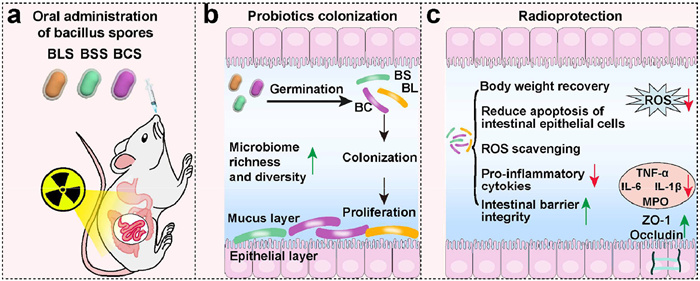
Scheme 1.
Schematic diagram of probiotic spores on prevention of radiation-induce intestinal injury.
Oral administration of Bacillus spores for mitigation of radiation-induced intestinal injury by regulating gut microbiota
Cuixia Zheng , Yueyue Kong , Xinxin Liu , Mengya Niu , Shuang Li , Yang Liu , Lei Wang , Chen Hong

Radiotherapy has long been an essential cancer treatment in clinical practice [1]. Although therapy planning and delivery methods have been improved, radiation still causes considerable damage to healthy tissue, particularly gastrointestinal tract and hematopoietic systems with a large number of rapidly proliferating cells, significantly limiting the clinical application of radiotherapy [2]. During radiotherapy of abdominal or pelvic solid tumors, ionizing radiation can lead serious gastrointestinal dysfunction characterized by symptoms such as vomiting, diarrhea, bleeding, infection and perforation [3,4]. Furthermore, a subset of patients may suffer these symptoms from months to years after radiotherapy and even become permanently, seriously affecting patients’ quality of life [5]. Therefore, it is necessary to induce effective countermeasures to reduce radiation-induced gastrointestinal toxicity.
Radiation-induced intestinal injury is mainly caused by radiation-induced DNA damage, reactive oxygen species (ROS) storms and dysbiosis of gut microbiota [6,7]. At present, antioxidants [8], polyphenols [9], sulfur compounds [10], trace element-based compounds [11], protein-based pharmaceuticals [12], hormone analogues [13] and bone marrow cell-derived extracellular vesicles [14] have been developed as radioprotectants to mitigate radiation induced intestinal injury. Alongside investigational radioprotective candidates, amifostine and palifermin remain the clinically selective normal tissue radioprotectants approved by the U.S. Food and Drug Administration (FDA) [15]. However, the use of radioprotective agents faces numerous challenges. First, majority of radioprotectants cannot meet the requirements for oral administration, because drugs administered orally must pass through stomach and small intestine, leading to their degradation by digestive enzymes and stomach acid [16,17], although it is the most ideal delivery route to achieve intestinal distribution. Moreover, these radioprotectants cannot be explicitly enriched in target organs with high radiosensitivity [18]. High doses of drugs may improve their accumulation in gastrointestinal tract, which inevitably exert off-target pharmacological effects [15,19]. Therefore, radioprotection agents for oral administration must exhibit resistance to destructive effects of gastric acid and digestive enzymes, while simultaneously targeting intestinal tissue.
Substantial evidences have shown correlations of gut microbiota with radiation induced intestinal injury following pelvic irradiation [20]. Ionizing radiation could cause imbalance or dysbiosis of gut microbial community and induce the release of microbe-derived metabolites such as tumor necrosis factor-α (TNF-α), interleukin-6 (IL-6) and IL-1β, which are vital mediators of radiation-induced intestinal damage [21]. The microbiota has been extensively studied for its influence in maintaining gut homeostasis, leading to investigations into its beneficial roles in radioprotection [22,23]. The effectiveness of fecal microbiota transplantation (FMT) demonstrates positive role in patients with radiation-induced injury [24]. In a recent pilot study, Ding et al. confirmed that FMT can alleviate diarrhea, rectal bleeding, abdominal/rectal discomfort, and fecal incontinence in patients with chronic radiation enteritis [25,26]. Consequently, researchers’ attention has been attracted on modifying or preserving microorganisms to mitigate irradiation-induced intestinal injury, including the use of probiotics, FMT and other methods [27,28]. Guo et al. examined the protective role of Lachnospiraceae and Enterococcaceae on gastrointestinal damage following irradiation [29,30]. In Riehl’s research, they found that Lactobacillus rhamnosus GG can activate Toll-like receptor 2 (TLR2) pathway and promote migration of cyclooxygenase 2-expressing mesenchymal stem cells, thus protecting small intestinal epithelium from radiation injury [31]. Nevertheless, the colonization and viability of probiotics as well as fecal microbiota are influenced quite-extent by ionizing radiation in fractionated radiotherapy [32,33]. Therefore, probiotic spores may provide a way out of these dilemmas due to their strong resistance to high temperature, ultraviolet radiation, ionizing radiation, and various chemical substances.
The present study aimed to evaluate the protective effect of spores, separated from three clinically approved probiotics (Bacillus coagulans (BC), Bacillus licheniformis (BL), and Bacillus subtilis (BS)), on acute radiation-induced intestinal injury (Scheme 1). We found that these spores showed higher resistance to X-ray radiation and superior proliferation efficiency than their parent probiotics. In whole abdominal irradiation (WAI) mouse models, these spores exhibited outstanding anti-inflammatory effect by inhibiting the typical proinflammatory factors such as IL-6, IL-1β and TNF-α, so as to alleviate the inflammatory injury of small intestine and colon. In addition, the three spores maintained intestinal flora homeostasis and promoted restoration of barrier integrity. Thus, this study demonstrated the protective and therapeutic potential of probiotic spores in preventing and treating radiation-induced intestinal injury in a preclinical setting.
The probiotic strains BC, BL, and BS, which are approved by the National Medical Products Administration (NMPA), were subjected to sporulation culture to obtain their respective spores (designated as BCS, BLS, and BSS, respectively). Scanning electron microscopy (SEM) images clearly demonstrated that all the three probiotics were rod-shaped of ~2.5 µm, as shown in Fig. S1a (Supporting information). Unlike these probiotics, their spores were short rod-shaped (~1.5 µm) with rough surface observed by both SEM and transmission electron microscopy (TEM) (Fig. S1b in Supporting information).
Oral probiotics often encounter harsh conditions in the gastrointestinal tract, which can compromise their activity [36]. We subsequently investigated the viability of probiotics and spore after exposure to simulated gastric fluid (SGF) and bile salts. As demonstrated in Fig. 1a, the survival rate of BCS, BSS and BLS in SGF were about 65, 35 and 14 times higher than that of BC, BS and BL, respectively. The same trend was observed after bile salt treatment (Fig. 1b), with all the three spores showing >40-fold higher survival rate than their parent probiotics after 6 h of treatment. These results confirmed that thick hydrophobic coat of spores could effectively enhance probiotics bacillus resistance to harsh gastrointestinal environment, thereby providing foundation for probiotics successful colonization in the intestine.
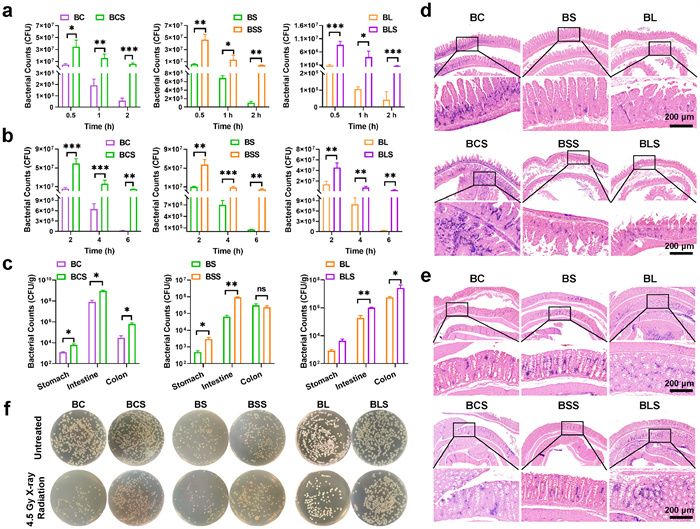
Colonization and proliferation efficiency were prerequisites for probiotics or spores to prevent and treat radiation-induced intestinal injury. To quantify the quantity of colonized probiotics in the gastrointestinal tract, bacterial colony counts and gram staining were performed at 48 h following oral administration. All the work performed on animals was in accordance with the Guidelines for Care and Use of Laboratory Animals of Zhengzhou University, and the experiments were approved by the Animal Ethics Committee of Zhengzhou University. As shown in Fig. 1c, the survival rate of spores in the mice’s gastrointestinal tract was significantly higher than that of Bacillus. Notably, the numbers of BCS, BSS and BLS in the intestine were approximately 11.2, 9.2 and 5.5 times higher than that of their parent probiotics. Similar results were obtained in the colon of BCS and BLS, with BCS and BLS being approximately 20.2 and 4.8 times higher than the BC and BL group, respectively. Next, the colonization efficiency of the three probiotics and their spores was observed directly through gram staining of intestine and colon tissues. As shown in Figs. 1d and e, a large number of BC and BCS colonized in small intestine, but BL and BLS colonization were observed in colon. Whereas relatively few BS and BSS colonization were detected both in small intestine and colon, which mainly because that BS was transient flora and regulated intestinal function by producing metabolites. Consistently with the colony count results, the colonization rate of probiotics was significantly lower than corresponding spores. These results demonstrated that spores could effectively pass harsh gastric acid environment, further conforming strong stress resistance of spore.
It was reported that ionizing irradiation could inactivate probiotics by specific way. While spores can resist ionizing radiation due to their unique structure. As shown in Fig. 1f, a large number of probiotics (including BC, BS and BL) lost proliferation activity after irradiation with 4.5 Gy X-ray. On the contrary, the colony number of probiotic spores showed no significant difference before and after X-ray irradiation, suggesting good germination activity of these spores. In this sense, probiotic spores could serve as an advantageous bioactive carrier for oral drug delivery system.
Subsequently, comprehensive evaluation, including complete blood count, blood biochemical indices, and hematoxylin-eosin (H&E), was conducted to evaluate in vivo biocompatibility of these three spores. As indicated in Fig. S2 (Supporting information), H&E staining revealed no changes in the major organs after oral administration of these three bacillus spores compared to the control group. Similarly, liver and kidney function biomarkers, as well as representative hematological parameters of the spores-treated groups were within the normal range relative to healthy mice in the control group as shown in Fig. S3 (Supporting information).
In the settings of clinical radiotherapy, ionizing irradiation could induce various abnormalities of intestinal tissue. Next, we evaluated the radioprotective effect of probiotic spores in vivo by visually monitoring various representative indicators in small intestine and colon in an abdominal irradiation BALB/c mice model (Fig. 2a). Following X-ray radiation, the mice were housed in sterile autoclaved cages and provided standard chow and water ad libitum. Body weight of mice was monitored through 8 days after radiation. As indicated in Fig. 2b, the mice’s weights in the X-ray treated group decreased successively in the first 5 days after irradiation and mice started to recover bodyweight on day 6. Furthermore, the body weight of mice maintained a slowly increasing trend. The three probiotic spores could prevent the bodyweight loss in different degrees and significantly improve weight gain. Notably, the average body weight almost returned to the degree of healthy mice in normal group on the 8th day (Fig. S4 in Supporting information). According to Fig. 2c. mice treated with X-ray radiation exhibited average reductions in colon length of 24.2%, compared with the normal group. The colon length in the three spore groups were similar to that of healthy mice in normal group and significant longer than that in X-ray treated group. This result confirmed the protective role of probiotics bacillus spores against radiation-induced damage.
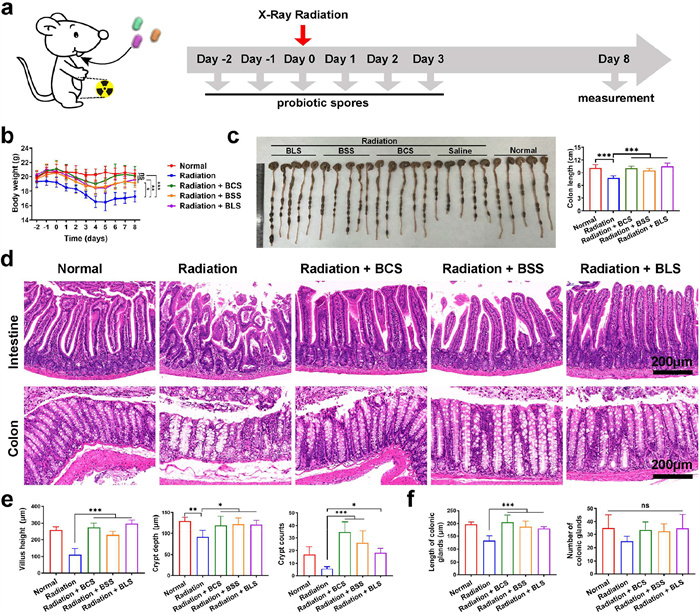
Next, pathological analysis of small intestine and colon was executed to directly verify the radioprotective ability of these spores. Some typical abnormalities of radiation-induced intestinal injury were observed in radiation group, such as reduced crypt (including crypt depth and count) and shortened intestinal villi (Figs. 2d and e), common epithelial responses to injury. Moreover, X-ray treated mice showed more colonic gland distortion and atrophy, with gaps between colonic gland bases and muscularis mucosa, as shown in Figs. 2d and f. By contrast, the flattened intestinal villi and damaged intestinal crypts were rescued by administration of the three probiotic spores. In addition, colonic gland of mice in BCS, BSS and BLS groups remained closely attached to the muscularis mucosa. Similarly, negligible apoptotic fluorescence was detected in radiation + BCS, radiation + BSS and radiation + BLS groups, whereas radiation group showed a large number of necrotic cells, both in small intestine and colon (Fig. S5 in Supporting information), demonstrating that the three bacillus spores could obviously inhibit the development of radiation-induced intestinal injury.
Additionally, more details of histopathology were analyzed. As shown in Fig. 3a, the representative H&E images indicated that X-ray radiation induced intestinal crypts atrophy and vanish (black arrow), ulcer lesions appearance (green arrow), glands hyperplasia (blue arrow) and inflammatory infiltration (yellow arrow). While slight glands damage and inflammatory infiltration were also observed in radiation + BCS, radiation + BSS and radiation + BLS groups. In addition, few ulcers suggested significant reduction in the inflammation/mucosal damage in the three groups. Moreover, the increased myeloperoxidase (MPO) activity was dramatically suppressed after treatment with BCS, BSS and BLS (Fig. 3b). Meanwhile, the typical proinflammatory factors such as IL-6, TNF-α and IL-1β were decreased in radiation + BCS, radiation + BSS and radiation + BLS groups (Figs. 3c–e), highlighting decreased tissue inflammation in the three groups. One of the main intestinal damages caused by radiation is ROS-induced DNA double-strand break [15,37]. As shown in Fig. 3f, all the three spores with a powerful ROS scavenging ability and can reduce ROS level in intestinal microenvironment after X-ray radiation. According to Fig. 3g, remarkable γ-H2AX was observed in the X-ray radiation group. By contrast, the γ-H2AX level in radiation + BCS, radiation + BSS and radiation + BLS groups was significantly decreased, indicating that bacillus spores could effectively reduce DNA damage. The anti-inflammatory effect and ROS-scavenging activity of bacillus spores played key role in preventing DNA damages.

We further assessed the impact of spores on intestinal barrier functions, since any breaking or even slightly perturbing of epithelial barriers might lead to serious pathological consequences. As shown in Figs. S6 and S7 (Supporting information), expressions of tight junction-associated protein zona occludens 1 (ZO-1) and occludin dramatically decreased after X-ray irradiation. Encouragingly, destruction of intestinal epithelial barrier was rescued by administration of these probiotic spores, which might be related to regulation of tight junction by probiotics and calcium ions released during spore germination [34,35].
In addition to anti-inflammatory and regulation of intestinal epithelial barrier, the crosstalk between intestinal flora and spores (probiotics, actually) plays a crucial role in protection of radiation induced intestinal injury. Subsequently, 16S ribosomal DNA (rDNA) sequencing technology was conducted to investigate the impact of ionizing radiation on intestinal microbiota. X-ray radiation and the three probiotic spores had almost no impact on bacterial richness, as observed coverage diversity indexes in Fig. S8 and operational taxonomic units (OTUs) richness in Fig. S9 (Supporting information). Although alpha diversity (Fig. S10 in Supporting information) presented that there was no obvious distinction in community richness (the index Chao) and community diversity (the index Shannon) between different groups. The Venn diagram (Fig. 4a) illustrated that BCS, BSS and BLS treatment increased species richness up to 468, 462 and 449 respectively compared to radiation group (418). Moreover, based on beta diversity, NonMetric Multi-Dimensional Scaling (NMDS) analysis in Fig. 4b demonstrated that the radiation group exhibited a distinct gut microbiota profile, while radiation + BCS, radiation + BSS and radiation + BLS groups clustered more closely to the healthy mice in normal group, suggesting that bacillus spores treatment reestablished a healthier microbiota.
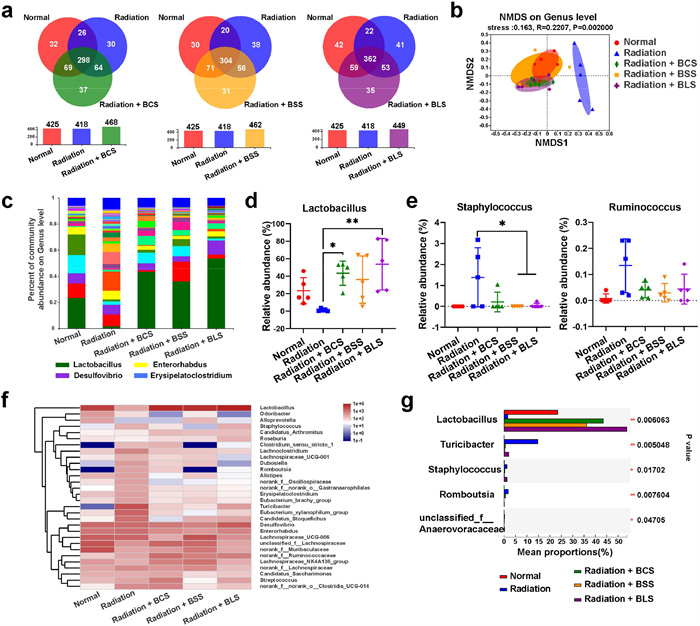
Next, species composition and relative abundance of gut microbiota were analyzed based on the annotation degree of all sample sequences. X-ray exposure resulted in changes in the compositions of bacterial communities at genus level across different groups (Figs. 4c and f). Notably, the relative abundance of Lactobacillus, a kind of typical probiotics in the intestine, displayed drastic decrease after X-ray irradiation (Fig. 4d). While the abundance of the pathogenic bacteria, such as Staphylococcus and Ruminococcus, showed a significant increase after irradiation (Fig. 4e). By contrast, the abundance of probiotics showed positive regulation and pathogenic bacteria was considerably lower in radiation + BCS, radiation + BSS and radiation + BLS groups. In particular, the relative abundance of Lactobacillus exhibited an upward trend compared to that of healthy mice in the control group (Fig. 4g and Fig. S11 in Supporting information). These results demonstrated that all of the three spores upregulated gut microbiota homeostasis, among which BLS exhibited a more forceful regulatory effect.
In this study, considering the vulnerability of probiotics in harsh gastric acid and ionizing irradiation environment, we employed spores of three clinically approved probiotics to conduct a series of studies on the protection of acute radiation-induced intestinal injury. We discovered that spores showed stronger resistance to X-ray radiation and higher colonization efficiency than their parent probiotics. These spores showed outstanding performance in gastrointestinal radioprotection though inhibiting inflammation, maintaining intestinal flora homeostasis and improving intestinal barrier function. Moreover, our studies showed that relative abundance of Lactobacillus upregulated after oral administration of the three spores, which might be associated with intestinal repair after X-ray radiation. In summary, spores of bacillus might have great potential in clinical application for intestinal radioprotection.
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Cuixia Zheng: Writing – review & editing, Writing – original draft, Funding acquisition, Data curation, Conceptualization. Yueyue Kong: Methodology, Investigation. Xinxin Liu: Validation, Supervision, Software. Mengya Niu: Investigation, Formal analysis. Shuang Li: Investigation. Yang Liu: Supervision. Lei Wang: Writing – review & editing, Project administration. Chen Hong: Software, Methodology, Investigation.
This work was supported by the National Natural Science Foundation of China (Nos. 82272847, 82303529), China Postdoctoral Science Foundation (No. 2023M730971), Henan Provincial Medical Science and Technology Public Relations Program Provincial-Ministerial Co-Construction Youth Project (No. SBGJ202303041).
Supplementary material associated with this article can be found, in the online version, at doi:
D. Schaue, W.H. McBride, Nat. Rev. Clin. Oncol. 12 (2015) 527–540. doi: 10.1038/nrclinonc.2015.120
R. Klaus, M. Niyazi, B. Lange-Sperandio, Radiat. Oncol. 16 (2021) 43. doi: 10.1186/s13014-021-01764-y
Z.N. Qiao, Z.Y. Li, Y.L. Shi, et al., Int. J. Biol. Macromol. 280 (2024) 135809. doi: 10.1016/j.ijbiomac.2024.135809
J. Fu, Z. Yang, S. Melemenidis, et al., Int. J. Radiat. Oncol. Biol. Phys. 119 (2024) 1001–1010. doi: 10.1016/j.ijrobp.2023.12.032
Q.Q. Xu, P.Y. Zhang, X.Y. Han, et al., Int. J. Biol. Macromol. 241 (2023) 124476. doi: 10.1016/j.ijbiomac.2023.124476
X.Z. Zhu, Y. Li, L.L. Yue, et al., Radiat. Res. 199 (2023) 252–263. doi: 10.1038/s41423-022-00969-9
J.J. Fan, B.W. Lin, M. Fan, et al., Front. Oncol. 12 (2022) 888962. doi: 10.3389/fonc.2022.888962
K.N. Mishra, B.A. Moftah, G.A. Alsbeih, Biomed. Pharmacother. 106 (2018) 610–617. doi: 10.1016/j.biopha.2018.06.150
J.N. Xie, Y. Yong, X.H. Dong, et al., ACS Appl. Mater. Interfaces 9 (2017) 14281–14291. doi: 10.1021/acsami.7b02622
P. Shan, J. Liao, J.Y. Li, et al., Chin. Chem. Lett. 35 (2024) 108545. doi: 10.1016/j.cclet.2023.108545
C.Y. Wang, M.R. Zhao, J.N. Xie, et al., Adv. Healthc. Mater. 12 (2023) e2301758. doi: 10.1002/adhm.202301758
T. Zhang, L. Shi, Y. Li, et al., J. Radiat. Res. 62 (2021) 46–57. doi: 10.1093/jrr/rraa093
L.W. Xie, S. Cai, H.Y. Lu, F.L. Tang, R.Q. Zhu, et al., Gut Microbes 16 (2024) 2347722. doi: 10.1080/19490976.2024.2347722
N.N. He, M.X. Dong, Y.X. Sun, et al., Theranostics 14 (2024) 5492–5511. doi: 10.7150/thno.97623
D.X. Zhang, D.N. Zhong, J. Ouyang, et al., Nat. Commun. 13 (2022) 1413. doi: 10.1080/0142159x.2022.2100251
Q.L. Song, J.F. Yang, X.C. Wu, et al., J. Nanobiotechnology 21 (2023) 302. doi: 10.1186/s12951-023-01995-z
J.F. Yang, K. Wang, S.X. Sun, et al., Chin. Chem. Lett. 36 (2025) 110180. doi: 10.1016/j.cclet.2024.110180
M. Zeeshan, H. Ali, S. Khan, S.A. Khan, B. Weigmann, Int. J. Pharm. 558 (2019) 201–214. doi: 10.1016/j.ijpharm.2018.12.074
N.P. Praetorius, T.K. Mandal, J. Pharm. Pharmacol. 60 (2008) 809–815. doi: 10.1211/jpp.60.7.0001
L.L. Zhang, J.Y. Xu, Y.F. Xing, et al., Microbiol. Res. 286 (2024) 127821. doi: 10.1016/j.micres.2024.127821
L.F. Tang, X.M. Ma, L.W. Xie, et al., Mol. Nutr. Food Res. 67 (2023) e2300232. doi: 10.1002/mnfr.202300232
G.H. Lu, D.W. Gao, W. Jiang, et al., Eur. J. Nucl. Med. Mol. Imaging 51 (2024) 2761–2773. doi: 10.1039/d4cy00024b
X.Q. Fan, C.T. Mai, L. Zuo, et al., Acta Pharm. Sin. B 13 (2023) 1164–1179. doi: 10.1016/j.apsb.2022.10.016
L. Chen, J.E. Wilson, M.J. Koenigsknecht, et al., Nat. Immunol. 18 (2017) 1270. doi: 10.1038/ni1117-1270b
Y.Y. Chen, Q. Zeng, Y.Y. Luo, et al., Ecotoxicol. Environ. Saf. 283 (2024) 116834. doi: 10.1016/j.ecoenv.2024.116834
M. Acharya, B.S. Venkidesh, K.D. Mumbrekar, Life Sci. 353 (2024) 122921. doi: 10.1016/j.lfs.2024.122921
C. Zheng, M. Niu, Y. Kong, et al., J. Nanobiotechnology 22 (2024) 303. doi: 10.1007/978-981-99-8469-5_24
S.L. Zhang, Z.Y. Wang, J. Jiang, G.X. Feng, S.J. Fan, Food Funct. 15 (2024) 3522–3538. doi: 10.1039/d3fo05422e
H. Guo, W.C. Chou, Y.J. Lai, et al., Science 370 (2020) 549–560.
J. Gu, L. Zhao, Y.Z. Chen, et al., Biomed. Pharmacother. 146 (2022) 112496. doi: 10.1016/j.biopha.2021.112496
T.E. Riehl, D. Alvarado, X.P. Ee, et al., Gut 68 (2019) 1003–1013. doi: 10.1136/gutjnl-2018-316226
Y.F. Chen, S.C. Li, E.Y. Huang, J. Radiat. Res. 65 (2024) 55–62. doi: 10.1093/jrr/rrad084
Q.L. Song, J.F. Yang, Y. Li, et al., Nano Today 58 (2024) 102375. doi: 10.1016/j.nantod.2024.102375
L.D. Knecht, P. Pasini, S. Daunert, Anal. Bioanal. Chem. 400 (2011) 977–989. doi: 10.1007/s00216-011-4835-4
B. Huang, X.M. Liu, Z.Y. Li, et al., Chem. Eng. J. 414 (2021) 128805. doi: 10.1016/j.cej.2021.128805
J.L. Yang, S.C. Tan, S.C. Ge, et al., Proc. Natl. Acad. Sci. U. S. A. 121 (2024) e2403417121. doi: 10.1073/pnas.2403417121
J.L. Yang, M.Y. Peng, S.C. Tan, et al., ACS Central Sci. 9 (2023) 1327–1341. doi: 10.1021/acscentsci.3c00227

Scheme 1 Schematic diagram of probiotic spores on prevention of radiation-induce intestinal injury.

Figure 1 (a, b) Equal amounts of bacillus and spores were exposed at 37 ℃ to (a) SGF for 2 h and (b) bile salts for 6 h. Bacterial quantities were determined using plate counting at the indicated time intervals, CFU, colony forming units. (c) Enumeration of bacillus and spores in the stomach, intestine and colon at 48 h following oral administration, respectively. (d, e) Gram staining of (d) small intestine and (e) colon of bacillus and their spores. (f) The bacteria count of different groups after X-ray radiation. Data are presented as mean ± SD (n = 3). P values were calculated by one-way or two-way ANOVA with a Tukey post-hoc test. *P < 0.05, **P < 0.01, ***P < 0.001. ns, not significant.

Figure 2 Radioprotective effects of BCS, BSS and BLS in BALB/c mouse model. (a) Therapeutic schedule. (b) The body weight changes in mice. (c) Representative colon images and Colon lengths of mice subjected to indicated treatments. (d) Representative images of H&E staining in intestinal and colonic tissues. (e) Villus height, crypt depth and crypt count of small intestine. (f) Length and number of colonic glands. Data are presented as mean ± SD (n = 5). P values were calculated by one-way or two-way ANOVA with a Tukey post-hoc test. *P < 0.05, **P < 0.01, ***P < 0.001.

Figure 3 Evaluation of radioprotective effect in vivo. (a) H&E staining images of intestine and colon. (b) Detection of MPO activity in intestine and colon. (c–e) Detection of pro-inflammatory cytokines (c) IL-6, (d) TNF-α and (e) IL-1β. (f) Representative images of ROS immunofluorescence in intestinal and colonic tissues. (g) Representative IHC images of γ-H2AX staining in intestinal and colonic tissues. Data are presented as mean ± SD (n = 3). P values were calculated by one-way ANOVA with a Tukey post-hoc test. *P < 0.05, **P < 0.01, ***P < 0.001.

Figure 4 16S rDNA gene sequencing analysis. (a) Venn chart to count the number of common and unique species in multiple groups. (b) Beta diversity. (c) Relative abundance of gut microbiome. Genus-level taxonomy was presented as a percentage of total sequences. (d) Relative abundance of intestinal probiotics Lactobacillus. (e) Relative abundance of Staphylococcus and Ruminococcus in the intestine. (f) Heatmap of the relative abundance of family-level taxa for each mouse. (g) Relative abundance of different treatments. Data are presented as mean ± SD (n = 5). P values were calculated by one-way ANOVA with a Tukey post-hoc test. *P < 0.05, **P < 0.01.

 扫一扫看文章
扫一扫看文章

扫一扫关注我们
 DownLoad:
DownLoad:
 下载:
下载:
 下载:
下载: