 图 1
光合作用理想状态下能量分布
Figure 1.
Energy distribution of photosynthesis under ideal state
图 1
光合作用理想状态下能量分布
Figure 1.
Energy distribution of photosynthesis under ideal state
引用本文:
孙增森, 沈蔡龙, 李敏, 张祥祥, 杨宇, 刘畅. 光合作用的热力学研究进展[J]. 化学通报,
2017, 80(6): 517-523.

Citation: Sun Zengsen, Shen Cailong, Li Min, Zhang Xiangxiang, Yang Yu, Liu Chang. Research Progress in the Thermodynamic Analysis of Photosynthesis[J]. Chemistry, 2017, 80(6): 517-523.
Citation: Sun Zengsen, Shen Cailong, Li Min, Zhang Xiangxiang, Yang Yu, Liu Chang. Research Progress in the Thermodynamic Analysis of Photosynthesis[J]. Chemistry, 2017, 80(6): 517-523.
光合作用的热力学研究进展
English
Research Progress in the Thermodynamic Analysis of Photosynthesis
Abstract:
The thermodynamic analysis of photosynthesis efficiency is critical to understand the photosynthesis mechanism, and also an important approach to achieve artificial photosynthesis with high efficiency. In this review, research progresses in the thermodynamic energy balance, entropy and exergy analysis of photosynthesis processes are discussed with particular focus on the models and development of exergy efficiency calculations. At the same time, the latest progress of artificial photosynthesis is also discussed in this paper.
-
Key words:
- Photosynthesis
- / Energy balance
- / Entropy balance
- / Exergy efficiency
-
光合作用直接或间接为包括人类在内的几乎所有生物的生存提供了物质和能量来源。在环境污染和能源危机愈演愈烈的今天,光合作用产生的生物质将成为重要的新能源。对人工光合作用的研究目前是新能源领域中的热点[1~4]。虽然人类早在18世纪就开始了对光合作用的定量研究[5],到20世纪40年代明确了二氧化碳中的碳素转化途径,但由于生物过程的复杂性,对其微观作用机理尚未研究透彻,即使对光合作用的热力学效率,仍充满争议。Petela[6]认为光合过程有效能效率很低,仅为2.61%;Lems[7]的研究则认为光合过程效率高达31%~34%。因此,确定光合过程的热力学效率是研究人工光合作用、开发新型可再生能源的关键一步。
在以化石燃料为主的“旧能源”时代,热力学的研究相当成熟,在热力学理论指导下热机效率不断提高[8],一些超超临界机组有效能效率达到45.9%[9]。但是在以利用太阳能为代表的新能源领域,热力学的研究还相当薄弱。原因主要有两个方面:(1) 光合作用的体系是一个特殊而复杂的体系,即存在物质进出也有能量交换,不是通常蒸汽动力循环研究的封闭、不做非体积功的体系[10];(2) 光合作用的一些反应发生在叶绿体中尺寸非常小(直径0.25~0.8 μm)的内囊体中。此时熵、焓、自由能等热力学量如何随维度、尺寸、形貌的变化尚不明确[11]。尽管如此,本文试图总结目前光合作用的热力学研究,以指导提高人工光合过程的效率。
1 光合作用机理
绿色植物产氧光合作用的总反应可用式(1) 来表示:
光合作用的具体过程为:(1) 原初反应:光合色素吸收、传递光能给反应中心的叶绿素分子,后者发生电荷分离,失去电子产生空穴。为填补空穴从水中夺取电子,造成水的分解。
(2) 同化力形成:在高等植物中,电荷分离后,在光合系统Ⅰ和光合系统Ⅱ参与下,并且将二者串联起来的电子线式传递,导致三磷酸腺苷(ATP)和还原型烟酰胺腺嘌呤二核苷酸磷酸(NADPH)的生成。
其中,NADP+为氧化型烟酰胺腺嘌呤二核苷酸磷酸,ADP为腺苷二磷酸。(3) 碳同化:利用上一阶段产生的同化力(ATP和NADPH),将CO2转化成淀粉和葡萄糖等光合作用产物。
2 光合作用热力学分析
2.1 能量衡算分析
能量衡算法是建立在热力学第一定律基础之上的热力学分析方法,通过物料和能量的衡算分析能量的转化、利用及损失情况,确定进、出能量的数量,求出能量利用率。
许大全专著《光合作用效率》[12]中用IRGA测定了不同植物光合量子效率(每吸收一个光量子所固定的CO2分子数或释放的O2分子数),其值大约在2%~4%,同时还分析了外部环境因素和植物自身因素对光量子效率的影响。这是基于不同环境条件(温度、二氧化碳浓度、光照强度)下,对植物光合作用效率的实验值。下面主要介绍不同研究基于其假定的条件,计算得到的理论能量效率。
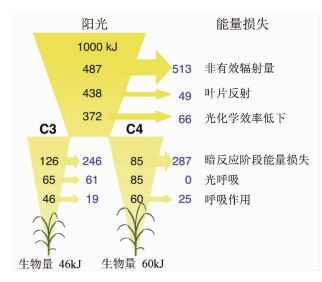
Zhu等[13]在研究光合作用的最大光能量转化效率中,将造成能量损失的原因分为:(1) 对非有效辐射(400~740 nm以外波长的太阳光)的低效吸收,导致占入射太阳光总能量51.3%的能量不能被利用;(2) 叶片反射部分光合有效辐射,大约有10%的有效辐射量被反射,这部分能量占入射光总能量的4.9%;(3) 叶绿素分子被光子激发后,前者电子的快速释放导致部分能量未被利用,这部分损失的能量为6.6%;(4) 暗反应阶段的能量损失,C3植物和C4植物在该阶段反应路径不同,其中C3植物的能量损失为24.6%,C4植物则为28.7%;(5) 光呼吸导致的能量损失,C4植物不进行光呼吸,C3植物光呼吸的能量损失为6.1%;(6) 呼吸作用导致的能量损失,C3植物为1.9%、C4植物为2.5%。以上数据均是在30℃、二氧化碳浓度0.38‰、其他条件理想的情况下的最大效率和最小能量损失(见图 1)。可以看出,C3植物的总能量效率为4.6%,而C4植物为6%。
图 1
光合作用理想状态下能量分布
Figure 1.
Energy distribution of photosynthesis under ideal state
该研究是目前关于光合作用能量效率最具代表性的研究之一,其结论对于提高植物能量效率具有重要指导意义。通过该研究可以发现,对于C3植物如果消除其光呼吸,理论能量效率可以达到10.7%。得益于基因工程技术的进步,通过改造C3植物的基因,削弱甚至消除C3植物的光呼吸,成为提高植物光合作用效率的途径。2002年,Kebeish等[14]通过基因工程将大肠杆菌表达乙醇酸代谢途径的基因导入到拟南芥中,得到采用大肠杆菌乙醇酸代谢途径的拟南芥。经过基因改造的拟南芥其叶绿体中的乙醇酸不经过光呼吸过程直接变为甘油酸用以合成糖类,从而提高了光合作用效率。提高C3植物细胞内的CO2浓度可以抑制二磷酸核酮糖羧化酶(Rubisco)的氧化活性,从而抑制C3植物的光呼吸。Lieman-Hurwitz等[15]将蓝藻几个控制HCO3-积累的基因导入到拟南芥中,提高了转基因植物的光合作用效率。除了将少数调控HCO3-的基因导入植物外,Price等[16]进行了一项更加宏大的工作,通过基因工程在C3植物叶绿体中建立一个完全的蓝藻的CO2浓缩机制。该研究还显示叶片反射也是造成光合作用效率降低的重要因素。
植物通过非光合化学猝灭(NPQ)将部分光能以热能的形式散失掉。NPQ受类囊体膜质子梯度诱导了类囊体膜构型变化的调节。但是,目前鲜有通过削弱植物NPQ来提高植物光合作用效率的研究。削弱植物叶片反射将是提高植物光合作用能量效率的一个重要研究方向。Kiss等[17]发现质子梯度的分子探头Psbs蛋白能够调节与NPQ有关的类囊体膜构型。该研究准确地分析了光合作用能量损失的原因,对提高光合作用效率的研究有一定的指导意义。但是,能量不仅有大小之分,还有品位的区别,能量衡算法不能反映能量品味高低变化;并且,该研究给出的植物光合作用最大能量转化效率仅仅是在一定条件下的最大效率,并不是热力学上能量转化的限度。
2.2 熵分析
2005年,Jennings等[18]计算了太阳光辐射过程的熵变和环境熵变。在这项研究中,他们通过计算得到在光强较弱的条件下光合作用过程的熵变小于零。这项研究在当时引起不小的争议。Knox等[19]仔细分析了他们的工作后提出,由于Jennings等忽略了光合色素受激发过程的熵变,才导致了光合作用熵变小于零的错误结论。并且他们认为Jennings的研究结果一定程度上是受“瞬时、微小范围系统不严格遵守热力学第二定律”[20, 21]观点的影响。在一段时间里该领域发表有多篇文献,科学家围绕这个问题展开了激烈论战[22~25]。两位学者争论的焦点在于光合色素受激发过程的熵变是否可以忽略。Jennings认为这个过程的熵变为可以忽略。
目前普遍认为,叶绿素吸收光子之后马上由基态变为激发态,光子不被存储。只要明确这个光吸收过程中的光量子吸收量和产物的自由能,就可以得到这个过程的熵变。得益于光声技术的进步,光量子能量已可以被准确测定[26]。通过计算可以知道,在光合作用的原初反应阶段,效率接近100%,光合色素受激发的熵变很小。因此,Jennings认为光合色素受激发过程的熵变可以忽略是有一定根据的。这里有两个过程很容易混淆,植物吸收光的过程和色素受光量子激发的过程。并不是所有照射到叶片表面的光量子都能被吸收用来激发色素,这两个研究都没有考虑光吸收过程中的熵变。一些研究认为植物对太阳光的低效吸收是光合作用过程中有效能损失的主要来源(在2.3中详细讨论),因此,也应该是熵增的重要来源,然而这在之前两个研究中都被忽略了。
Albarran-Zavala等[27]通过熵分析得到高等植物光合作用过程中总熵变的计算式(式(5)),计算得到高等植物光合作用过程中总熵变ΔSuniverse=37.543kJ/(mol·K)。有意思的是,在该项研究中熵增主要来自植物将未利用的能量散失到环境中,这部分的熵增大约为25.742kJ/(mol·K)。在一些有效能的研究中,认为植物的蒸腾作用是最大的有效能损失的来源,水蒸发带走了大量有效能[6]。因此有理由相信,控制蒸腾作用或者采用不含蒸腾作用的过程来利用太阳能将极大地提高太阳能利用效率。
2.3 有效能分析
有效能分析与熵分析法类似也是以热力学第一、第二定律为基础,可以确定过程的有效能损失和有效能效率,更全面地反映有效能损失的部位和数量。化学有效能的标准解释如式(6) 所示[28],其中B代表有效能,H为焓,S为熵,T0为环境温度。
早期对光合作用的研究使用吉布斯自由能来计算过程中的有效能变化[29~31],如式(7),其中G为吉布斯自由能,T为系统的温度。因为生物化学反应发生在接近周围环境(或者基态)的压力和温度条件下,因此吉布斯自由能与有效能基本上是相等的。
一直以来,光合作用的有效能效率是将反应(1) 的吉布斯自由能变化除以实验测得的光量子有效能得到的[29],如式(8) 所示,其建立的模型大致如图 2所示。其中,ηps为光合作用的有效能效率,△GR1是反应(1) 的吉布斯自由能变化,∑Bphoton为驱动反应(1) 的光量子有效能的和。应该注意的是,∑Bphoton为光量子的能量值。
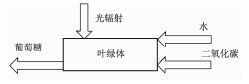 图 2
早期光合作用有效能研究模型
Figure 2.
The model of photosynthesis exergy in early research
图 2
早期光合作用有效能研究模型
Figure 2.
The model of photosynthesis exergy in early research
之后的一些研究[27]将光合作用分为光反应(如反应(9))和暗反应(如反应(10))。其中,Pi为磷酸基团。光反应的有效能效率ηLR用等式(8) 计算,其中用反应(9) 的吉布斯自由能变换△GR9代替△GR1。暗反应有效能效率如式(11) 所示,等于产生葡萄糖的反应(1) 的吉布斯自由能变化比上产生ATP和NADPH反应的吉布斯自由能之和。因此,光合作用总的有效能效率ηps为光反应效率ηLR和暗反应效率ηDR的乘积,如式(12) 所示。式(12) 的计算和式(8) 计算是等价的。
1977年,Chain等[32]发现植物对不同波长的光的吸收率是不同的。400~700 nm为有效光合辐射(PAR)。通过普朗克辐射定律(等式(13))可以计算得有效辐射所含的有效能大约占到达地球表面总辐射有效能的43%。建立如图 3所示的光合作用模型,则修正后的光合作用有效能效率为式(14),ηps=13%[33]。应该注意到该处通过普朗克辐射定律计算得到的是有效光合辐射的能量密度,而不是有效能密度,一般认为用光量子能量代替有效能产生的误差不大。通过比较图 2和图 3的模型不难发现,如果能够采取措施使得植物对400~700 nm以外的非有效光合辐射区具有较好的吸收能力,光合作用的有效能效率将大幅度提高,甚至达到30%以上。
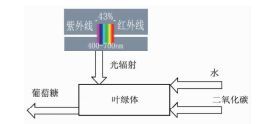 图 3
考虑有效光辐射的光合作用模型
Figure 3.
The model of photosynthesis exergy in view of photosynthetic active radiation(PAR)
图 3
考虑有效光辐射的光合作用模型
Figure 3.
The model of photosynthesis exergy in view of photosynthetic active radiation(PAR)
1996年,Miyashita等[34]发现作为蓝藻主要光合色素的叶绿素d其最大光吸收波长达到710nm;而Chen等[35]发现的叶绿素f更是将光合作用的光吸收范围拓展到722nm,随后他们发现其吸收范围可以进一步拓展到750nm[36]。可以吸收750 nm的光合色素的发现,为拓展光合作用吸收光谱的研究提供了重要材料。通过基因工程将叶绿素f的相关基因导入高等植物,使得高等植物拓展其光谱利用范围是提高光合作用有效能效率的不错思路。但是,由于叶绿素f不能独立完成光化学反应,如何将其吸收的远红光高效地传递给主要吸收红光的叶绿素a,进而推动以叶绿素a为反应中心的光化学作用的进行是该类研究不得不解决的问题。
人工光合作用是解决全光谱吸收问题的另一个重要方向。2016年,Wang等[37]将植物光合系统Ⅱ和硅基光化学电池耦合设计了自然-人工光合杂化系统。该系统通过独特的串联设计实现了人工系统对短波段、自然系统对可见光段太阳能的吸收。这种通过串联结构互补吸收太阳光谱的方法,为设计人工光合作用系统充分利用太阳能不同波段来提高光合作用效率提供了新策略。
近些年对于光合作用的有效能分析有两项深入研究。第一项是Petela[6]在2008年提出的,他将植物的叶片视为“黑箱”,只考虑物质携带有效能进出和光量子能量输入叶片,不考虑叶片内部复杂的生理机制。由此光合作用的有效能效率为产物的有效能与输入的反应物携带的有效能以及光量子的能量和的比值,如式(15) 所示。其中,ηb为有效能效率,nsu为葡萄糖的生产速率,bsu为葡萄糖的摩尔有效能,nw为液态水的输入速率,bw为液态水的摩尔有效能,nco2为二氧化碳的输入速率,bco2为二氧化碳的摩尔有效能。建立合理模型、基于可信的实验数据准确计算物质输入输出的量和携带的单位有效能是该文的创新点之一。在处理输入的光辐射能量的时候,该研究认为辐射削弱系数γ=0.7,对有效光合辐射的吸收率αv=0.88,对非有效光合辐射的吸收率为0.55,然而,在以往的研究中通常认为植物对有效光合辐射完全吸收,对非有效光合辐射完全不吸收;bs为太阳辐射的有效能,bv为有效光合辐射的有效能,这两个数据是基于实验数据建立近似模型得到的。
据此计算得到光合作用的有效能效率为2.61%,最大的有效能损失来自水分的蒸腾作用,高达93.1%。图 4为考虑了蒸腾作用带走有效能的热力学模型。蒸腾作用是陆生植物吸收和运输水分的主要推动力。假如没有蒸腾作用,植物无法将水分运达远离根系的植株顶端,同时矿物盐在植物中的运输也无从谈起。
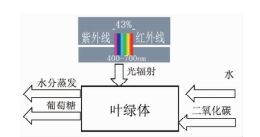 图 4
考虑有效光辐射及蒸腾作用的光合作用模型
Figure 4.
The model of photosynthesis exergy in view of photosynthetic active radiation(PAR)and transpiration
图 4
考虑有效光辐射及蒸腾作用的光合作用模型
Figure 4.
The model of photosynthesis exergy in view of photosynthetic active radiation(PAR)and transpiration
第二项深入研究是Silva等[38]开展的,与Petela[6]的研究方法完全不同,他们关注植物复杂的生理机制,建立了迄今为止最为详尽的植物光合作用模型。模型采用Lems等提供的方法计算化学有效能[39],其有效能流动情况见图 5。他们把光合作用的步骤细分为:光合有效辐射吸收、光合非有效辐射吸收、电子在光合系统Ⅱ和光合系统Ⅰ中的电子传递、ATP合成、卡尔文循环,在每一步都讨论研究物质和有效能流动;同时,他们还研究了影响植物总体的因素:光反射、蒸腾作用、光呼吸、呼吸作用。
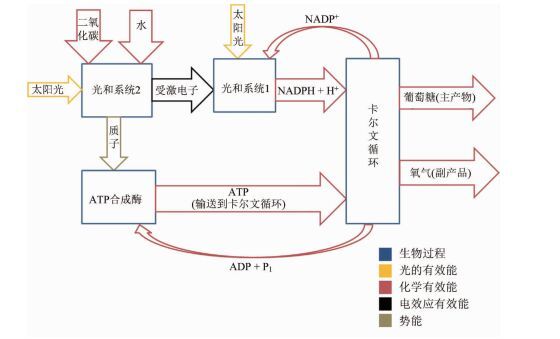 图 5
光合作用过程中有效能流动
Figure 5.
Energy flow in photosynthesis
图 5
光合作用过程中有效能流动
Figure 5.
Energy flow in photosynthesis
该研究基于修正的普朗克公式来计算光子的摩尔有效能,如式(16)。与之前的研究类似,这里也是用光子能量来代替有效能。长期以来用光量子的能量值来代替有效能来进行计算,说明目前对光量子有效能的理解还很缺乏。
在光合作用中,色素失去的电子将在一系列中间载体的驱动下远离色素。分析这些中间载体的工作已经远远超出目前热力学的研究范畴,有兴趣的读者可以参考许大全在2013年发表的专著《光合作用学》[40]。该研究基于标准吉布斯自由能的公式(式(17)),得到描述电子在载体间传递有效能变化的公式(式(18))。其中△Belec为电子在载体间有效能变化,△ε0为标准还原电位[41],[A]i为载体i的活度,vi为载体i的化学计量数。值得注意的是,一个过程的自由能变化,只有当这个过程达到平衡了才能确定。因为需要获取该状态存在的过程中电子转移的平衡关系式,所以通过瞬时激发态的还原电位来计算吉布斯自由能变化是不准确的。
该研究认为电子传递链传递的有效能可合成18mol ATP。光反应向暗反应传递18mol ATP、12mol NADPH,其中含有3509191J的有效能,卡尔文循环中有6611139J的有效能被损耗。则卡尔文循环的有效能效率为81.2%。
综合以上分析,再将叶片反射光、蒸腾作用、光呼吸作用、光合作用消耗的有效能考虑在内,得到的植物光合作用有效能效率为3.91%,最大的有效能损失来自叶片对太阳光的低效吸收。
与光合过程效率低下的陆生植物相比,藻类和光合细菌生活在液相环境中,无需蒸腾作用也能获得足够的水分,因此蒸腾过程的有效能损失可以避免,所以,对于藻类的光合过程,其理论有效能效率可望达到13%,也有研究认为微藻的最大效率为9%[42],藻类的理论光合过程有效能效率可以达到13%。目前对人工光合作用的研究,重点关注的就是利用微藻和细菌生产有机物。之前所述提高光合作用效率的方法大多与基因工程有关。虽然基因工程是改善能源作物生产性能、提高光合作用效率的有力武器,但是种植、推广转基因作物对生态环境、人类健康的危害尚不明确。不能把提高光合作用效率的全部希望寄托在基因工程上。
近年来人工光合作用的研究取得令人瞩目的成就。Sakimoto等改造在厌氧状态下可以产生乙酸的细菌Moorella thermoacetica[43],使之能够在表面析出硫化镉纳米粒子。改造过的细菌吸收光生产乙酸的效率达到(2.44 ± 0.62)%,这个数值明显超过自然环境中陆生植物1%左右的效率,接近生物反应器条件下微藻的效率(3.8%)[44]。Liu等[45]开发了一种基于新型钴-磷(Co-P)水分解催化剂和细菌Ralstonia eutropha的高效率人工光合作用系统。催化剂利用太阳能产生的电能将水分解成氢气和氧气,细菌利用产生的氢气和吸收的CO2产生有机物,在CO2浓度100%条件下,每消耗1kWh的电能可固定4.1mol的CO2,能量转化效率为(54±4)%,将现有光伏发电效率(18%)考虑进去,该系统整体效率为9.7%左右;而在空气条件下(CO2浓度0.4‰),每消耗1kWh电能固定1.5mol的CO2,能量转化效率(20±3)%,整体效率为3.6%。Liu等的研究无疑是振奋人心的,有力地证明了更高的太阳能转化效率是可以实现的。与光伏发电相比人工光合作用的优势在于它可以很好地克服太阳能发电和供电不稳定的问题,吸收的太阳能可以转化成稳定的化学能储存起来。该研究的突出创新是高效水分解催化剂的研制,而性能更优良催化剂的出现势必助推人工光合作用的研究。Yang等[46]通过等离子辅助原子层沉积技术在p+n-Si晶片上沉积Co3O4/Co(OH)2两相薄膜得到了目前报道的最高的光电催化氧化HO的性能,并且催化剂维持活性长达70h。与目前许多关于光电催化水分解催化剂的研究相比,该研究不仅在理想的电化学测试条件下检验了催化剂活性,并且把催化剂负载到光电极上测试真实条件下的催化性能,充分证明其在人工光合作用研究领域的实际应用价值。
除了开发新型催化剂研究光电催化水分解外,基于植物光系统Ⅱ水裂解催化中心的原理进行仿生研究,构建人工水裂解催化中心也是一个重要的研究方向。光系统Ⅱ晶体结构的揭示[47]为人工模拟水裂解催化中心提供了蓝图。2015年,Zhang等[1]合成了一种Mn4Ca簇合物,其结构接近于植物水裂解催化中心。该模拟物中Mn离子在氧化-还原性质、电子顺磁特性及化学反应性质方面与生物水裂解催化剂类似,而且同样具有催化水裂解的功能。该研究不仅对于构建人工光合作用系统利用太阳能具有重要的科学意义和应用价值,对研究自然界光系统Ⅱ水裂解中心的结构和水裂解机理也有重要的参考价值。
3 结语
利用热力学的理论和方法对光合作用各个过程进行热力学分析,不仅可以明确能量损耗的数量,更重要的是了解能量损耗的部位和原因。通过以上分析可知,如果植物能进行全光谱吸收并且不进行蒸腾作用,其理论光合作用效率可以达到32%~37%。考虑到植物只能高效吸收可见光(400~700 nm),理论光合作用效率降低到13%。陆生植物为了维持正常的生理机能而必须进行蒸腾作用,将理论光合作用效率进一步降低到2.61%~3.91%。由此可知,生活在液相中不进行蒸腾作用的微藻和细菌其理论光合作用效率可以达到13%,是进行人工光合作用研究的理想材料。目前对于人工光合作用的研究主要关注改造细菌,让其成为人工光合作用的反应器。微藻作为一种比细菌更高级的生命形式,其进化更加成熟、生理机制更加完善,其作为人工光合作用反应器的潜力巨大。未来,开展对微藻的深入研究,是实现人工光合作用的重要途径。
-
-
[1]
C Zhang, C Chen, H Dong et al. Science, 2015, 348:690~693. doi: 10.1126/science.aaa6550
-
[2]
X Li, J Wen, J Low et al. Sci. China Mater., 2014, 57(1):1~31. doi: 10.1007/s40843-014-0007-1
-
[3]
M R Singh, A T Bell. Energy Environ. Sci., 2015, 9(1):193~199.
-
[4]
李晓慧, 范同祥. 化学进展, 2011, (9):1841~1853.
-
[5]
柯世省. 生物学通报, 2003, 38(8):59~60.
-
[6]
R Petela. Solar Energy, 2008, 82(4):311~328. doi: 10.1016/j.solener.2007.09.002
-
[7]
S Lems. Int. J. Exergy, 2010, 7(3):333~351. doi: 10.1504/IJEX.2010.031988
-
[8]
姚明宇, 张广才, 聂剑平. 热力发电. 2012, 41(8):1~5, 23.
-
[9]
刘翔, 方曹明, 范浩杰等. 锅炉技术. 2016, 47(3):1~5, 30.
-
[10]
D Mauzerall. Photosynth. Res., 2013, 116(2):363~366.
-
[11]
W Qi. Acc. Chem. Res., 2016, 49(9):1587~1595. doi: 10.1021/acs.accounts.6b00205
-
[12]
许大全. 光合作用效率. 上海:上海科学技术出版社, 2002.
-
[13]
X G Zhu, S P Long, D R Ort. Curr. Opin. Biotech., 2008, 19(2):153~159. doi: 10.1016/j.copbio.2008.02.004
-
[14]
R Kebeish, M Niessen, K Thiruveedhi et al. Nature Biotechnol., 2007, 25(5):593~599. doi: 10.1038/nbt1299
-
[15]
J Lieman-Hurwitz, S Rachmilevitch, R Mittler et al. Plant Biotech. J., 2003, 1(1):43~50. doi: 10.1046/j.1467-7652.2003.00003.x
-
[16]
G D Price, M R Badger, S V Caemmerer. Plant Physiol., 2010, 155(1):20~26.
-
[17]
A Z Kiss, A P Ruban. J. Biol. Chem., 2008, 283(7):3972~3978. doi: 10.1074/jbc.M707410200
-
[18]
R C Jennings, E Engelmann, F Garlaschi et al. Biochim. Biophys. Acta, 2005, 1709(3):251~255. doi: 10.1016/j.bbabio.2005.08.004
-
[19]
R S Knox, W W Parson. Biochim. Biophys. Acta, 2007, 1767(10):1189~1193. doi: 10.1016/j.bbabio.2007.07.004
-
[20]
G M Wang, E M Sevick, E Mittag et al. Phys. Rev. Lett., 2002, 89(5):716~722.
-
[21]
C Bustamante, J Liphardt, F Ritort. CR Phys., 2005, 58(7):43~48.
-
[22]
J Lavergne. Biochim. Biophys. Acta, 2006, 1757(11):1453~1459. doi: 10.1016/j.bbabio.2006.05.017
-
[23]
R C Jennings, A P Casazza, E Belgio et al. Biochim. Biophys. Acta-Bioenerg., 2006, 1757(11):1460~1462. doi: 10.1016/j.bbabio.2006.06.012
-
[24]
R S Knox, W W Parson. Biochim. Biophys. Acta-Bioenerg., 2007, 1767(10):1198~1199. doi: 10.1016/j.bbabio.2007.08.002
-
[25]
R C Jennings, E Belgio, A P Casazza et al. Biochim. Biophys. Acta, 2007, 1767(1767):1194~1197.
-
[26]
S P Mielke, N Y Kiang, R E Blankenship et al. Biochim. Biophys. Acta, 2011, 1807(9):1231~1236. doi: 10.1016/j.bbabio.2011.06.007
-
[27]
E Albarrán-Zavala, F Angulobrown. Entropy, 2007, 9(4):152~168. doi: 10.3390/e9040152
-
[28]
H K Joseph. Brit. J. Appl. Phys., 1951, 2(7):183. doi: 10.1088/0508-3443/2/7/302
-
[29]
I Asimov. Photosynthesis. New York:Basic Books, Inc., 1968.
-
[30]
J A Bassham, B B Buchanan. Photosynthesis. 1982, 2:141~189.
-
[31]
A L Lehninger. Bioenergetics:the molecular basis of biological energy transformations. Menlo Par.:WA Benjamin, 1971.
-
[32]
R K Chain, D I Arnon. PNAS, 1977, 74(8):3377~3381. doi: 10.1073/pnas.74.8.3377
-
[33]
J R Bolton, D O Hall. Photochem. Photobiol., 1991, 53(4):545~548. doi: 10.1111/php.1991.53.issue-4
-
[34]
H Miyashita, H Ikemoto, N Kurano et al. Nature, 1996, 383(6599):402. doi: 10.1038/383402a0
-
[35]
M Chen, H Scheer. Science, 2010, 329(5997):1318~1319. doi: 10.1126/science.1191127
-
[36]
M Chen, R E Blankenship. Trends Plant Sci., 2011, 16(8):427~431. doi: 10.1016/j.tplants.2011.03.011
-
[37]
W Wang, H Wang, Q Zhu et al. Angew. Chem., 2016, 128(32):9375~9379. doi: 10.1002/ange.201604091
-
[38]
C S Silva, W D Seider, N Lior. Chem. Eng. Sci., 2015, 130:151~171. doi: 10.1016/j.ces.2015.02.011
-
[39]
S Lems. Int. J. Exergy, 2007, 4(4):339~356. doi: 10.1504/IJEX.2007.015077
-
[40]
许大全. 光合作用学. 北京:科学出版社, 2013.
-
[41]
D G Nicholls, S J Ferguson. Bioenergetics 3. San Diego, Calif.:Academic Press, 2002.
-
[42]
R H Wijffels, M J Barbosa. Science, 2010, 329(5993):796~799. doi: 10.1126/science.1189003
-
[43]
K K Sakimoto, A B Wong, P Yang. Science, 2016, 351(6268):74~77. doi: 10.1126/science.aad3317
-
[44]
E Sorgüven, M Özilgen. Energy, 2013, 58(9):679~687.
-
[45]
C Liu, B C Colón, M Ziesack et al. Science, 2016, 352(6290):1210~1213. doi: 10.1126/science.aaf5039
-
[46]
J Yang, J K Cooper, F M Toma et al. Nat. Mater., 2017,16:335~341.
-
[47]
Y Umena, K Kawakami, J R Shen et al. Nature, 2011, 473(7345):55~60. doi: 10.1038/nature09913
-
[1]
-

图 3 考虑有效光辐射的光合作用模型
Figure 3 The model of photosynthesis exergy in view of photosynthetic active radiation(PAR)

图 4 考虑有效光辐射及蒸腾作用的光合作用模型
Figure 4 The model of photosynthesis exergy in view of photosynthetic active radiation(PAR)and transpiration
-


 下载:
下载:




 扫一扫看文章
扫一扫看文章
计量
- PDF下载量: 0
- 文章访问数: 0
- HTML全文浏览量: 0
 下载:
下载:
