 图1
4种不同序列的肽片段在37 ℃环境下培养3 d后的AFM图像
Figure1.
AFM images of four peptide fragments incubated at 37 ℃ for three days
图1
4种不同序列的肽片段在37 ℃环境下培养3 d后的AFM图像
Figure1.
AFM images of four peptide fragments incubated at 37 ℃ for three days
引用本文:
陆澄, 李琳, 鄢鹏, 张楠, 陈武超, 张公军, 周星飞. 二价铜离子对人胰岛淀粉样蛋白(hIAPP)(11~28)多肽聚集的影响[J]. 应用化学,
2018, 35(2): 147-153.
doi:
10.11944/j.issn.1000-0518.2018.02.170378

Citation: LU Cheng, LI Lin, YAN Peng, ZHANG Nan, CHEN Wuchao, ZHANG Gongjun, ZHOU Xingfei. Effect of Copper Ions on the Aggregation of Human Islet Amyloid Polypeptide(11~28)[J]. Chinese Journal of Applied Chemistry, 2018, 35(2): 147-153. doi: 10.11944/j.issn.1000-0518.2018.02.170378
Citation: LU Cheng, LI Lin, YAN Peng, ZHANG Nan, CHEN Wuchao, ZHANG Gongjun, ZHOU Xingfei. Effect of Copper Ions on the Aggregation of Human Islet Amyloid Polypeptide(11~28)[J]. Chinese Journal of Applied Chemistry, 2018, 35(2): 147-153. doi: 10.11944/j.issn.1000-0518.2018.02.170378
二价铜离子对人胰岛淀粉样蛋白(hIAPP)(11~28)多肽聚集的影响
摘要:
人胰岛淀粉样蛋白(hIAPP)与Ⅱ型糖尿病(T2DM)密切相关,被认为是导致胰岛β细胞凋亡的致病因素之一,研究发现环境因素(如金属离子、pH值和温度等)对hIAPP的聚集过程有很大影响。本文采用多种生物物理的实验方法,研究了二价铜离子对hIAPP及其片段聚集的影响。原子力显微镜(AFM)和硫代黄素T(ThT)荧光的测量表明,铜离子能够明显地抑制hIAPP(11~28)聚集成纤维,其抑制程度随铜离子浓度的增加而明显加剧。显微傅里叶变换红外光谱(Micro-FTIR)的结果表明,铜离子能够抑制hIAPP多肽中α螺旋结构向β折叠的转变。另外,氨基酸定点突变实验结果表明,hIAPP(11~28)中的组氨酸(His18)可能对多肽的聚集行为和金属铜离子的相互作用起到了决定性的影响。
English
Effect of Copper Ions on the Aggregation of Human Islet Amyloid Polypeptide(11~28)
Abstract:
Human islet amyloid polypeptide(hIAPP) is closely associated with type 2 diabetes mellitus(T2DM), which is one of possible pathogenic factors of islet beta cell apoptosis. It has been suggested that the environmental factors(such as metal ions, pH and temperature) have great effects on the aggregation process of hIAPP. In this study, we investigate the influence of copper(Ⅱ) ions on the aggregation of hIAPP and its fragments by a variety of biophysical methods. Atomic force microscope(AFM) and thioflavin T(ThT) fluorescence measurements show that the copper ions can inhibit hIAPP(1~37) and hIAPP(11~28) to aggregate into fibers. In addition, the micro-Fourier transform infrared spectroscopy(Micro-FTIR) shows that copper ions can restrain the transition from alpha helices structure to beta sheets formation during the peptide incubation. By the single amino acid mutation experiment, we speculate that the His18 in hIAPP(11~28) may have a dominant effect on the aggregation behavior and the interaction with copper ions.
-
Key words:
- copper(Ⅱ) ions
- / human islet amyloid protein
- / type 2 diabetes
-
淀粉样多肽的错误折叠(即从蛋白单体到纤维的形成)与许多退行性疾病有关,如Ⅱ型糖尿病(T2DM),阿尔茨海默氏病(AD)和帕金森氏病(PD)[1-4]。T2DM是一种常见的糖尿病,通常认为它的发病与人胰岛淀粉样多肽(hIAPP,也称为人胰淀素)的聚集有密切关系,高达90%的T2DM患者的胰岛β细胞中存在这种淀粉样沉积[5],这与老年痴呆症中特征性β折叠的淀粉样蛋白聚集很类似[6-7]。hIAPP是由37个残基组成的多肽,从可溶性的α螺旋/无规卷曲到不溶性的β折叠构象的异常转变就有可能产生β细胞毒性[8-10],导致胰岛β细胞功能障碍和死亡。研究表明,淀粉样多肽的聚集不仅与氨基酸种类和序列密切相关,还与非遗传因素有关。例如,金属离子、pH值、纳米颗粒、温度和空间约束等[11-13]。
铜是人体内普遍存在的一种微量元素,成人每天需摄入1~3 mg,正常条件下人脑中自由铜离子浓度非常低,对神经功能的影响并不显著性。但是T2DM和帕金森症等一些神经退行性疾病的病人体内的铜离子浓度比正常人高一个数量级[14-15]。健康人血浆铜离子浓度为(13.08±2.53) μmol/L,糖尿病患者或者阿尔茨海默病患者的Aβ斑块中铜离子浓度高达400 μmol/L[16],因此我们选择3种铜离子浓度(10、50和100 μmol/L)来研究其对多肽聚集的影响。目前研究表明,铜离子和hIAPP相互作用改变了多肽的聚集途径和聚集产物[17-18]。鉴于hIAPP不同种类的氨基酸对铜离子的作用不同[19],我们合成了不同序列的3个肽片段,分别是hIAPP(1~11)、hIAPP(11~28)和hIAPP(28~37)。利用原子力显微镜(AFM)和硫代黄素T(ThT)荧光研究了3种hIAPP片段的纤维化趋势及金属铜离子对其聚集的作用。
1 实验部分
1.1 仪器和试剂
NanoscopeⅤ型原子力显微镜(AFM,德国Bruker公司);Fluoroskan Ascent型标准型荧光光谱仪(美国Thermo Scientific公司);Cary660型傅里叶变换红外光谱仪(FTIR,美国Agilent公司)。
hIAPP购于美国AnaSpec公司,纯度≥96%;hIAPP(1~11)、hIAPP(11~28)和hIAPP(28~37)购于上海强耀生物科技有限公司,纯度≥98.47%;hIAPP(H18R)和hIAPP(H18S)购于上海吉尔生化有限公司,纯度≥98.76%;氯化铜和硫代黄素T(ThT)荧光购于上海Sigma-Aldrich公司。磷酸盐缓冲液(PBS,磷酸氢二钠8 mmol/L、氯化钠136 mmol/L、磷酸二氢钾2 mmol/L、氯化钾2.6 mmol/L,pH值~7.4)购于国药化学试剂有限公司。实验用水为Milli-Q水,其电导率为18.2 MΩ/cm。为了制备新鲜的肽溶液,首先将1 mg hIAPP以及各种多肽片段溶解在超纯水(18.2 MΩ/cm)中,配制浓度为4 mmol/L的储存溶液,保存在-20 ℃下,使用前用PBS稀释至所需浓度。将ThT荧光试剂用超纯水稀释成浓度4 mmol/L的溶液,保存在4 ℃。实验时将储存的ThT试剂稀释为终浓度50 μmol/L。
1.2 AFM观察
首先,用PBS稀释hIAPP以及各种不同序列的片段短肽储存溶液,与不同浓度的铜离子混合,使多肽的最终浓度均为100 μmol/L,然后在37 ℃恒温箱中培养3 d。接着,将4~5 μL培养过的样品滴加在新解离的云母片上,吸附3 min后在室温下自然晾干,用于AFM观察,实验中AFM在空气中采用轻敲模式观察样品的表面形貌[20]。AFM使用的探针型号为NSC11(三角形悬臂,爱沙尼亚MikroMash公司),其典型共振频率为330 kHz,典型弹性常数为50 N/m,扫描速率为1 Hz。实验数据使用NanoScope V1.20软件分析。
1.3 ThT荧光方法
为了研究hIAPP和金属铜离子相互作用的动力学性质,将hIAPP(11~28)溶液分别加入ThT荧光分子和不同浓度的CuCl2,使多肽和ThT的终浓度分别为100和50 μmol/L,铜离子的浓度分别是0、10、50和100 μmol/L。这些样品被滴加到无盖平底96孔板(美国Corning公司3925型多孔板)中进行荧光测量。每个样品重复3次,每5 min采集1次荧光数据。实验中,所采用的荧光光谱仪的激发波长和发射波长分别为440和485 nm。
1.4 Micro-FTIR光谱方法
对于Micro-FTIR测量,分辨率为0.4 cm-1,所需样品制备与上述AFM实验相同。hIAPP(11~28)(终浓度为100 μmol/L)分别和两种不同浓度的铜离子混合(10和100 μmol/L),37 ℃培养3 d。在金刚石样品台表面滴加2~3 μL的多肽溶液,然后用红外线灯烘干。FTIR分析的数据集中在酰胺Ⅰ模式,这是肽二级结构的一个的重要指标[21-22]。
2 结果与讨论
2.1 hIAPP不同片段的聚集行为
图 1A是hIAPP在37 ℃下不加金属铜离子时培养3 d的AFM图,我们可以观察到绝大部分hIAPP单体均聚集成了很长的纤维(平均长度约为(2.53±0.69) μm,纤维统计数目为70根)。图 1B~1D是3种不同短肽,即hIAPP(11~28)、hIAPP(1~11)和hIAPP(28~37)在相同实验条件下聚集的形貌图。在3种肽片段中,只有hIAPP(11~28)能够形成纤维状聚集体,绝大部分蛋白单体聚集成了纤维,其长度大约是(1.81±0.47) μm,而hIAPP(1~11)和hIAPP(28~37)大部分形成了颗粒状的聚集体。说明只有全序列的hIAPP和hIAPP(11~28)片段具有很强的聚集能力,而hIAPP(1~11)和hIAPP(28~37)不具有聚集行为。因此,实验结果表明hIAPP(11~28)片段是hIAPP多肽聚集的核心区域[23]。
图1
4种不同序列的肽片段在37 ℃环境下培养3 d后的AFM图像
Figure1.
AFM images of four peptide fragments incubated at 37 ℃ for three days
2.2 铜离子对hIAPP聚集行为的影响
用AFM研究了不同浓度的铜离子对hIAPP(11~28)聚集的影响。通过AFM的轻敲模式获得沉积在云母衬底上的样品的形貌。hIAPP(11~28)与不同浓度的铜离子与hIAPP(11~28)共同培养时,其聚集行为被明显抑制了。当铜离子的浓度为10 μmol/L时,的短纤维形成了,如图 2A所示,其长度大约是(0.61±0.20) μm;随着铜离子浓度的增加,形成的纤维长度进一步缩短到(0.39±0.08) μm;当铜离子浓度达到100 μmol/L时,几乎观察不到纤维的形成,只有大块的颗粒状聚集体,说明铜离子可以很有效地抑制hIAPP(11~28)的纤维化。
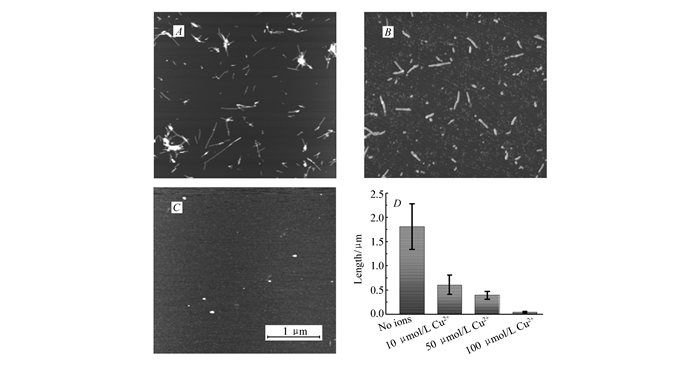 图2
铜离子对hIAPP(11~28)纤维化的影响。A~C是hIAPP (11~28)分别与不同浓度的氯化铜混合培养3 d后的AFM形貌及纤维长度统计图
Figure2.
AFM images of hIAPP(11~28) incubated at 37 ℃ for three days in the presence of copper ions at different concentrations(A~C), and the statistical diagram(D) of lengths of hIAPP(11~28) aggregation incubated with copper ions at different concentration
图2
铜离子对hIAPP(11~28)纤维化的影响。A~C是hIAPP (11~28)分别与不同浓度的氯化铜混合培养3 d后的AFM形貌及纤维长度统计图
Figure2.
AFM images of hIAPP(11~28) incubated at 37 ℃ for three days in the presence of copper ions at different concentrations(A~C), and the statistical diagram(D) of lengths of hIAPP(11~28) aggregation incubated with copper ions at different concentration
为了进一步研究铜离子影响hIAPP(11~28)聚集的动力学过程,利用ThT荧光光谱对hIAPP(11~28)的聚集过程进行监测。ThT荧光是用于检测与蛋白质错误折叠相关的淀粉样多肽纤维化的常用方法,其原理是ThT荧光分子可以与蛋白纤维中的β片层结构特异性结合,并在波长482 nm附近产生新的发射峰[24-25]。
如图 3所示,对于未引入铜离子的hIAPP(11~28),ThT荧光曲线由初始的滞后期、快速生长期和具有最大荧光强度的最终平稳期3个阶段组成,这与文献报道的数据一致[26]。由图 3可知,在hIAPP(11~28)溶液中加入不同浓度的铜离子后,ThT荧光强度变弱了。当加入的铜离子浓度较低时(10和50 μmol/L),尽管在整个实验过程中我们还是能够观察到典型的多肽聚集的三阶段曲线,但荧光强度明显减弱了,说明铜离子抑制了hIAPP(11~28)的聚集。这与我们之前AFM的观察结果是吻合的。当铜离子浓度增加到100 μmol/L时,几乎观察不到ThT荧光的变化,即使经过几天的培养后,ThT荧光强度也很弱,说明引入高浓度的铜离子后多肽的折叠路径完全改变了[27]。
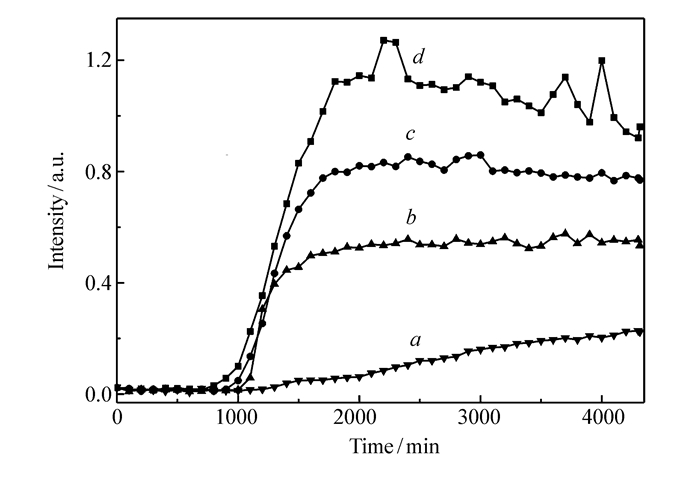 图3
ThT荧光光谱检测铜离子影响hIAPP(11~28)聚集过程
Figure3.
Kinetic analysis of hIAPP (11~28) fibril formation monitored by ThT fluorescence in the absence and presence of different concentration Cu2+
图3
ThT荧光光谱检测铜离子影响hIAPP(11~28)聚集过程
Figure3.
Kinetic analysis of hIAPP (11~28) fibril formation monitored by ThT fluorescence in the absence and presence of different concentration Cu2+
进一步利用显微FTIR(Micro-FTIR)来揭示淀粉样多肽纤维化过程的构象变化,Micro-FTIR是一种可以提供关于酰胺Ⅰ带(1600~1700 cm-1)和酰胺Ⅲ带(1220~1330 cm-1)分子结构信息的光谱技术,酰胺带的不同频率与蛋白质的二级结构成分有密切的关系。
使用Micro-FTIR监测在铜离子缺失和存在情况下的hIAPP(11~28)的二级结构。图 4为加入和未加入铜离子的hIAPP(11~28)在37 ℃下培养3 d后典型的Micro-FTIR光谱。对于不加铜离子的hIAPP(11~28),仅在1629 cm-1附近出现一个波峰,对应于典型的β-折叠结构[28],这意味着hIAPP(11~28)聚集形成纤维的二级结构以β折叠为主。相比之下,当引入10 μmol/L的铜离子后,在1652 cm-1附近出现了一个小肩峰,这个峰可以归因于无规卷曲或螺旋结构[29]。实验结果表明,当引入较低浓度的铜离子时,hIAPP(11~28)聚集体的二级结构主要为β-折叠和α-螺旋共存。随着铜离子浓度进一步增加至100 μmol/L,在1629 cm-1处的吸收峰强度急剧下降,说明聚集体中β折叠结构明显减少,主要是以α-螺旋结构存在。这些结果表明,hIAPP(11~28)的聚集在铜离子存在下受到明显的抑制。
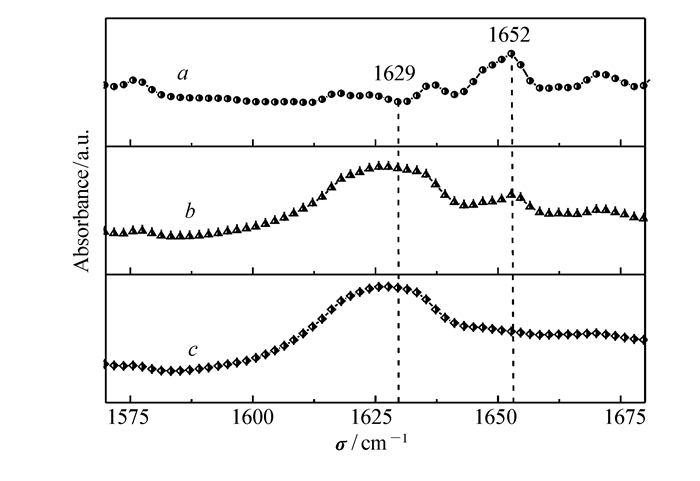 图4
hIAPP(11~28)在加入不同浓度的铜离子后在37 ℃下培养3 d后的Micro-FTIR图
Figure4.
Micro-FTIR spectra of hIAPP(11~28) incubated at 37 ℃ for three days in the absence or in the presence of Cu2+ ions at different concentrations
图4
hIAPP(11~28)在加入不同浓度的铜离子后在37 ℃下培养3 d后的Micro-FTIR图
Figure4.
Micro-FTIR spectra of hIAPP(11~28) incubated at 37 ℃ for three days in the absence or in the presence of Cu2+ ions at different concentrations
2.3 hIAPP中组氨酸的重要作用
Carolina等[30]曾利用固体核磁共振实验,发现铜离子能够和hIAPP中的组氨酸(His18)有很强的相互作用。实验中发现铜离子能够抑制hIAPP(11~28)的聚集,这很可能是因为片段中的(His18)和铜离子结合影响了其局域的结构[31],进而影响了hIAPP(11~28)的聚集行为。为了验证这一猜想,将His18分别突变成丝氨酸(Ser)和精氨酸(Arg),将这个2个突变后的样品hIAPP(H18S)和hIAPP(H18R)在37 ℃下培养3 d,发现组氨酸被丝氨酸替换后出现了图 5A所示的短棒状聚集,其长度大约是(0.28±0.02) μm,明显比hIAPP(11~28)形成的纤维(长度为(1.81±0.47) μm)要短,说明这种hIAPP(H18S)突变体聚集能力明显下降。当组氨酸被精氨酸替换后,只观察到颗粒状和块状的聚集体,而且数量较少(图 5B)。以上实验说明,His18在hIAPP(11~28)的聚集中起到很大的作用。当其与铜离子结合后很可能改变了局域的分子结构,从而抑制了多肽的聚集。
 图5
hIAPP(H18S)(A)和hIAPP(H18R)(B)氨基酸突变后的肽片段在37 ℃环境下培养3 d后的AFM图像及纤维长度统计图(C)
Figure5.
AFM images of hIAPP(H18S)(A)和hIAPP(H18R)(B) peptide fragments incubated at 37 ℃ for three days, and the statistical diagram(C) of lengths for 3 peptide fragments
图5
hIAPP(H18S)(A)和hIAPP(H18R)(B)氨基酸突变后的肽片段在37 ℃环境下培养3 d后的AFM图像及纤维长度统计图(C)
Figure5.
AFM images of hIAPP(H18S)(A)和hIAPP(H18R)(B) peptide fragments incubated at 37 ℃ for three days, and the statistical diagram(C) of lengths for 3 peptide fragments
 图6
hIAPP(H18S)(A)和hIAPP(H18R)(B)氨基酸突变后的肽片段在37 ℃环境下与50 μmol/L氯化铜混合培养3 d后的AFM形貌
Figure6.
AFM images of hIAPP(H18S)(A) and hIAPP(H18R)(B) peptide fragments incubated at 37 ℃ for three days in the presence of 50 μmol/L CuCl2
图6
hIAPP(H18S)(A)和hIAPP(H18R)(B)氨基酸突变后的肽片段在37 ℃环境下与50 μmol/L氯化铜混合培养3 d后的AFM形貌
Figure6.
AFM images of hIAPP(H18S)(A) and hIAPP(H18R)(B) peptide fragments incubated at 37 ℃ for three days in the presence of 50 μmol/L CuCl2
3 结论
人胰岛淀粉样多肽(hIAPP)是和Ⅱ型糖尿病密切相关的一类淀粉样多肽,而金属离子对淀粉样多肽的聚集有很大的影响。初步研究了铜离子和hIAPP的相互作用,原子力显微镜和硫代黄素(ThT)荧光的结果表明铜离子能够抑制多肽的聚集,其抑制程度和铜离子的浓度密切相关,而且,铜离子能够明显改变hIAPP多肽的聚集途径以及聚集体的二级结构。铜离子能够抑制hIAPP(11~28)多肽的聚集很可能是与组氨酸(His18)结合,改变了多肽的局域结构。通过这个研究将有助于更加深入地了解铜离子与hIAPP(11~28)的相互作用,并对Ⅱ型糖尿病的起因有新的理解。
-
-
[1]
Ashraf G M, Greig N H, Khan T A. Protein Misfolding and Aggregation in Alzheimer's Disease and Type 2 Diabetes Mellitus[J]. CNS Neurol Disord-Dr, 2014, 13(7): 1280-1293. doi: 10.2174/1871527313666140917095514
-
[2]
Detoma A S, Salamekh S, Ramamoorthy A. Misfolded Proteins in Alzheimer's Disease and Type Ⅱ Diabetes[J]. Chem Soc Rev, 2011, 41(2): 608-621.
-
[3]
王承克, 王振新. Cu(Ⅱ)离子诱导的β-淀粉样蛋白聚集抑制剂筛选及分子构效关系分析[J]. 应用化学, 2016,33,(7): 834-840. doi: 10.11944/j.issn.1000-0518.2016.07.150375WANG Chengke, WANG Zhenxin. Screening of Cu(Ⅱ) Ion Induced β-Amyloid Peptide Aggregation Inhibitor and Their Molecular Structure Analysis[J]. Chinese J Appl Chem, 2016, 33(7): 834-840. doi: 10.11944/j.issn.1000-0518.2016.07.150375
-
[4]
吕小平, 谭相石. 阿尔茨海默病相关的金属内稳态平衡调控研究[J]. 化学进展, 2013,25,(4): 511-519. LV Xiaoping, TAN Xiangshi. Metals Homeostasis and Related Proteins in Alzheimer's Disease[J]. Prog Chem, 2013, 25(4): 511-519.
-
[5]
Association A D. American Diabetes Association. Diagnosis and Classification of Diabetes Mellitus[J]. Diabetes Care, 2010, 33(Suppl 1): s62-s69.
-
[6]
Chimon S, Shaibat M A, Jones C R. Evidence of Fibril-Like Beta-Sheet Structures in a Neurotoxic Amyloid Intermediate of Alzheimer's Beta-Amyloid[J]. Nat Struct Mol Biol, 2007, 14: 1157-1164. doi: 10.1038/nsmb1345
-
[7]
Hardy J, Selkoe D J. The Amyloid Hypothesis of Alzheimer's Disease:Progress and Problems on the Road to Therapeutics[J]. Science, 2002, 297(5580): 353-356. doi: 10.1126/science.1072994
-
[8]
Konarkowska B, Aitken J F, Kistler J. The Aggregation Potential of Human Amylin Determines Its Cytotoxicity Towards Islet β-Cells[J]. FEBS J, 2006, 273(15): 3614-3624. doi: 10.1111/ejb.2006.273.issue-15
-
[9]
Cooper G J, Willis A C, Clark A. Purification and Characterization of a Peptide from Amyloid-Rich Pancreases of Type 2 Diabetic Patients[J]. Proc Natl Acad Sci, 1987, 84(23): 8628-8632. doi: 10.1073/pnas.84.23.8628
-
[10]
Zraika S, Hull R L, Udayasankar J. Oxidative Stress is Induced by Islet Amyloid Formation and Time-Dependently Mediates Amyloid-Induced Beta Cell Apoptosis[J]. Diabetologia, 2009, 52(4): 626-635. doi: 10.1007/s00125-008-1255-x
-
[11]
张勇, 竹俊苗, 刘长林. 金属离子Cu2+和Zn2+诱导β-淀粉样蛋白的聚集[J]. 药学学报, 2012,47,(3): 399-404. ZHANG Yong, ZHU Junmiao, LIU Changlin. Cu2+ and Zn2+-induced Aggregation of Amyloid-β Peptide[J]. Acta Pharm Sin, 2012, 47(3): 399-404.
-
[12]
Malisauskas M, Zamotin V, Jass J. Amyloid Protofilaments from the Calcium-binding Protein Equine Lysozyme:Formation of Ring and Linear Structures Depends on pH and Metal Ion Concentration[J]. J Mol Biol, 2003, 330(4): 879-890. doi: 10.1016/S0022-2836(03)00551-5
-
[13]
崔成毅, 张金海, 周星飞. 温度对胰高血糖素纤维生长的影响[J]. 核技术, 2008,31,(11): 877-880. doi: 10.3321/j.issn:0253-3219.2008.11.018CUI Chengyi, ZHANG Jinhai, ZHOU Xingfei. Temperature Effects on Growth of Glucagon Fibers[J]. Nucl Sci Tech, 2008, 31(11): 877-880. doi: 10.3321/j.issn:0253-3219.2008.11.018
-
[14]
Bush A I. The Metallobiology of Alzheimer's Disease[J]. Trends Neurosci, 2003, 26(4): 207-214. doi: 10.1016/S0166-2236(03)00067-5
-
[15]
Zargar A H, Bashir M I, Khan A R. Copper, Zinc and Magnesium Levels in Fibrocalculous Pancreatic Diabetes[J]. Exp Clin Endocrinol Diabetes, 2000, 108: 397-400. doi: 10.1055/s-2000-8135
-
[16]
Smith D, Ciccotosto G D, Fodero-Tavoletti M. Concentration Dependent Cu2+ Induced Aggregation and Dityrosine Formation of the Alzheimer's Disease Amyloid-beta Peptide[J]. Biochemistry, 2007, 46: 2881-2891. doi: 10.1021/bi0620961
-
[17]
Masad A, Hayes L, Tabner B J. Copper-mediated Formation of Hydrogen Peroxide from the Amylin Peptide:A Novel Mechanism for Degeneration of Islet Cells in Type-2 Diabetes Mellitus?[J]. FEBS Lett, 2007, 581(18): 3489-3493. doi: 10.1016/j.febslet.2007.06.061
-
[18]
Magrì A, La M D, Nicoletti V G. New Insight in Copper-Ion Binding to Human Islet Amyloid:The Contribution of Metal-Complex Speciation to Reveal the Polypeptide Toxicity[J]. Chemistry, 2016, 22(37): 13287-13300. doi: 10.1002/chem.201602816
-
[19]
Sinopoli A, Magrì A, Milardi D. The Role of Copper(Ⅱ) in the Aggregation of Human Amylin[J]. Metallomics, 2014, 6(10): 1841-1852. doi: 10.1039/C4MT00130C
-
[20]
黎虹颖, 古宁宇, 唐纪琳. 基于原子力显微镜的单分子力谱在生物研究中的应用[J]. 应用化学, 2012,29,(12): 1356-1363. LI Hongying, GU Ningyu, TANG Jilin. Application of Atomic Force Microscopy Based Single Molecule Force Spectroscopy in Biological Research[J]. Chinese J Appl Chem, 2012, 29(12): 1356-1363.
-
[21]
Haris P I, Chapman D. Analysis of Polypeptide and Protein Structures Using Fourier Transform Infrared Spectroscopy[J]. Methods Mol Biol, 1994, 22: 183-202.
-
[22]
董炎明, 王勉, 吴玉松. 用FTIR测定邻苯二甲酰化壳聚糖的取代度[J]. 应用化学, 2001,18,(4): 259-263. DONG Yanming, WANG Mian, WU Yusong. Determination of Degree of Substitution for Phthaloyl Chitosan by FTIR[J]. Chinese J Appl Chem, 2001, 18(4): 259-263.
-
[23]
Qi R, Luo Y, Ma B. Conformational Distribution and α-Helix to β-Sheet Transition of Human Amylin Fragment Dimer[J]. Biomacromolecules, 2014, 15(1): 122-131. doi: 10.1021/bm401406e
-
[24]
Robbins K J, Liu G, Selmani V. Conformational Analysis of Thioflavin T Bound to the Surface of Amyloid Fibrils[J]. Langmuir, 2012, 28(48): 16490-16495. doi: 10.1021/la303677t
-
[25]
Khurana R, Coleman C, Ionescu-Zanetti C. Mechanism of Thioflavin T Binding to Amyloid Fibrils[J]. J Struct Biol, 2005, 151(3): 229-238. doi: 10.1016/j.jsb.2005.06.006
-
[26]
Faller P, Hureau C, Berthoumieu O. Role of Metal Ions in the Self-assembly of the Alzheimer's Amyloid-β Peptide[J]. Inorg Chem, 2013, 52: 12193-12206. doi: 10.1021/ic4003059
-
[27]
Suzuki K, Miura T, Takeuchi H. Inhibitory Effect of Copper(Ⅱ) on Zinc(Ⅱ)-induced Aggregation of Amyloid-beta Peptide[J]. Biochem Biophys Res Commun, 2001, 285(4): 991-996. doi: 10.1006/bbrc.2001.5263
-
[28]
Goormaghtigh E, Cabiaux V, Ruysschaert J M. Determination of Soluble and Membrane Protein Structure by Fourier Transform Infrared Spectroscopy[J]. Subcell Biochem, 1994, 23: 329-362. doi: 10.1007/978-1-4615-1863-1
-
[29]
Kong J, Yu S. Fourier Transform Infrared Spectroscopic Analysis of Protein Secondary Structures[J]. Acta Biochim Biophys Sin, 2007, 39(8): 549-559. doi: 10.1111/abbs.2007.39.issue-8
-
[30]
Sánchezlópez C, Cortésmejía R, Miotto M C. Copper Coordination Features of Human Islet Amyloid Polypeptide:The Type 2 Diabetes Peptide[J]. Inorg Chem, 2016, 55: 10727-10740. doi: 10.1021/acs.inorgchem.6b01963
-
[31]
Rivillasacevedo L, Amero C, Quintanar L. Structural Basis for the Inhibition of Truncated Islet Amyloid Polypeptide Aggregation by Cu(Ⅱ):Insights into the Bioinorganic Chemistry of Type Ⅱ Diabetes[J]. Inorg Chem, 2015, 54(8): 3788-3796. doi: 10.1021/ic502945k
-
[1]
-

图 1 4种不同序列的肽片段在37 ℃环境下培养3 d后的AFM图像
Figure 1 AFM images of four peptide fragments incubated at 37 ℃ for three days
A.hIAPP(1~37); B.hIAPP(11~28); C.hIAPP(1~11); D.hIAPP(28~37)

图 2 铜离子对hIAPP(11~28)纤维化的影响。A~C是hIAPP (11~28)分别与不同浓度的氯化铜混合培养3 d后的AFM形貌及纤维长度统计图
Figure 2 AFM images of hIAPP(11~28) incubated at 37 ℃ for three days in the presence of copper ions at different concentrations(A~C), and the statistical diagram(D) of lengths of hIAPP(11~28) aggregation incubated with copper ions at different concentration
c(Cu2+)/(μmol·L-1):A.10; B. 50; C.100

图 3 ThT荧光光谱检测铜离子影响hIAPP(11~28)聚集过程
Figure 3 Kinetic analysis of hIAPP (11~28) fibril formation monitored by ThT fluorescence in the absence and presence of different concentration Cu2+
c(Cu2+)/(μmol·L-1):a.100; b.50; c.10; d.0

图 4 hIAPP(11~28)在加入不同浓度的铜离子后在37 ℃下培养3 d后的Micro-FTIR图
Figure 4 Micro-FTIR spectra of hIAPP(11~28) incubated at 37 ℃ for three days in the absence or in the presence of Cu2+ ions at different concentrations
c(Cu2+)/(μmol·L-1):a.100; b.10; c.0

图 5 hIAPP(H18S)(A)和hIAPP(H18R)(B)氨基酸突变后的肽片段在37 ℃环境下培养3 d后的AFM图像及纤维长度统计图(C)
Figure 5 AFM images of hIAPP(H18S)(A)和hIAPP(H18R)(B) peptide fragments incubated at 37 ℃ for three days, and the statistical diagram(C) of lengths for 3 peptide fragments
-


 下载:
下载:





 扫一扫看文章
扫一扫看文章
计量
- PDF下载量: 7
- 文章访问数: 2744
- HTML全文浏览量: 1094
 下载:
下载:
