,Liber等[49]演示了DNA机器人在两个折纸单元之间来回运动。
图 3
 图 3. 结构复杂的DNA机械装置:(A) (i)DNA蜘蛛是由单链DNA通过链置换原理释放的,它遵循底物的轨迹,转弯,并继续到一个停止点;(ii)轨道EABD的示意图;(iii)DNA折纸二维示意图;(B)(i)双层DNA折纸三维和二维示意图;(ii)DNA机器人到达目标位置的机制Figure 3. The DNA mechanical device with complex configurations: (A) (i) A DNA spider is released by a single-stranded DNA using a strand displacement principle, and follows the substrate track, turns, and continues to a STOP site; (ii) Schematic of the track EABD; (iii) 2D schematic diagram of the DNA origami; (B) (i) 3D and 2D schematic diagrams of the double-layer DNA origami; (ii) Mechanism of the robot reaching a goal location
图 3. 结构复杂的DNA机械装置:(A) (i)DNA蜘蛛是由单链DNA通过链置换原理释放的,它遵循底物的轨迹,转弯,并继续到一个停止点;(ii)轨道EABD的示意图;(iii)DNA折纸二维示意图;(B)(i)双层DNA折纸三维和二维示意图;(ii)DNA机器人到达目标位置的机制Figure 3. The DNA mechanical device with complex configurations: (A) (i) A DNA spider is released by a single-stranded DNA using a strand displacement principle, and follows the substrate track, turns, and continues to a STOP site; (ii) Schematic of the track EABD; (iii) 2D schematic diagram of the DNA origami; (B) (i) 3D and 2D schematic diagrams of the double-layer DNA origami; (ii) Mechanism of the robot reaching a goal location到目前为止,DNA机器人已经从非自主发展到自主,从按控制方向行走发展到转弯、选分支、捡货物,甚至可以进行药物递送。例如,为了在轨道上进行随机行走,一个单链DNA机器人被设计成两个足域各有6个核苷酸,一个腿域有15个核苷酸。轨道是由许多单链构成的,延伸在二维DNA折纸表面。每条轨道链通过碱基互补与机器人的一个足部和腿部区域结合,使机器人的另一个足部区域处于自由状态。自由足域首先与相邻轨道的互补足域结合。机器人通过可逆的链置换反应从轨道上的一个位置移动到另一个位置。此外,Thubagere等[50]开发了一个能在DNA折纸表面进行货物分类的机器人(图 3(B))。首先机器人随机行走,如果它撞上了货物,机器人会把它捡起来,继续随机行走。如果它撞上了一个目标,也就是货物的指定目的地,机器人就会把它放下。然后,机器人将重复这个过程,直到所有的货物被分类。除此之外,DNA机器人也有潜力成为智能药物输送系统,对分子触发器作出反应。Li等[51]利用DNA折纸技术构建了一个自主的DNA机器人,它被用于运输有效载荷,并将其专门用于肿瘤治疗。随着DNA机械技术的发展,DNA机器人所取得的成就令人瞩目。研究人员能够设计出动态的DNA系统,这些系统有能力在未来执行更复杂的动作。
2.2 DNA张力传感器
机械力在细胞粘附、免疫识别、转移和凝血等过程中起重要作用[52~55]。例如,整合素受体在细胞骨架和细胞外基质(ECM)之间双向传递机械力。研究这些力有几种不同的方法。传统的SMFS技术(包括原子力谱、磁镊和光镊)被用来探测细胞受体与其配体之间的相互作用[9]。这些方法可捕获配体-受体相互作用的物理化学过程,但往往不能很好地描述生物学过程,因为许多膜受体是在低聚物中起作用的,而不是作为孤立的分子起作用。例如,整合素组装成黏着斑(FAs),其中包含数百种不同的结构和信号蛋白,它们在物理上将ECM与细胞骨架连接起来[56]。另一种研究细胞机械生物学的方法是牵引力显微镜(TFM),它测量细胞驱动的聚合物基板形变[57]。TFM具有微米级的空间分辨率和纳牛级的灵敏度,能够描绘出成熟FAs所产生的力,但不能解决成熟FAs形成之前的新生粘连或局灶性复合物所产生的力。
为了弥补以上方法的不足,人们尝试利用经改造的DNA张力传感器来对机械力进行研究。和宏观测力仪一样,DNA张力传感器也包括两个主要部分:一个是在外力作用下延伸的弹簧,另一个是报告弹簧延伸情况的量尺。在现有的DNA张力传感器中,一个茎环发夹作为弹簧,荧光猝灭报告发夹的延伸。每个传感器的一端固定在表面上,另一端是与细胞受体结合的配体。在荧光图像中,荧光强度与单位面积内展开的发夹数量成正比。
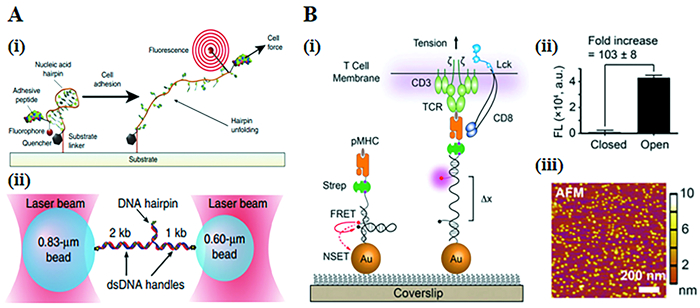
2014年,Blakely等[58]在DNA发夹的基础上设计了一种新的分子张力探针,它能在时空上报告牵引力(图 4(A))。发夹的5′端与整合素粘附受体结合的GGRGDS肽连接,3′端经化学偶联到细胞培养基上。DNA发夹能够通过改变DNA序列的长度和组成,在精确的力的作用下展开[59]。将不同长度和序列的DNA发夹连接到不同的荧光团-猝灭剂上,荧光团可以在折叠状态下猝灭,而在未折叠状态下发出荧光。
图 4
 图 4. (A) 基于DNA发夹的分子张力探针的设计与表征:(i)张力探针的示意图;(ii)用于表征发夹力学的实验几何示意图;(B)基于DNA发夹的金纳米颗粒张力传感器的设计与表征:(i)基于DNA的AuNP传感器的原理图;(ii)图中显示发夹闭合和打开形态的荧光强度;(iii)在玻璃上固定的AuNP传感器的AFM图像Figure 4. (A) Design and characterization of DNA hairpin force probe: (i) Schematic depiction of the tension probes; (ii) Schematic of the experimental geometry used to characterize the mechanics of the hairpins; (B) Design and characterization of AuNP tension sensor based on DNA hairpin: (i) Schematic of DNA-based AuNP sensor; (ii) Plot showing the fluorescence intensity of the closed and open forms of the hairpin; (iii)The AFM image of the immobilized AuNP sensors on a glass coverslip
图 4. (A) 基于DNA发夹的分子张力探针的设计与表征:(i)张力探针的示意图;(ii)用于表征发夹力学的实验几何示意图;(B)基于DNA发夹的金纳米颗粒张力传感器的设计与表征:(i)基于DNA的AuNP传感器的原理图;(ii)图中显示发夹闭合和打开形态的荧光强度;(iii)在玻璃上固定的AuNP传感器的AFM图像Figure 4. (A) Design and characterization of DNA hairpin force probe: (i) Schematic depiction of the tension probes; (ii) Schematic of the experimental geometry used to characterize the mechanics of the hairpins; (B) Design and characterization of AuNP tension sensor based on DNA hairpin: (i) Schematic of DNA-based AuNP sensor; (ii) Plot showing the fluorescence intensity of the closed and open forms of the hairpin; (iii)The AFM image of the immobilized AuNP sensors on a glass coverslip2016年Liu等[60]开发了基于DNA的金纳米颗粒张力传感器(图 4(B)),可以直接对T细胞激活过程中单个T细胞受体(TCR)和抗原肽-MHC分子复合物(pMHC)结合时的机械力进行成像和量化。这个传感器具有较高的空间和时间分辨率,可以发现小鼠CD8+ T细胞通过其T细胞受体(TCRs)施加12~19 pN的张力对抗原进行识别,这支持了TCR利用力来区分外来抗原和自身抗原的理论。此外还发现,TCR力增强到大于19pN,淋巴细胞功能相关抗原1(LFA-1)介导的粘附会调节TCR-pMHC张力并进行空间重组,这说明了在受体机制中协同作用的重要性。
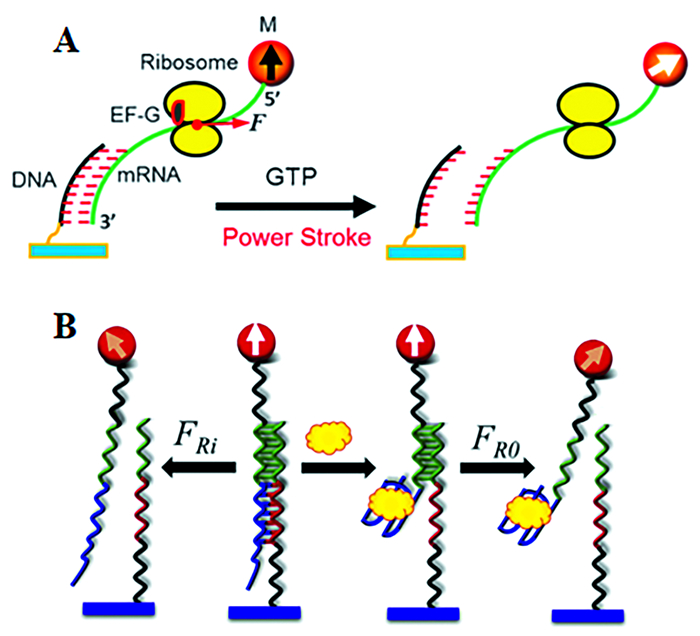
近年来,Dutta等[61]利用DNA折纸技术构建了DNA折纸张力探针(DOTPs),它可以并行地将粘附肽和力敏感发夹复合在一起,用来描绘由活细胞产生的pN力。DNA张力传感器能够对生命系统中的pN力实时成像,这对机械生物学产生了重大影响。除此之外,Brockman等[62]将荧光偏振(FP)技术与DNA张力探针相结合,使分子力显微镜(MFM)成为可能,能够在pN分辨率下揭示细胞牵引力的方向。Yao等[63]基于力诱导的剩磁谱(Force-induced remanence magnetic spectroscopy,FIRMS)构建的DNA-RNA双链张力传感器能够测量运动蛋白EF-G在核糖体转位过程中产生的机械力,并且证明了EF-G和GTP结合导致转位的机制是EF-G动力冲程引起核糖体的转位(图 5(A))。Lu等[64]借助核酸适配体构建双链DNA张力传感器,利用FIRMS技术测量免标记蛋白与其核酸适配体之间的相互作用,并且能够实现靶蛋白宽范围高灵敏的检测(图 5(B))。尽管如此,测量罕见的且存在时间很短的机械力仍然是一个挑战。Ma等[65]开发了一种可以存储与机械力相关信息的新型探测器,使得测量不频繁或短暂的TCR力的能力有了指数级的提高,这将为机械免疫学提供更深入的见解。
图 5
 图 5. (A) DNA-RNA双链张力传感器测量运动蛋白EF-G在核糖体转位过程中产生的机械力示意图;(B)DNA张力传感器检测凝血酶蛋白的示意图Figure 5. (A)A DNA-RNA duplexes tension sensor measures the mechanical forces generated by the motor protein (EF-G) during ribosomal translocation; (B) The detection of thrombin proteins by DNA tension sensors
图 5. (A) DNA-RNA双链张力传感器测量运动蛋白EF-G在核糖体转位过程中产生的机械力示意图;(B)DNA张力传感器检测凝血酶蛋白的示意图Figure 5. (A)A DNA-RNA duplexes tension sensor measures the mechanical forces generated by the motor protein (EF-G) during ribosomal translocation; (B) The detection of thrombin proteins by DNA tension sensors基于DNA的张力传感器的一个缺陷是它们对DNA酶降解很敏感。在细胞力成像中,这些DNA酶会损伤传感器,降低信噪比,甚至产生假信号。Zhao等[66]测试了肽核酸(PNA)、化学修饰的RNA及其与DNA的杂交物作为构建张力传感器的替代生物材料,对四个双链体dsDNA、PNA/DNA、dsRNA(修饰的RNA)和PNA/RNA的DNA酶抗性、细胞力成像能力和材料坚固性进行了评估。结果显示,PNA/DNA杂交是合成耐DNA酶张力传感器的一种可行材料,该传感器保留了机械力报告能力,并在表达DNA酶的细胞中保持稳定。此外,力介导的受体信号通常涉及多个相互作用的共受体(与TCR一样),DNA折纸张力传感器虽然可以同时与多个配体结合[61],但尚未用于研究多种相互作用。
2.3 其他DNA机械装置
DNA分子已被证明是设计和建造机械分子装置的优良材料,当暴露于外部信号时,这些机械分子装置能够感知、驱动和发挥功能。近年来各种由DNA分子构成的机械装置被设计合成,除了上述的DNA机器人和DNA张力传感器外,DNA镊子、DNA曲柄滑块、DNA旋转装置等也是DNA机械装置的重要组成部分。
2000年的一篇开创性论文引入了DNA链置换反应的概念,这对许多DNA机器的构建至关重要[67]。在这个概念中,通过添加DNA燃料链关闭DNA镊子,然后用不同的DNA链重新打开,通过测量两个染料之间的荧光能量转移来测量机械运动(图 6(A))。除此之外,DNA纳米机器也可以利用DNA链杂交释放的能量产生机械力,或者增强纳米结构的形状变化。例如,Powell等[68]构建了具有可编程结构转换能力的大型DNA折纸轮烷,并进一步分析了两种由DNA杂交介导的轮烷结构转换行为。另外,Kuzuya等[69]设计了一种具有目标识别位点的DNA镊子,可以用作单分子信标(图 6(B))。DNA纳米机械装置的形状转变,如镊子的关闭或打开,可以通过与目标分子的相互作用被选择性地触发,并使用原子力显微镜成像来可视化。由于可以使用多种相互作用,例如蛋白质-配体、DNA-RNA或DNA-金属离子等,这种传感方法适用于多种靶标。
图 6
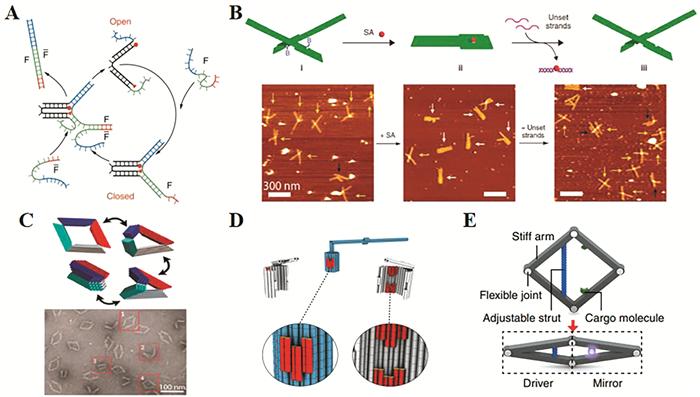
 图 6. (A) DNA镊子,在打开状态时(顶部),两个荧光团是分开的;链F将蓝色和绿色的链连接在一起,关闭镊子,增加荧光共振能量转移(FRET);链F*通过链置换反应移除链F,重新打开DNA镊子;(B)用DNA折纸镊子检测小分子;(C)具有Bennett连杆机构四杆DNA机器,可以在闭合状态和打开状态之间通过复杂的三维运动路径;(D)基于DNA旋转装置的设计。蓝色:转子单元;灰色:夹具单元;红色:插座单元;(E)DNA纳米开关的三维模型,通过调整两个货物分子之间的距离来控制它们之间的相互作用。Figure 6. (A) A DNA tweezer. In the open form (top), the two fluorophores are separated. Strand F brings the blue and green strands together and closes the tweezer, increasing fluorescence resonance energy transfer (FRET). Strand F* removes F by strand displacement, restoring the open form of the tweezer. (B)Detection of small molecule with DNA origami tweezers. (C)A four-bar DNA machine that has Bennett linkages and that can traverse a complex 3D motion path between closed and opened states. (D) Design of a DNA-based rotary apparatus. Blue: rotor unit; gray: clamp units; red: socket units. (E) Three-dimensional models of the DNA nanoswitch that controls the interaction between two cargo molecules by adjusting the distance between them.
图 6. (A) DNA镊子,在打开状态时(顶部),两个荧光团是分开的;链F将蓝色和绿色的链连接在一起,关闭镊子,增加荧光共振能量转移(FRET);链F*通过链置换反应移除链F,重新打开DNA镊子;(B)用DNA折纸镊子检测小分子;(C)具有Bennett连杆机构四杆DNA机器,可以在闭合状态和打开状态之间通过复杂的三维运动路径;(D)基于DNA旋转装置的设计。蓝色:转子单元;灰色:夹具单元;红色:插座单元;(E)DNA纳米开关的三维模型,通过调整两个货物分子之间的距离来控制它们之间的相互作用。Figure 6. (A) A DNA tweezer. In the open form (top), the two fluorophores are separated. Strand F brings the blue and green strands together and closes the tweezer, increasing fluorescence resonance energy transfer (FRET). Strand F* removes F by strand displacement, restoring the open form of the tweezer. (B)Detection of small molecule with DNA origami tweezers. (C)A four-bar DNA machine that has Bennett linkages and that can traverse a complex 3D motion path between closed and opened states. (D) Design of a DNA-based rotary apparatus. Blue: rotor unit; gray: clamp units; red: socket units. (E) Three-dimensional models of the DNA nanoswitch that controls the interaction between two cargo molecules by adjusting the distance between them.为了实现更复杂的机械运动和结构,Marras等[70]通过宏观机械装置的设计原理来构建DNA纳米机械装置。他们成功搭建了包括铰链和曲柄滑块的能够执行复杂三维动作的一组DNA装置(图 6(C))。两个不同运动范围和刚度的滑块由ssDNAs的长度控制,短连接的设计类似于刚度为0.42pN/nm的线性弹簧,而长连接的设计则表现出非线性力。接下来,设计了一个DNA装置,它由两个通过支架沿边缘连接的刚性臂组成,用来进行纯旋转运动。从理论上讲,旋转运动和线性运动可以结合起来进行任何复杂的运动。2016年,Dietz等[71]构建了一个纳米尺度的旋转装置,该装置能够在不需要任何能量输入的情况下连续旋转(图 6(D))。其由三个DNA折纸部分(分别作为转子单元、夹具单元和插座单元)通过碱基堆积或粘性末端杂交的相互作用组装而成。根据旋转装置中使用条件的改变,转子可以停在对接点,也可以围绕中心轴旋转进行随机行走。
2016年,Ke等[72]设计了一种DNA纳米开关(图 6(E))。该装置由4个以菱形连接的刚性臂组成,这种设计使得在一半的菱形中产生的精确距离变化可以定量地传播到另一半的菱形中。两个不同的机制被用来驱动该装置。一种机制是增加额外的结构锁定链,形成两个刚性的DNA双螺旋,其决定连接位点之间的距离和菱形的角度。通过将增强型绿色荧光蛋白(eGFP)连接到DNA纳米器件上,构建了具有可调荧光性质的DNA-蛋白杂交分子。分子的构象可通过改变“连接支柱”的长度来精确控制,进而开启和关闭荧光信号。第二种机制是一个压缩弹簧,通过DNA菱形顶部和底部铰链上的锁定机制来维持其关闭状态。这一新的机制能够建立一组传感器,对广泛的刺激包括缓冲成分、限制性内切酶的活性和RNA-DNA杂交的改变等作出反应。DNA纳米开关可以作为研究单分子水平上的弱分子相互作用(例如,弱蛋白-蛋白相互作用)的通用平台。另外,该装置在特定目标存在时的剧烈构象变化可以通过原子力显微镜或电子显微镜在单分子水平上直观地检测到。
3. 总结与展望
近年来DNA机械技术得到了广泛的关注和深入的研究。DNA机械装置适合测量和操控pN尺度的力,这个尺度的机械力在分子生物学、免疫学、再生医学、材料科学等多个领域都具有重要意义。DNA独特的分子结构和性质是构建DNA机械装置的力学基础。除此之外,DNA折纸技术更是成为了研究DNA机械技术的强有力工具。
DNA机械装置的结构决定了机械运动的复杂性。到目前为止,DNA机器人已经从非自主发展到自主,从按控制方向行走发展到转弯、选分支、捡货物,甚至发展了专门用于肿瘤的智能药物递送系统。除此之外,机械力在细胞粘附、免疫识别、转移和凝血等过程中起重要作用。DNA张力传感器正开始对这些机械力进行研究。此外,DNA镊子、DNA曲柄滑块、DNA旋转装置等也是DNA机械装置的重要组成部分。
DNA机械技术仍然存在许多挑战,包括结构预测的局限性,复杂结构的组装率低,在低离子强度、高温或非水环境中的稳定性差。目前的研究正在致力于解决这些问题。其中一项工作涉及到共价交联DNA折纸,它最初是为了在高温下稳定DNA纳米结构而提出的[73],现在其也可用于扩大各种DNA机械装置力的范围。由于合成生物学、基因组学和个性化医学的需求不断增长,DNA制造的成本可能会继续下降,这也会进一步促进DNA机械技术的广泛研究与应用。也许在未来,DNA机械技术可以通过对病人细胞的机械特性进行个性化的医学研究,进而推动这些领域的进步。从广义上讲,DNA机械技术领域的装置和工具不应局限于基于核酸的设计,还可以扩展到基于蛋白质、脂类和有机聚合物等替代性纳米材料来进行设计。
-
-
[1]
Seeman N C. J. Theor. Biol., 1982, 99: 237-247. doi: 10.1016/0022-5193(82)90002-9
-
[2]
Luo D. Mater. Today, 2003, 6: 38-43.
-
[3]
LaBean T H, Li H Y. Nano Today, 2007, 2: 26-35. doi: 10.1016/S1748-0132(07)70056-7
-
[4]
Seeman N C. Nature, 2003, 421: 427-431. doi: 10.1038/nature01406
-
[5]
Seeman N C, Lukeman P S. Rep. Prog. Phys., 2005, 68: 237-270. doi: 10.1088/0034-4885/68/1/R05
-
[6]
Lin C, Liu Y, Yan H. Biochemistry, 2009, 48: 1663-1674. doi: 10.1021/bi802324w
-
[7]
Feldkamp U, Niemeyer C M. Angew. Chem. Int. Ed., 2006, 45: 1856-1876. doi: 10.1002/anie.200502358
-
[8]
Blanchard A T, Bazrafshan A S, Yi J, et al. Nano Lett., 2019, 19: 6977-6986. doi: 10.1021/acs.nanolett.9b02311
-
[9]
Neuman K C, Nagy A. Nat. Methods, 2008, 5: 491-505. doi: 10.1038/nmeth.1218
-
[10]
Kilchherr F, Wachauf C, Pelz B, et al. Science, 2016, 353: aaf5508.
-
[11]
Han D, Pal S, Nangreave J, et al. Science, 2011, 332: 342-346. doi: 10.1126/science.1202998
-
[12]
Langecker M, Arnaut V, Martin T G, et al. Science, 2012, 338: 932-936. doi: 10.1126/science.1225624
-
[13]
Douglas S M, Bachelet I, Church G M. Science, 2012, 335: 831-834. doi: 10.1126/science.1214081
-
[14]
Douglas S M, Chou J J, Shih W M. PNAS, 2007, 104: 6644-6648. doi: 10.1073/pnas.0700930104
-
[15]
Ding B, Deng Z, Yan H, et al. J. Am. Chem. Soc., 2010, 132: 3248-3258. doi: 10.1021/ja9101198
-
[16]
Maune H T, Han S, Barish R D, et al. Nat. Nanotechnol., 2010, 5: 61-66. doi: 10.1038/nnano.2009.311
-
[17]
Derr N D, Goodman B S, Jungmann R, et al. Science, 2012, 338: 662-665. doi: 10.1126/science.1226734
-
[18]
Tinland B, Pluen A, Sturm J, et al. Macromolecules, 1997, 30: 5763-5765. doi: 10.1021/ma970381+
-
[19]
Yang D, Campolongo M J, Tran T N N, et al. DNA materials and their applications. Wiley Interdiscip. Rev-Nanomed Nanobiotechnol., 2010, 2: 648-669. doi: 10.1002/wnan.111
-
[20]
Zhang K, Zhu X, Jia F, et al. J. Am. Chem. Soc., 2013, 135: 14102-14105. doi: 10.1021/ja408465t
-
[21]
Amodio A, Adedeji A F, Castronovo M, et al. J. Am. Chem. Soc., 2016, 138: 12735-12738. doi: 10.1021/jacs.6b07676
-
[22]
Zhu B, Zhao Y, Dai J B, et al. ACS Appl. Mater. Interf., 2017, 9: 18434-18439. doi: 10.1021/acsami.7b04784
-
[23]
Surana S, Shenoy A R, Krishnan Y. Nat. Nanotechnol., 2015, 10: 741-747. doi: 10.1038/nnano.2015.180
-
[24]
Zhu G, Zheng J, Song E, et al. PNAS, 2013, 110: 7998-8003. doi: 10.1073/pnas.1220817110
-
[25]
Xie N, Huang J, Yang X, et al. Chem. Commun., 2016, 52: 2346-2349. doi: 10.1039/C5CC09980C
-
[26]
Niemeyer C M, Ceyhan B, Hazarika P. Angew. Chem. Int. Ed., 2003, 42: 5766-5770. doi: 10.1002/anie.200352744
-
[27]
Nickels P C, Wunsch B, Holzmeister P, et al. Science, 2016, 354: 305-307. doi: 10.1126/science.aah5974
-
[28]
Holliday R. Genet. Res., 1964, 5: 282-290. doi: 10.1017/S0016672300001233
-
[29]
Meselson M S, Radding C M. PNAS, 1975, 72: 358-361. doi: 10.1073/pnas.72.1.358
-
[30]
Lin C X, Liu Y, Rinker S, et al. Chem. Phys. Chem., 2006, 7: 1641-1647. doi: 10.1002/cphc.200600260
-
[31]
Winfree E, Liu F R, Wenzler L A, et al. Nature, 1998, 394: 539-544. doi: 10.1038/28998
-
[32]
Fu T J, Seeman N C. Biochemistry, 1993, 32: 3211-3220. doi: 10.1021/bi00064a003
-
[33]
Li X J, Yang X P, Qi J, et al. J. Am. Chem. Soc., 1996, 118: 6131-6140. doi: 10.1021/ja960162o
-
[34]
Liu F R, Sha R J, Seeman N C. J. Am. Chem. Soc., 1999, 121: 917-922. doi: 10.1021/ja982824a
-
[35]
Reishus D, Shaw B, Brun Y, et al. J. Am. Chem. Soc., 2005, 127: 17590-17591. doi: 10.1021/ja0557177
-
[36]
LaBean T H, Yan H, Kopatsch J, et al. J. Am. Chem. Soc., 2000, 122: 1848-1860. doi: 10.1021/ja993393e
-
[37]
Liu Y, Lin C X, Li H Y, et al. Angew. Chem. Int. Ed., 2005, 44: 4333-4338. doi: 10.1002/anie.200501089
-
[38]
Liu D, Park S H, Reif J H, et al. PNAS, 2004, 101: 717-722. doi: 10.1073/pnas.0305860101
-
[39]
Yan H, Zhang X P, Shen Z Y, et al. Nature, 2002, 415: 62-65. doi: 10.1038/415062a
-
[40]
Shen Z Y, Yan H, Wang T, et al. J. Am. Chem. Soc., 2004, 126: 1666-1674. doi: 10.1021/ja038381e
-
[41]
Liu W Y, Wang X, Wang T, et al. Nano Lett., 2008, 8: 317-322. doi: 10.1021/nl072803r
-
[42]
Rothemund P W. Nature, 2006, 440: 297-302. doi: 10.1038/nature04586
-
[43]
ChaoY C, Hong Y J, Lee C Y, et al. Nanoscale, 2020, 12: 2992-2998. doi: 10.1039/C9NR06314E
-
[44]
Shin J S, Pierce N A. J. Am. Chem. Soc., 2004, 126: 10834-10835. doi: 10.1021/ja047543j
-
[45]
Gu H, Chao J, Xiao S J, et al. Nature, 2010, 465: 202-205. doi: 10.1038/nature09026
-
[46]
Lund K, Manzo A J, Dabby N, et al. Nature, 2010, 465: 206-210. doi: 10.1038/nature09012
-
[47]
Wickham S F, Bath J, Katsuda Y, et al. Nat. Nanotechnol., 2012, 7: 169-173. doi: 10.1038/nnano.2011.253
-
[48]
Yang Y, Goetzfried M A, Hidaka K, et al. Nano Lett., 2015, 15: 6672-6676. doi: 10.1021/acs.nanolett.5b02502
-
[49]
Liber M, Tomov T E, Tsukanov R, et al. Small, 2015, 11: 568-575. doi: 10.1002/smll.201402028
-
[50]
Thubagere A J, Li W, Johnson R F, et al. Science, 2017, 357: eaan6558.
-
[51]
Li S P, Jiang Q, Liu S L, et al. Nat. Biotechnol., 2018, 36: 258-261. doi: 10.1038/nbt.4071
-
[52]
Evans E A, Calderwood D A. Science, 2007, 316: 1148-1153. doi: 10.1126/science.1137592
-
[53]
Levental K R, Yu H, Kass L, et al. Cell, 2009, 139: 891-906. doi: 10.1016/j.cell.2009.10.027
-
[54]
Kim S T, Takeuchi K, Sun Z Y, et al. J. Biol. Chem., 2009, 284: 31028-31037. doi: 10.1074/jbc.M109.052712
-
[55]
Qiu Y, Brown A C, Myers D R, et al. PNAS, 2014, 111: 14430-14435. doi: 10.1073/pnas.1322917111
-
[56]
Kanchanawong P, G Shtengel, Pasapera A M, et al. Nature, 2010, 468: 580-U262. doi: 10.1038/nature09621
-
[57]
Schwarz U S, Soine J R D. Biochim. Biophys. Acta-Mol. Cell Res., 2015, 1853: 3095-3104. doi: 10.1016/j.bbamcr.2015.05.028
-
[58]
Blakely B L, Dumelin C E, Trappmann B, et al. Nat. Methods, 2014, 11: 1229-1241. doi: 10.1038/nmeth.3145
-
[59]
KongF, García A J, Mould A P, et al. J. Cell Biol., 2009, 185: 1275-1284. doi: 10.1083/jcb.200810002
-
[60]
Liu Y, Blanchfield L, Ma V P, et al. PNAS, 2016, 113: 5610-5615. doi: 10.1073/pnas.1600163113
-
[61]
Dutta P K, Zhang Y, Blanchard A T, et al. Nano Lett., 2018, 18: 4803-4811. doi: 10.1021/acs.nanolett.8b01374
-
[62]
Ma R, Kellner A V, Ma V P Y, et al. PNAS, 2019, 116: 16949-16954. doi: 10.1073/pnas.1904034116
-
[63]
Yao L, Li Y, W T Tsai, et al. Angew. Chem. Int. Ed., 2013, 52: 14041-14044. doi: 10.1002/anie.201307419
-
[64]
Lu P, Zhang D, Chai Y H, et al. Anal. Chim. Acta, 2019, 1045: 1-9. doi: 10.1016/j.aca.2018.09.019
-
[65]
Brockman J M, Blanchard A T, Ma V P, et al. Nat. Methods, 2018, 15: 115-117. doi: 10.1038/nmeth.4536
-
[66]
Zhao Y C, Sarkar A, Wang X F. Biosens. Bioelectron., 2020, 150: 111959. doi: 10.1016/j.bios.2019.111959
-
[67]
Yurke B, Turberfield A J, Mills A P, et al. Nature, 2000, 406: 605-608. doi: 10.1038/35020524
-
[68]
Powell J T, Akhuetie-Oni B O, Zhang Z, et al. Angew. Chem. Int. Ed., 2016, 55: 11412-11416. doi: 10.1002/anie.201604621
-
[69]
Kuzuya A, Sakai Y, Yamazaki T, et al. Nat. Commun., 2011, 2: 449. doi: 10.1038/ncomms1452
-
[70]
Marras A E, Zhou L, Su H J, et al. PNAS, 2015, 112: 713-718. doi: 10.1073/pnas.1408869112
-
[71]
Ketterer P, Willner E M, Dietz H, et al. Sci. Adv., 2016, 2: e1501209.
-
[72]
Ke Y, Meyer T, Shih W M, et al. Nat. Commun., 2016, 7: 10935. doi: 10.1038/ncomms10935
-
[73]
Gerling T, Kube M, Kick B et al. Sci. Adv., 2018, 4: eaau1157.
-
[1]
-

图 2 DNA自组装的基本原理:(A)Watson-Crick碱基配对规则是指腺嘌呤(A)与胸腺嘧啶(T)配对,鸟嘌呤(G)与胞嘧啶(C)配对;(B)两个dsDNA可以通过粘性末端杂交连接,两个dsDNAs之间的缺口可以通过酶连接在一起,形成一个单一的dsDNA;(C)分支DNA基序必须具有不对称碱基对,才能稳定地作为DNA组装的组成部分;(D)分支DNA基序的粘性末端组装,左边显示一个带有四个粘性末端的分支DNA基序:X和X’互补,Y和Y’互补,四个分支DNA基序组合成一个四边形,如右图所示,外侧有额外的粘性末端,进一步使结构形成一个二维晶格
Figure 2 Basic principles of DNA self-assembly: (A) The Watson-Crick base-paring rules mean that adenine (A) pairs with thymine (T) and guanine (G) pairs with cytosine (C); (B) Two double-stranded DNAs (dsDNA) can be connected by hybridization of sticky ends. The nick between the two dsDNAs can be chemically linked by enzyme ligation, resulting in a single dsDNA; (C) A branched DNA motif must have base-pair asymmetry to be stable enough to function as a component for DNA assembly; (D)Sticky-end assembly of branched DNA motifs. A branched DNA motif is shown on the left with four sticky ends: X complementary to X' and Y complementary to Y'. The four motifs assemble to form a quadrilateral, shown on the right, with further sticky ends on the outside, further allowing the structure to form a 2D lattice
图 3 结构复杂的DNA机械装置:(A) (i)DNA蜘蛛是由单链DNA通过链置换原理释放的,它遵循底物的轨迹,转弯,并继续到一个停止点;(ii)轨道EABD的示意图;(iii)DNA折纸二维示意图;(B)(i)双层DNA折纸三维和二维示意图;(ii)DNA机器人到达目标位置的机制
Figure 3 The DNA mechanical device with complex configurations: (A) (i) A DNA spider is released by a single-stranded DNA using a strand displacement principle, and follows the substrate track, turns, and continues to a STOP site; (ii) Schematic of the track EABD; (iii) 2D schematic diagram of the DNA origami; (B) (i) 3D and 2D schematic diagrams of the double-layer DNA origami; (ii) Mechanism of the robot reaching a goal location
图 4 (A) 基于DNA发夹的分子张力探针的设计与表征:(i)张力探针的示意图;(ii)用于表征发夹力学的实验几何示意图;(B)基于DNA发夹的金纳米颗粒张力传感器的设计与表征:(i)基于DNA的AuNP传感器的原理图;(ii)图中显示发夹闭合和打开形态的荧光强度;(iii)在玻璃上固定的AuNP传感器的AFM图像
Figure 4 (A) Design and characterization of DNA hairpin force probe: (i) Schematic depiction of the tension probes; (ii) Schematic of the experimental geometry used to characterize the mechanics of the hairpins; (B) Design and characterization of AuNP tension sensor based on DNA hairpin: (i) Schematic of DNA-based AuNP sensor; (ii) Plot showing the fluorescence intensity of the closed and open forms of the hairpin; (iii)The AFM image of the immobilized AuNP sensors on a glass coverslip
图 5 (A) DNA-RNA双链张力传感器测量运动蛋白EF-G在核糖体转位过程中产生的机械力示意图;(B)DNA张力传感器检测凝血酶蛋白的示意图
Figure 5 (A)A DNA-RNA duplexes tension sensor measures the mechanical forces generated by the motor protein (EF-G) during ribosomal translocation; (B) The detection of thrombin proteins by DNA tension sensors
图 6 (A) DNA镊子,在打开状态时(顶部),两个荧光团是分开的;链F将蓝色和绿色的链连接在一起,关闭镊子,增加荧光共振能量转移(FRET);链F*通过链置换反应移除链F,重新打开DNA镊子;(B)用DNA折纸镊子检测小分子;(C)具有Bennett连杆机构四杆DNA机器,可以在闭合状态和打开状态之间通过复杂的三维运动路径;(D)基于DNA旋转装置的设计。蓝色:转子单元;灰色:夹具单元;红色:插座单元;(E)DNA纳米开关的三维模型,通过调整两个货物分子之间的距离来控制它们之间的相互作用。
Figure 6 (A) A DNA tweezer. In the open form (top), the two fluorophores are separated. Strand F brings the blue and green strands together and closes the tweezer, increasing fluorescence resonance energy transfer (FRET). Strand F* removes F by strand displacement, restoring the open form of the tweezer. (B)Detection of small molecule with DNA origami tweezers. (C)A four-bar DNA machine that has Bennett linkages and that can traverse a complex 3D motion path between closed and opened states. (D) Design of a DNA-based rotary apparatus. Blue: rotor unit; gray: clamp units; red: socket units. (E) Three-dimensional models of the DNA nanoswitch that controls the interaction between two cargo molecules by adjusting the distance between them.
-

 下载:
下载:

 下载:
下载:





 扫一扫看文章
扫一扫看文章
计量
- PDF下载量: 0
- 文章访问数: 0
- HTML全文浏览量: 0
 下载:
下载:
