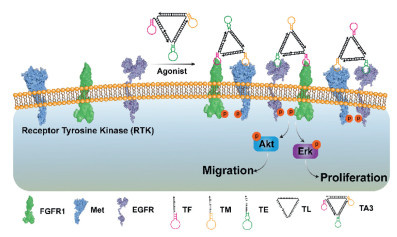
Figure 1.
Schematic representation of a DNA Nano-windmill induced FGFR1, Met and EGFR activation to regulate downstream signaling pathways and cellular behavior.
A transformable DNA Nano-windmill for the activation and regulation of the heterodimerization receptors across RTK families
Kun Liu , Junjun Lu , Jincheng Lin , Yulin Cong , Xiongfeng Luo , Zihong Huang , Tianyi Zhang , Miao Wang , Zhiyong Xie , Hao Li

Receptor tyrosine kinase (RTKs) is closely related to cell motility, growth, differentiation as well as survival. RTKs are a class of important transmembrane signaling proteins that play a key role in physiological and pathological conditions by regulating processes such as cell proliferation, differentiation, survival, and metabolism [1-3]. Generally, RTKs receptor is activated by specific ligands (growth factors, cytokines, hormones, etc.), which promotes homodimerization and then autophosphorylation [3-5]. Epidermal growth factor receptor (EGFR) are widely expressed in epithelial cells of the epidermis, respiratory tract, and gastrointestinal tract, where they regulate barrier function and tissue repair [6]. Fibroblast growth factor receptor (FGFR) are highly expressed in osteoblasts, chondrocytes, and mesenchymal stem cells, they regulate bone development [7]. Hepatocyte growth factor receptor (Met) is widely expressed in hepatocytes, fibroblasts, and mesenchymal stem cells, where it mediates liver regeneration and metabolic regulation hepatic growth factor (HGF)/Met is a key signaling pathway for liver regeneration) and regulates cell migration and tissue remodeling [8]. The challenges of current RTK receptor activation methods mainly include ligand stability and difficulty in artificial ligand design. Native ligands (such as HGF and epidermal growth factor (EGF)) are prone to proteolytic degradation, requiring frequent replenishment in vitro experiments. Developing high-specificity synthetic ligands (such as antibodies or small-molecule agonists) requires precise matching of the receptor conformation, which is technically demanding. RTK interactions are very complicated, which can interact independently of ligands and heterodimerize between RTKs from different subfamilies. Therefore, exploring the heterogeneous interactions of RTKs contributes to a better understanding of fundamental RTK signaling processes in health and disease.
Heterodimerization is a common phenomenon among RTK receptors, and the activity of human epidermal growth factor receptor 2 (ErbB2) appears to primarily depend on EGFR−ErbB2 heterodimers through EGF [9]. Indeed, heterodimerization between RTK receptors lead to phosphorylation of different RTK receptors, which can alter their signal strength as well as duration and mediate different biological responses [10,11]. For example, in myeloid cells, activation of the EGFR−ErbB2 and EGFR−ErbB3 heterodimeric receptor can prolong the signal compared with EGFR homodimers [12]. In hematopoietic cells, ErbB3−ErbB2 heterodimer receptors are able to control the formation and spreading of PI3 kinase-dependent lamellipodia [13]. Therefore, the development of RTK heterodimerization agonists has important implications in biomedical fields. Currently, our group has developed a DNA agonist for the activation of heterodimeric RTK receptors based on the bivalent aptamers [38]. Moreover, many growth factors can activate three receptors at the same time through heterodimerization, such as FGF23 promotes the heterodimerization of FGFR1c and FGFR3c or FGFR1c and FGFR4 while neu differentiation factor (NDF) promotes ErbB2−ErbB3 and ErbB2−ErbB4 heterodimers [14,15]. Developing a novel agonist which activate three receptors by heterodimeric RTK receptors is still unexplored.
Deoxyribonucleic acids (DNAs) have been widely used in various nano-devices to perform unique tasks such as detection, sensing, regulation and computation due to its unique Watson-Crick base pairing rules, sequence controllability, predictable structure and self-assembly programmability [16-28]. Among these, aptamers can specifically bind small molecules or proteins and play an important role in DNA nanodevices [29-35]. Based on aptamers that specifically bind the receptor, we and others have been constructed several novel DNA agonists to activate RTK receptors by homodimerization or heterodimerization [36-39]. We propose to utilize aptamers as building blocks to construct a novel controllable Nano-windmill for the activation of 3 different families RTK receptors by heterodimerization.
Herein, we developed a universal method to construct novel DNA Nano-windmill for the activation of 3 different RTK receptor families by heterodimerization. FGFR1, Met and EGFR of 3 different RTKs respond to fibroblast growth factor (FGF), HGF and EGF respectively, and play important role in the activation of protein kinase B (Akt) and extracellular regulated protein kinases (Erk) signaling pathways, which are closely related to cell migration and proliferation [36,37,40,41]. Our work reported a DNA agonist (DNA Nano-windmill) achieved the simultaneously activation of FGFR1, Met and EGFR signaling by the heterodimerization (Fig. 1). Furthermore, the activation of specific receptors by DNA Nano-windmill was selectively inhibited by complementary strands or small molecules. We believe that the DNA Nano-windmill will have broad applicability in cell therapy and regenerative medicine.
The DNA Nano-windmill consists of an oligonucleotide backbone (TL) as the scaffold and three different aptamers hooking on the extracellular region of FGFR1, Met and EGFR as "capture toes", respectively (Fig. 1) [36,37,39]. The structure diagram of the DNA Nano-windmill was showed in Fig. S1 (Supporting information). The DNA Nano-windmill was named as TA3. The novel DNA Nano-windmill can regulate the activation of FGFR1, Met and EGFR by heterodimerization, following by the activation of the Akt and Erk signal pathway, thereby inducing the cell migration and proliferation.
As a proof of concept, we first dynamically assembled the precursors of DNA Nano-windmill (TFM-F and TFE-T) on the cell membrane (Fig. 2a, Figs. S2 and S3 in Supporting information). The strands TM-F and TE-T with receptor binding activity were both blocked by a complementary strand M-B1 and E-B2 to prevent hybridization with DNA Nano-windmill, called TMR and TER, respectively. Initially, DNA Nano-windmill is anchored to the FGFR1 receptor (TFR) on the cell membrane by TF, and TMR and TER bind to the Met and EGFR receptors, respectively. After adding initiate strand TLM-S or TLE-S, the TMR and TER could interaction with TFR through toehold-mediated strand displacement reaction, which could construct the agonist TFM-F or TFE-T and activate the FGFR1-Met or FGFR1-EGFR heterodimerization. As shown in a fluorescence analysis, with the formation of TFM-F or TFE-T, the quenched FAM and TAMRA fluorescence significantly recovered (Fig. 2b). These results indicated that the agonist TFM-F or TFE-T could successfully assembling and hooking on the FGFR1-Met or FGFR1-EGFR.
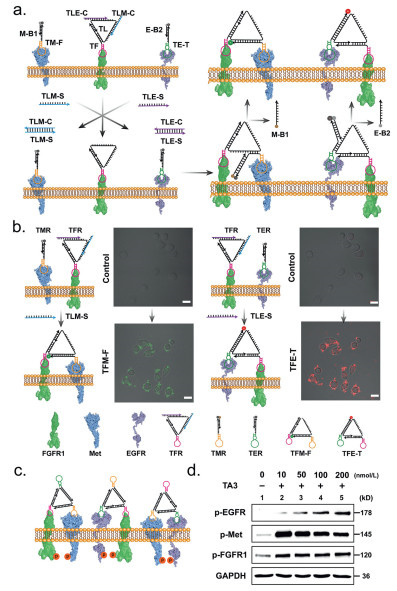
Next, we assembled DNA Nano-windmill (TA3) in vitro using TF, TM, TE and TL for regulating FGFR1, Met and EGFR signaling. Gel analysis indicated that the desired DNA agonist was successfully assembled (Fig. S4a in Supporting information). The strand TLTB tagged with TAMRA (5′) and BHQ2 (3′) was designed to form TA3-F instead of TL (Fig. S4b in Supporting information). The fluorescence of TA3-F was almost eliminated compared with unassembled TA3-F (Figs. S4b and c in Supporting information), suggesting that TA3 forms the expected Nano-windmill structure. After anchoring FGFR1, Met, and EGFR, the agonist TA3 would induce proximity of receptor for cross-phosphorylation. The Met signaling was activated through TA3 within 10 min (Fig. S5 in Supporting information). Indeed, TA3 increased the FGFR1, Met and EGFR phosphorylation levels of A549 cells, with an effective dose as low as 10 nmol/L (Figs. 2c and d). Together, we have developed a novel Nano-windmill that simultaneously activates FGFR1, Met and EGFR signaling.
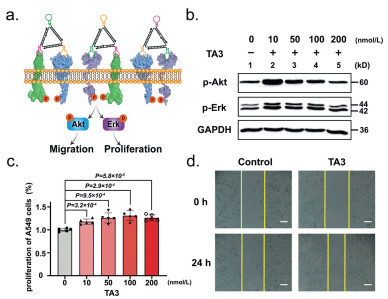
Next, we evaluated the effects of multi-receptor heterodimeric activation on downstream key regulators Erk and Akt in cell motility and proliferation (Fig. 3a). A significant increase in p-Erk and p-Akt expression was observed in A549 cells treated with TA3 in 10 min (Fig. S6 in Supporting information). Compared with 10 nmol/L TA3 treatment, the p-Erk and p-Akt expression remarkably decreased after increasing TA3 concentration to 200 nmol/L in A549 cells (Fig. 3b), probably due to the high concentration of TA3 binding to a single receptor reducing the probability of heterodimerization. Finally, we confirmed that the DNA Nano-windmill (TA3) could enhance motility and proliferation of A549 cells. TA3-promoted cell proliferation increased 1.2X than untreated cells in 24 h (Fig. 3c). The cell proliferation curve of A549 cells treated with TA3 (100 nmol/L) for 24 h was shown in Fig. S7 (Supporting information). Using the scratch wound assay to mimic wound healing in vitro, we investigated the effect of TA3 activation on wound-closure events. In the presence of TA3, the cell motility rate of A549 cells was obviously increased 2.9X compared with untreated group (Fig. 3d and Fig. S8 in Supporting information). The scratch widths of A549 cells treated with TA3 (100 nmol/L) for 24 h at different times were shown in Figs. S9 and S10 (Supporting information). As shown in Fig. S11 (Supporting information), occludin, zonula occludens protein-1 (ZO-1) and claudin-1 protein level enhancement was found in TA3 treated cells compared to the presence of lipopolysaccharides (LPS) respectively. Therefore, these findings demonstrate that DNA Nano-windmill (TA3) could induce the phosphorylation of both Akt and Erk through heterodimerization, enhancing cell motility and proliferation.
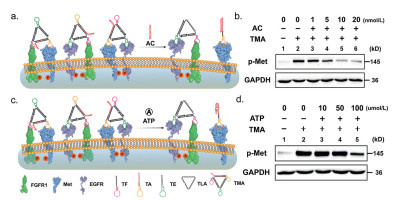
To evaluate the effect of FGFR1, Met and EGFR aptamer deletion of TA3 on signal activation, we designed three sequence TFC, TMC and TEC that does not contain aptamer to replace TF, TM and TE to construct Nano-kite (T1, T2 and T3) (Fig. S12 in Supporting information). The natural polyacrylamide gel electrophoresis showed that TF, TM and TE could be replaced by TFC, TMC and TEC to form Nano-kite (T1, T2 and T3) (Fig. S13 in Supporting information). Next, we evaluated the FGFR1, Met and EGFR phosphorylation levels of A549 cells incubated with T1, T2 and T3 (Fig. S14 in Supporting information). The phosphorylation of EGFR and FGFR1 showed significant crosstalk, while the phosphorylation of Met was independent. Above all, we designed a new Nano-windmill (TMA) to regulate Met signaling. The transformable DNA Nano-windmill (TMA) was designed a specific sequence TA capable of binding complementary chains or small molecules to replace TM strand (Fig. S15 in Supporting information). The strands of TF, TA, TE and TLA could self-assemble to construct desired Nano-windmill TMA (Fig. S16 in Supporting information). The minor structure alterations of TMA did not affect the activation of FGFR1, Met and EGFR signaling (Fig. S17 in Supporting information). We first developed a strand (AC)-responsive TMA to control Met activation (Fig. S18 in Supporting information). After adding strand AC, TA targeting Met receptors was dissociated from TMA due to the complementary hybridization of TA with AC, leading to the transformation of Nano-windmill into Nano-kite. The recovery of TAMRA fluorescence and native gel confirms the occurrence of this process (Figs. S19–S21 in Supporting information). As TA dissociates from TMA, Met activation caused by TMA was significantly weakened (Figs. 4a and b), suggesting the activation of Met receptors by DNA Nano-windmill was selectively inhibited by complementary strands. Furthermore, we extended the responsiveness of TMA to small molecules (e.g., ATP) (Fig. S22 in Supporting information). Initially, ATP-responsive TA binding on TMA results in TAMRA fluorescence quenching. In the presence of ATP, ATP drives the dissociation of TA from TMA resulting in TAMRA fluorescence recovery (Fig. S23 in Supporting information). This finding was confirmed by the rapid TAMRA fluorescence recovery and native gel (Figs. S24 and S25 in Supporting information). In addition, the conversion process is specific to ATP (Fig. S26 in Supporting information). As we expected, the p-Met expression caused by TMA remarkably decreased due to the transformation of ATP-responsive TMA into Nano-kite TK (Figs. 4c and d). Therefore, the designed TMA that can convert DNA Nano-windmill into DNA Nano-kite using complementary strands or small molecules to reduce the activation of specific receptor.
In summary, we have developed a transformable DNA Nano-windmill for the activation of the heterodimerization receptors between three RTK families. To the best of our knowledge, this is the first example of using controllable DNA Nano-windmill for the activation of heterodimerization three RTK receptors and signaling pathways. The DNA Nano-windmill shows excellent versatility in potential applications owing to (1) the controllability of the DNA Nano-windmill, which can be transformed into DNA Nano-kite by the complementary strands or small molecule to reduce the specific RTK receptor activation caused by TMA, and (2) the wide application of the DNA Nano-windmill, simulates more complex signaling pathways activated by multiple growth factors. Notably, we have simultaneously controlled multiple receptors heterodimerization and activated the receptor signaling pathways by using DNA Nano-windmill, potentially offering a facile and practical nongenetic approach for customized cell collective behaviors in vivo. The DNA nanostructures have shown prominent potential in the field of cancer immunotherapy [42]. The Nano-windmill system has potential applications in inflammatory diseases, such as ulcerative colitis and acute liver injury [39,43]. This strategy represents a new avenue for the smart regulation of cell behavior in cell-based therapy and regenerative medicine.
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Kun Liu: Writing – review & editing, Writing – original draft, Visualization, Validation, Software, Resources, Project administration, Methodology, Investigation, Formal analysis, Data curation, Conceptualization. Junjun Lu: Validation, Software, Methodology, Investigation, Data curation, Conceptualization. Jincheng Lin: Software, Methodology, Data curation, Conceptualization. Yulin Cong: Methodology, Investigation, Formal analysis, Data curation, Conceptualization. Xiongfeng Luo: Visualization, Validation, Supervision, Software. Zihong Huang: Software, Resources, Project administration. Tianyi Zhang: Visualization, Validation, Supervision. Miao Wang: Writing – review & editing. Zhiyong Xie: Writing – review & editing, Funding acquisition. Hao Li: Writing – review & editing, Writing – original draft, Visualization, Supervision, Software, Funding acquisition, Formal analysis.
The authors are grateful to the National Natural Science Foundation of China (Nos. 22104158, 82174104, U1903211), Guangdong Basic and Applied Basic Research Foundation (No. 2023A1515011346), Shenzhen Science and Technology Program (No. SZBH202130), and State Key Laboratory of Chemo/Biosensing and Chemometrics (No. 20230762).
Supplementary material associated with this article can be found, in the online version, at doi:
Z. Du, C.M. Lovly, Mol. Cancer 17 (2018) 58. doi: 10.1186/s12943-018-0782-4
J. Li, L. Wang, J. Tian, et al., Chem. Soc. Rev. 49 (2020) 1545–1568. doi: 10.1039/c9cs00473d
M.A. Lemmon, J. Schlessinger, Cell 141 (2010) 1117–1134. doi: 10.1016/j.cell.2010.06.011
Y. Xie, N. Su, J. Yang, et al., Signal. Transduct. Target. Ther. 5 (2020) 181. doi: 10.1109/cyber50695.2020.9279188
N. Ebrahimi, E. Fardi, H. Ghaderi, et al., Cell. Mol. Life Sci. 80 (2023) 104. doi: 10.1007/s00018-023-04729-4
J. Chen, F. Zeng, S.J. Forrester, et al., Physiol. Rev. 96 (2016) 1025–1069. doi: 10.1152/physrev.00030.2015
S. Zhu, W. Chen, A. Masson, et al., Cell Discov. 10 (2024) 71. doi: 10.23919/pcmp.2023.000101
L. Trusolino, A. Bertotti, P.M. Comoglio, Nat. Rev. Mol. Cell Biol. 11 (2010) 834–848. doi: 10.1038/nrm3012
J.L. Macdonald-Obermann, L.J. Pike, J. Biol. Chem. 289 (2014) 26178–26188. doi: 10.1074/jbc.M114.586826
M.A. Olayioye, R.M. Neve, H.A. Lane, et al., EMBO J. 19 (2000) 3159–3167. doi: 10.1093/emboj/19.13.3159
M.D. Paul, K. Hristova, Chem. Rev. 119 (2019) 5881–5921. doi: 10.1021/acs.chemrev.8b00467
A.E. Lenferink, R. Pinkas-Kramarski, M.L. van de Poll, et al., EMBO J. 17 (1998) 3385–3397. doi: 10.1093/emboj/17.12.3385
A. Chausovsky, H. Waterman, M. Elbaum, et al., Oncogene 19 (2000) 878–888. doi: 10.1038/sj.onc.1203410
L. Chen, L. Fu, J. Sun, et al., Nature 618 (2023) 862–870. doi: 10.1038/s41586-023-06155-9
J.M. Daly, C.B. Jannot, R.R. Beerli, et al., Cancer Res. 57 (1997) 3804–3811.
Y. Du, Y. Lyu, J. Lin, et al., Nat. Nanotechnol. 18 (2023) 818–827. doi: 10.1038/s41565-023-01333-2
D.Y. Zhang, A.J. Turberfield, B. Yurke, et al., Science 318 (2007) 1121–1125. doi: 10.1126/science.1148532
X. Mao, M. Liu, Q. Li, et al., JACS Au 2 (2022) 2381–2399. doi: 10.1021/jacsau.2c00292
Y.J. Chen, B. Groves, R.A. Muscat, et al., Nat. Nanotechnol. 10 (2015) 748–760. doi: 10.1038/nnano.2015.195
M. You, L. Peng, N. Shao, et al., J. Am. Chem. Soc. 136 (2014) 1256–1259. doi: 10.1021/ja4114903
S. Wang, L. Zhang, S. Wan, et al., ACS Nano 11 (2017) 3943–3949. doi: 10.1021/acsnano.7b00373
Q. Gao, Y. Zhao, K. Xu, et al., Angew. Chem. Int. Ed. 59 (2020) 23564–23568. doi: 10.1002/anie.202011198
Y. Zhang, Z. Shuai, H. Zhou, et al., J. Am. Chem. Soc. 140 (2018) 3988–3993. doi: 10.1021/jacs.7b12772
D. Zhu, H. Pei, G. Yao, et al., Adv. Mater. 28 (2016) 6860–6865. doi: 10.1002/adma.201506407
H. Pei, L. Liang, G. Yao, et al., Angew. Chem. Int. Ed. 51 (2012) 9020–9024. doi: 10.1002/anie.201202356
K. He, Y. Li, B. Xiang, et al., Chem. Sci. 6 (2015) 3556–3564. doi: 10.1039/C5SC00371G
N.C. Seeman, Nature 421 (2003) 427–431. doi: 10.1038/nature01406
Z. Wang, L. Wang, H. Zhao, Chin. Chem. Lett. 35 (2024) 109637. doi: 10.1016/j.cclet.2024.109637
R. Nutiu, Y. Li, J. Am. Chem. Soc. 125 (2003) 4771–4778. doi: 10.1021/ja028962o
J. Liu, Z. Cao, Y. Lu, Chem. Rev. 109 (2009) 1948–1998. doi: 10.1021/cr030183i
N. Nakatsuka, K.A. Yang, J.M. Abendroth, et al., Science 362 (2018) 319–324. doi: 10.1126/science.aao6750
W. Tan, M.J. Donovan, J. Jiang, Chem. Rev. 113 (2013) 2842–2862. doi: 10.1021/cr300468w
K. Zhang, H. Gao, R. Deng, et al., Angew. Chem. Int. Ed. 58 (2019) 4790–4799. doi: 10.1002/anie.201809006
K. Zhang, R. Deng, Y. Sun, et al., Chem. Sci. 8 (2017) 7098–7105. doi: 10.1039/C7SC02489D
Y. Liu, Y. Hu, Y. Tan, et al., Chin. Chem. Lett. 36 (2025) 110289. doi: 10.1016/j.cclet.2024.110289
R. Ueki, S. Atsuta, A. Ueki, et al., Chem. Commun. 55 (2019) 2672–2675. doi: 10.1039/c8cc08080a
R. Ueki, A. Ueki, N. Kanda, et al., Angew. Chem. Int. Ed. 55 (2016) 579–582. doi: 10.1002/anie.201508572
K. Liu, Y. Cong, X. Luo, et al., Chin. Chem. Lett. 36 (2025) 109839. doi: 10.1016/j.cclet.2024.109839
Y. Cong, K. Liu, Z. Huang, et al., ACS Chem. Biol. 19 (2024) 1280–1290. doi: 10.1021/acschembio.4c00098
H. Li, M. Wang, T. Shi, et al., Angew. Chem. Int. Ed. 57 (2018) 10226–10230. doi: 10.1002/anie.201806155
H. Li, J. Gao, L. Cao, et al., Angew. Chem. Int. Ed. 60 (2021) 26087–26095. doi: 10.1002/anie.202108210
Q. Chen, Y. Liu, Q. Chen, et al., Small 20 (2024) 2405231. doi: 10.1002/smll.202405231
F. He, M. Wang, J. Wang, et al., Angew. Chem. Int. Ed. 62 (2023) e202305227. doi: 10.1002/anie.202305227

Figure 1 Schematic representation of a DNA Nano-windmill induced FGFR1, Met and EGFR activation to regulate downstream signaling pathways and cellular behavior.

Figure 2 Proof of concept and activation effect of the DNA Nano-windmill (TA3). (a) The scheme of fluorescence chain reaction. After TLM-S and TLE-S was added, it will form a double chain with TLM-C and TLE-C. TFR’s toehold will be exposed with the departure of the TLM-C and TLE-C, which in turn will form a double chain with the TM-F or TE-T, resulting in the separation of the M-B1 or E-B2 and the fluorescence recovery of TM-F or TE-T. (b) Confocal laser scanning microscope (CLSM) images of chain reaction. The A549 cells were incubated with 300 nmol/L TFR, TMR and TER with or without TLM-S and TLE-S (300 nmol/L). Red channel: TAMRA excitation 560 nm/emission 582 nm. Green channel: FAM excitation 494 nm/emission 518 nm. Scale bar: 20 µm. (c) Schematic illustration of DNA Nano-windmill TA3 induced protein-pairing on the cell membrane. (d) Western blot analysis of p-EGFR, p-Met and p-FGFR1 expression in A549 cells with different concentration DNA Nano-windmill TA3 treatment. The glyceraldehyde 3 phosphate dehydrogenase (GAPDH) was used as an internal reference.

Figure 3 The effects of DNA Nano-windmill (TA3) activation on downstream key regulators Erk and Akt in cell motility and proliferation. (a) Schematic representation of signaling pathways with TA3. (b) Western blot analysis of p-Akt and p-Erk expression in A549 cells with different concentration TA3 treatment. (c) Viability of TA3 treated with different concentration for 24 h. Mean ± standard deviation (three technical replicates, n = 5). P values were calculated using t tests on the averages of replicates. (d) Cell mobility regulated by TA3 in A549 cells was determined by wound healing assay. The images were taken at 0 and 24 h. The yellow lines indicate boundaries between cells in the monolayer and the scratched areas uncovered by cells. Scale bar: 200 µm.

Figure 4 The scheme and Western blot of AC or ATP responsible with TMA. (a) Schematic illustration of DNA Nano-kite TK induced protein-pairing on the cell membrane, which TMA react with AC. (b) Western blot analysis of p-Met expression in A549 cells with different concentration AC with TMA (10 nmol/L) treatment. (c) Schematic illustration of DNA Nano-kite TK induced protein-pairing on the cell membrane, which TMA react with ATP. (d) Western blot analysis of p-Met expression in A549 cells with different concentration ATP with TMA (10 nmol/L) treatment.

 扫一扫看文章
扫一扫看文章

扫一扫关注我们
 DownLoad:
DownLoad:
 下载:
下载:
 下载:
下载: