引用本文:
杨科成, 崔凤超, 李云琦. 核糖核酸酶Sa表面水和尿素分子的分布和动力学行为的分子动力学模拟[J]. 应用化学,
2018, 35(10): 1243-1248.
doi:
10.11944/j.issn.1000-0518.2018.10.170342 Citation:
YANG Kecheng, CUI Fengchao, LI Yunqi. Distribution and Dynamics of Water and Urea in Hydration Shell of Ribonuclease Sa: A Molecular Dynamics Simulation Study[J]. Chinese Journal of Applied Chemistry,
2018, 35(10): 1243-1248.
doi:
10.11944/j.issn.1000-0518.2018.10.170342
Key Laboratory of Synthetic Rubber, Changchun Institute of Applied Chemistry, Chinese Academy of Sciences, Changchun 130022, China
b.
University of Chinese Academy of Sciences, Beijing 100049, China
Corresponding author:
CUI Fengchao, research assistant; Tel:0431-85262524, Fax:0431-85262535, E-mail:fccui@ciac.ac.cn Research interests:simulation research of macromolecules and biomolecules; LI Yunqi, professor; Tel/Fax:0431-85262535, E-mail:yunqi@ciac.ac.cn; Research interests:simulation research of macromolecules and biomolecules
Received Date:
18 September 2017 Accepted Date:
24 November 2017 Revised Date:
16 October 2017 Available Online:
10 October 2018
Fund Project:
Supported by the National Natural Science Foundation of China(No.21374117, No.21504092), the One Hundred Person Project of the Chinese Academy of Sciences
Abstract:
Extensive molecular dynamics simulations were performed to study the distribution and dynamics of water and urea in the hydration shell of ribonuclease Sa(RNase Sa) under different urea concentrations. It is found that urea molecules have stronger interactions with protein than water molecules and are enriched on the surface of RNase Sa by displacing water molecules. Urea molecules prefer to interact with hydrophobic residues and form hydrogen bonds with the backbone of RNase Sa. The transitional and rotational dynamics of urea molecules are much slower than those of water molecules. Besides, the increased urea concentrations can slow down the transitional and rotational dynamics of water molecules, but have no regular influences on the dynamics of urea molecules. Our results can help understanding the different influences of urea and water molecules on the stability of proteins.
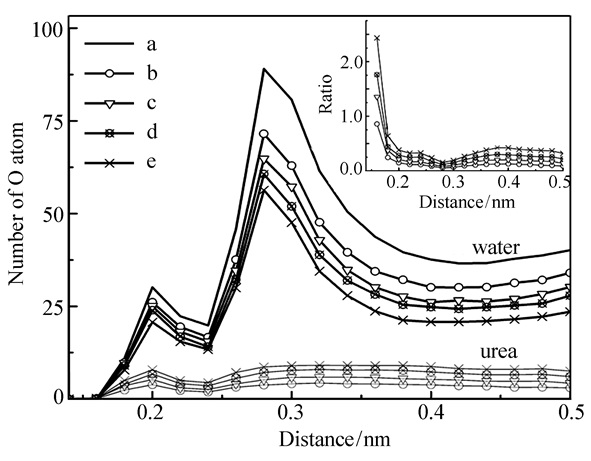
Figure 2.
Distributions of oxygen atoms of water(dark color) and urea(light color) depending on the distance away from any atom of RNase Sa surface. The inset shows the distribution of ratio of oxygen atoms of urea and water
a, b, c, d, e denote the 0, 2, 4, 6, 8 mol/L urea concentrations, respectively
Table 1.
The partition coefficient of urea in the hydration shell and bulk(Kp) and distributions of urea molecules around hydrophobic and polar/charged residues of RNase Sa under different urea concentrations
Kp is the ratio of Ruw(the urea/water ratio) in the hydration shell to Ruw in bulk water. SASA is the solvent accessible surface area of a residue. Nurea is the number of urea molecules. RO* is the ratio of Nurea around hydrophobic residues and polar/charge residues at unit SASA.
RD/A is the ratio of urea and water acting as hydrogen bond(HB) donor and acceptor; RBB/SC is the ratio of HBs of urea and water formed with backbone(BB) and side chain(SC) of RNase Sa; RU/W is the ratio of protein-urea(U) HBs and protein-water(W) HBs normalized with the number of urea and water molecules in the hydration shell.
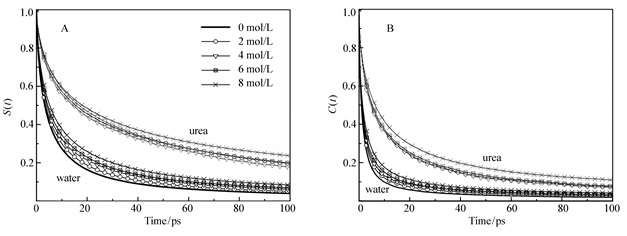
Figure 3.
(A) Survival time correlation function S(t), and (B)rotational time correlation function(TCF) Cμ(t) of water(dark color) and urea(light color) in the hydration shell under different urea concentrations
Table 3.
Amplitude-weighted average survival time and rotational time of water and urea molecules in hydration shell and average interaction strength between them and protein in different urea concentrations
Biedermannová L, Schneider B. Hydration of Proteins and Nucleic Acids:Advances in Experiment and Theory. A Review[J]. Biochim Biophys Acta,
2016, 1860(9):
1821-1835.
doi: 10.1016/j.bbagen.2016.05.036
[2]
Hu J, Cao Z X. Water Science on the Molecular Scale:New Insights into the Characteristics of Water[J]. Natl Sci Rev,
2014, 1(2):
179-181.
doi: 10.1093/nsr/nwt015
[3]
Dill K A, MacCallum J L. The Protein-Folding Problem, 50 Years On[J]. Science,
2012, 338(6110):
1042-1046.
doi: 10.1126/science.1219021
[4]
Walters J, Milam S L, Clark A C. (2009)Practical Approaches to Protein Folding and Assembly: Spectroscopic Strategies in Thermodynamics and Kinetics[M]//Michael L Johnson, Jo M Holt, Gary K Ackers. Methods in Enzymology. Academic Press, 2009, 8: 1-39.
[5]
Bellissent-Funel M C, Hassanali A, Havenith M. Water Determines the Structure and Dynamics of Proteins[J]. Chem Rev,
2016, 116(13):
7673-7697.
doi: 10.1021/acs.chemrev.5b00664
[6]
Doster W, Cusack S, Petry W. Dynamical Transition of Myoglobin Revealed by Inelastic Neutron Scattering[J]. Nature,
1989, 337(6209):
754-756.
doi: 10.1038/337754a0
[7]
Qvist J, Ortega G, Tadeo X. Hydration Dynamics of a HalophilicProtein in Folded and Unfolded States[J]. J Phys Chem B,
2012, 116(10):
3436-3444.
doi: 10.1021/jp3000569
[8]
Canchi D R, García A E. Cosolvent Effects on Protein Stability[J]. Annu Rev Phys Chem,
2013, 64(1):
273-293.
doi: 10.1146/annurev-physchem-040412-110156
[9]
Frank H S, Franks F. Structural Approach to the Solvent Power of Water for Hydrocarbons; Urea as a Structure Breaker[J]. J Chem Phys,
1968, 48(10):
4746-4757.
doi: 10.1063/1.1668057
[10]
Hua L, Zhou R, Thirumalai D. Urea Denaturation by Stronger Dispersion Interactions with Proteins than Water Implies a 2-Stage Unfolding[J]. Proc Natl Acad Sci USA,
2008, 105(44):
16928-16933.
doi: 10.1073/pnas.0808427105
[11]
Candotti M, Esteban-Martín S, Salvatella X. Toward an Atomistic Description of the Urea-Denatured State of Proteins[J]. Proc Natl Acad Sci USA,
2013, 110(15):
5933-5938.
doi: 10.1073/pnas.1216589110
[12]
Canchi D R, García A E. Backbone and Side-Chain Contributions in Protein Denaturation by Urea[J]. Biophys J,
2011, 100(6):
1526-1533.
doi: 10.1016/j.bpj.2011.01.028
[13]
Candotti M, Pérez A, Ferrer-Costa C. Exploring Early Stages of the Chemical Unfolding of Proteins at the Proteome Scale[J]. PLoS Comput Biol,
2013, 9(12):
e1003393.
doi: 10.1371/journal.pcbi.1003393
[14]
Laurents D, Perez-Canadillas J M, Santoro J. Solution Structure and Dynamics of Ribonuclease Sa[J]. Proteins:Struct Funct Bioinf,
2001, 44(3):
200-211.
doi: 10.1002/prot.v44:3
[15]
Phillips J C, Braun R, Wang W. Scalable Molecular Dynamics with Namd[J]. J Comput Chem,
2005, 26(16):
1781-1802.
doi: 10.1002/(ISSN)1096-987X
[16]
Mackerell A D, Feig M, Brooks C L. Extending the Treatment of Backbone Energetics in Protein Force Fields:Limitations of Gas-Phase Quantum Mechanics in Reproducing Protein Conformational Distributions in Molecular Dynamics Simulations[J]. J Comput Chem,
2004, 25(11):
1400-1415.
doi: 10.1002/jcc.v25:11
[17]
MacKerell A D, Bashford D, Bellott M. All-atom Empirical Potential for Molecular Modeling and Dynamics Studies of Proteins[J]. J Phys Chem B,
1998, 102(18):
3586-3616.
doi: 10.1021/jp973084f
[18]
Vanommeslaeghe K, Hatcher E, Acharya C. Charmm General Force Field:A Force Field for Drug-Like Molecules Compatible with the Charmm All-atom Additive Biological Force Fields[J]. J Comput Chem,
2010, 31(4):
671-690.
[19]
Berman H M, Westbrook J, Feng Z. The Protein Data Bank[J]. Nucl Acids Res,
2000, 28(1):
235-242.
doi: 10.1093/nar/28.1.235
[20]
Humphrey W, Dalke A, Schulten K. Vmd:Visual molecular dynamics[J]. J Mol Graph,
1996, 14(1):
33-38.
doi: 10.1016/0263-7855(96)00018-5
Figure 2
Distributions of oxygen atoms of water(dark color) and urea(light color) depending on the distance away from any atom of RNase Sa surface. The inset shows the distribution of ratio of oxygen atoms of urea and water
a, b, c, d, e denote the 0, 2, 4, 6, 8 mol/L urea concentrations, respectively
Figure 3
(A) Survival time correlation function S(t), and (B)rotational time correlation function(TCF) Cμ(t) of water(dark color) and urea(light color) in the hydration shell under different urea concentrations
Table 1.
The partition coefficient of urea in the hydration shell and bulk(Kp) and distributions of urea molecules around hydrophobic and polar/charged residues of RNase Sa under different urea concentrations
c(urea)/(mol·L-1)
Kp
Hydrophobic Residues
Polar/Charge Residues
R*
SASA/nm2
Nurea
SASA/nm2
Nurea
2
2.48±0.50
0.420 5±0.318 0
1.01±0.62
0.822 8±0.535 9
1.48±0.78
1.34
4
1.91±0.33
0.422 1±0.321 8
1.41±0.88
0.805 4±0.532 1
1.97±1.03
1.37
6
1.95±0.26
0.462 4±0.347 8
1.94±1.04
0.843 9±0.562 5
2.62±1.24
1.35
8
1.77±0.14
0.413 0±0.297 6
2.35±1.18
0.866 3±0.578 2
3.45±1.61
1.43
Kp is the ratio of Ruw(the urea/water ratio) in the hydration shell to Ruw in bulk water. SASA is the solvent accessible surface area of a residue. Nurea is the number of urea molecules. RO* is the ratio of Nurea around hydrophobic residues and polar/charge residues at unit SASA.
Table 2.
Descriptions of water-protein and urea-protein hydrogen bonds formed
c(urea)/(mol·L-1)
RD/A
RBB/SC
RU/W
Water
Urea
Water
Urea
0
1.43±0.09
-
1.07±0.07
-
-
2
1.35±0.10
2.76±1.14
1.02±0.08
1.36±0.52
1.56±0.26
4
1.28±0.11
2.62±0.81
1.00±0.10
1.15±0.29
1.52±0.21
6
1.30±0.13
2.69±0.62
1.06±0.10
1.35±0.35
1.42±0.19
8
1.30±0.41
2.36±0.40
1.02±0.12
1.20±0.23
1.47±0.18
RD/A is the ratio of urea and water acting as hydrogen bond(HB) donor and acceptor; RBB/SC is the ratio of HBs of urea and water formed with backbone(BB) and side chain(SC) of RNase Sa; RU/W is the ratio of protein-urea(U) HBs and protein-water(W) HBs normalized with the number of urea and water molecules in the hydration shell.
Table 3.
Amplitude-weighted average survival time and rotational time of water and urea molecules in hydration shell and average interaction strength between them and protein in different urea concentrations

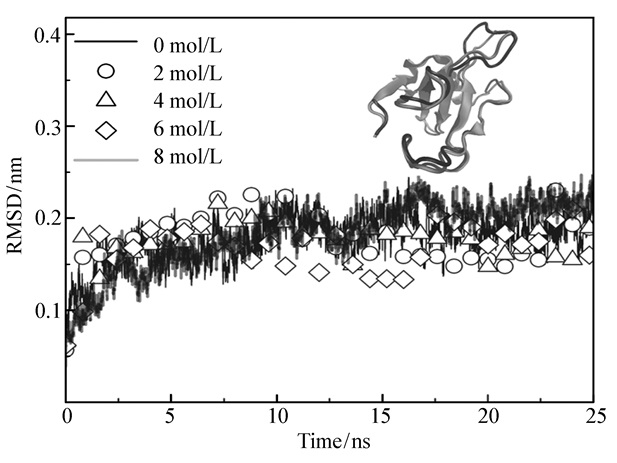
 下载:
下载:

 下载:
下载:




 下载:
下载:
