Table 1.
Impacts of micro(nano)plastics on methane production performance and microbial community variation in AD system.
Citation:
Weixin Zhao, Shuocheng Shao, Hao Ma, Jianyuan Zhen, Shufei He, Chuandong Wu, Liangliang Wei. Deciphering the potential mechanisms and influencing factors of the effects of micro(nano)plastics on microbe in sludge anaerobic digestion system[J]. Chinese Chemical Letters,
2025, 36(11): 110846.
doi:
10.1016/j.cclet.2025.110846

Deciphering the potential mechanisms and influencing factors of the effects of micro(nano)plastics on microbe in sludge anaerobic digestion system
English
Deciphering the potential mechanisms and influencing factors of the effects of micro(nano)plastics on microbe in sludge anaerobic digestion system
Abstract:
Approximately 99% of micro(nano)plastics in wastewater are retained in waste activated sludge, inhibiting anaerobic digestion, while their specific effects on functional microbes remain unclear. To break through the limitations of current knowledge, in this review, we focused on summarizing the impacts of micro(nano)plastics on the microbial communities within anaerobic digestion systems, analyzing the toxicity mechanisms and developing strategies to mitigate their inhibitory effects. Firstly, the impacts of micro(nano)plastics on methane production and functional microbes were summarized, including direct cell pitting effects, inhibition caused by toxic leachates, and the adsorption of pollutants onto micro(nano)plastics surfaces, which further interfere with microbial activity and metabolic processes. Then, the specific performances and potential mechanisms by which micro(nano)plastics affect microbes were innovatively analyzed from the aspects of community variation, cellular activity and genetic expression. Moreover, various factors of micro(nano)plastics were found to influence their effects on microbes, including hormesis-like effects at different dosages, increased toxicity with decreasing particle size, enhanced biotoxicity due to surface functional groups, and variations in toxicity based on morphology and aggregation state. Furthermore, potential mitigation strategies, including activated carbon addition, thermal hydrolysis and cationic polyacrylamide application, were firstly summarized in here to alleviate inhibition on microbe. Finally, the current challenges and future directions were fully discussed and prospected. These insights could not only elucidate the biotoxic effects of micro(nano)plastics, but also provide a new avenue for helping to develop effective remediation techniques in micro(nano)plastic pollution management.
-
Key words:
- Micro(nano)plastics
- / Mitigation strategy
- / Biotoxicity
- / Microbial inhibition
- / Anaerobic digestion
-
1. Introduction
Plastic products, widely used in production and daily life due to their lightweight, durability, and low cost, have seen global production rise from 1.4 million tons in 1950 to 360 million tons in 2018, with an expected increase to 1.8 billion tons by 2050 [1,2]. Due to the massive use and improper disposal of plastics, large amounts of micro- or nanoplastic particles are inevitably generated [3]. Various and huge amounts of micro(nano)plastics have been detected in the various natural and artificial ecosystems, including oceans, rivers, soils and wastewater treatment plants (WWTPs) [4]. Micro(nano)plastics might pose severe ecological risks due to their harmful effects of tiny particle size, adsorbed contaminants, and/or toxic additives [5]. For example, micro(nano)plastics may exert toxic effects on aquatic organisms, including algae, which form the base of the food chain, and can be transferred through the food chain, ultimately disrupting the balance of aquatic ecosystems and impacting human health [6]. The ubiquitous characteristics and huge hazardous effects of micro(nano)plastics have raised increasing public concern.
Notably, large amounts of micro(nano)plastics generated from manufacturing and daily life are discharged into wastewater, and finally collected into the WWTPs via the urban drainage system [7,8]. It has been found that more than 99% of these micro(nano)plastics are finally retained or transferred to the waste activated sludge (WAS) during the wastewater treatment process [9]. The micro(nano)plastics accumulated in WAS were reported to composed of polyvinyl chloride (41.18%), polybutylene (23.53%), polytetrafluoroethylene (11.76%), polyethylene (11.76%) and polyacrylonitrile (5.88%) [10]. In terms of morphology, micro(nano)plastics in WAS were primarily composed of fibers, accounting for 63%, followed by shafts at 15%, films at 14%, flakes at 7.3%, and spheres at 1.3% [11]. The abundances of micro(nano)plastics accumulated in sludge were reported to range from 1.6 × 103 particles/kg to 5.6 × 104 particles/kg dry weight, indicating that WAS may act as an important long-term sink for micro(nano)plastics [12,13]. The high-level abundance of micro(nano)plastics in WAS undoubtedly has a serious impact on the subsequent sludge treatment and disposal processes, which deserve close attention.
Anaerobic digestion (AD) is considered to be the most widely used sludge treatment process around the world due to its dual advantages of organic disposal and energy recovery [14,15]. Moreover, AD is a multifaceted and complex biochemical process relying on the interaction of a diverse and syntrophic microbiota [16]. Through the interactions of various functional microbes (e.g., acidogens, acetogens, and methanogens), organic matters were transformed into methane [17]. However, the operation efficiency and system stability of anaerobic ecosystem are fragile and susceptible to exogenous biotoxins, resulting in decreased methane production efficiency and a longer system start-up period [12,18]. As a type of emerging pollutants, micro(nano)plastics are known for their biological toxicity, which can negatively impact the functional microbes and disturb the operation of digestion system [5].
To date, negative impacts of micro(nano)plastics on the methane production performance and operation stability of AD have been widely reported in the previous studies [19-21]. The presence of micro(nano)plastics not only decreases the methane yield, but also prolongs the lag phase [22]. Furthermore, owing to the large specific surface area and strong hydrophobicity, micro(nano)plastics act as potential carriers of heavy metals, antibiotics and other pollutants, thus inducing greater toxicity in the AD of WAS [4]. For example, the simultaneous presence of micro(nano)plastics and organic contaminants leads to compounded effects, such as heightened toxicity, (bio)accumulation, physical damage, shifts in microbial populations, and disturbances to ecosystems [23]. Despite growing awareness of the negative influences of micro(nano)plastics on AD, significant research gaps remain. Previous studies mostly focused on the presence and impacts of micro(nano)plastics in macroscopic digestion performance, with less attention given to their microbial aspects, particularly in community succession, cell growth, and gene expression. Actually, micro(nano)plastics and their leachates could affect functional genes expression, influence the activities of key enzymes, and change microbial community composition, which further disturb the organics transformation and carbon cycling [24]. Besides, systematically summarizing and analyzing the current research about how to alleviate the adverse impacts of micro(nano)plastics on AD is also rare. Additionally, the understanding of the fate, degradation, and transport of micro(nano)plastics within digestion systems remains limited, highlighting the need for further investigation in this area.
Thus, the purpose of this study is to comprehensively summarize and analyze the impacts and mechanism of micro(nano)plastics on the functional microbes in the AD process of WAS. Firstly, the comprehensive overview of the negative effects of micro(nano)plastics on microorganisms was firstly elucidated. Then, the specific performances and mechanisms by which micro(nano)plastics influence microbial behaviors were analyzed. Moreover, the impacts of various characteristics of micro(nano)plastics, including dosage, particle size, surface functional groups and shape, on microbial activities were systematically analyzed. Furthermore, several potential strategies aimed at mitigating the inhibition effects of micro(nano)plastics on microbes were discussed. Finally, future perspectives and potential directions were recommended. In all, this study will offer valuable insights into a thorough comprehension of how micro(nano)plastics affect microbial activity in AD systems, which will be helpful to developing effective strategies to prevent hazards caused by micro(nano)plastics.
2. Overview of the effects of micro(nano)plastics on sludge digestion system
2.1 Impacts of microplastics on methane production
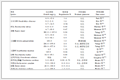
As shown in Table 1, it can be summarized that most previous studies have documented an inhibitory impact of micro(nano)plastics on methane production processes. Although several studies have observed that micro(nano)plastics may increase the abundance of hydrolytic acidogenic bacteria (such as Acidobacteriota and Longilinea) and promote the hydrolysis of soluble polysaccharides and proteins, while the abundance of methanogenic archaea were greatly reduced, thereby inhibiting the methanogenesis phase and ultimately leading to a decrease in the methane production yield. Especially, Methanosaeta, as an obligate acetoclastic methanogen, is highly sensitive to environmental conditions and is easily inhibited by micro(nano)plastics [25]. Since acetoclastic methanogenesis is the primary pathway for methane production in anaerobic sludge digestion, the inhibition of Methanosaeta significantly reduces methane yield [10]. Besides, except for the inhibition on the cumulative yield, previous studies also reported that the micro(nano)plastics could prolong the lag phase and decrease maximum methane production rate [25,26]. It should be noted that the effects of micro(nano)plastics on methane production have shown considerable variation across different studies, likely due to factors such as the type, concentration, and particle size of the micro(nano)plastics. It should be pointed out that the majority of previous explorations on the influences of micro(nano)plastics on methane production in the AD of WAS were predominantly employed batch biochemical methane potential tests and conducted under mesophilic conditions. This approach corresponds with the widespread implementation of mesophilic AD for sewage sludge stabilization in wastewater treatment facilities. The is methodology likely aligns with the prevalent application of mesophilic AD for sludge stabilization in WWTPs. To comprehensively assess the impacts of microplastics, further research should expand to include long-term digestion experiments and explore a range of operational temperatures.
Table 1
 DownLoad:
CSV
DownLoad:
CSV
Micro(nano)plastics type Dosage Size Substrate Digestion mode Methane production performance Bacteria variation Methanogen variation Ref. Polystyrene 150 µg/L 50 nm WAS Batch, 37 ℃, 300 rpm Methane yield decreased by 29.34% Cloacamonaceae_W22 decreased by 14.6% Methanosaeta decreased by 16% [28] Polyvinyl Chloride 60 particles/g TS 1 mm WAS Batch, 37 ± 1 ℃ Methane yield decreased by 24.2% Proteiniborus decreased by 25% Methanosaeta decreased by 16.5% [57] Polyethylene 200 particles/g TS 40 ± 2 µm WAS Batch, 37 ± 1 ℃ Methane yield decreased by 27.5% Rhodobacter decreased by 15.2% Methanosaeta decreased by 16% [119] Polystyrene 160 particles/g TS 100 µm WAS Batch, 37 ℃, 150 rpm Methane yield decreased by 11.04% Leptolinea decreased by 1.99% Methanobacterium decreased by 5.31% [39] Polyvinyl chloride 2.4 g/g VS 3000 µm WAS Semi-continuous, OLR = 15 g VS/L/d Methane yield improved by 34.9% Acidobacteriota improved by 17.7% Methanosaeta decreased by 53.6% [52] Polystyrene 150 µg/L 50 nm WAS Batch, 37 ± 2 ℃, 300 rpm Methane yield decreased by 32.3% Treponema completely disappeared Methanosaeta decreased by 14% [91] Polystyrene 50 mg/g TS 50 nm WAS Batch, 35 ± 2 ℃ Methane yield decreased by 15.5% Sulfurovum decreased by 36.6% Candidatus Methanofastidiosum
decreased by 61%[22] Polyethylene 100 mg/g TS 180 µm WAS Batch, 37 ℃, 120 rpm Methane yield decreased by 13.8% Atopostipes increased by 2.1% Methanosaeta decreased by 3.4% [120] Polycarbonate 30 particles/g TS 40 µm WAS Batch, 37 ± 1 ℃, 300 rpm Methane yield improved by 24.7% Longilinea increased by 1.34% Methanosaeta decreased by 8.4% [35] Polyvinyl chloride 30 mg/g TS 0.35 mm WAS Batch, 35 ± 1 ℃, 140 rpm Methane yield decreased by 15.62% Actinobacteria decreased by 12% Methanosaeta decreased by 1.1% [64] 2.2 Negative effects of micro(nano)plastics on microbes
2.2.1 Cell pitting induced by direct physical contact
As displayed in Fig. 1, micro(nano)plastics can disrupt microbial cells through direct physical attachment and penetration of the biolipid layer on the cell walls, a process known as cell pitting, which not only induces membrane permeation and cell rupture, but also impacts the synergistic relationships within microbial communities, consequently reducing methane production during the AD process [27,28]. Micro(nano)plastics were observed adhere to the cell surface of hydrogenogenic Acetobacteroides hydrogenigenes via scanning electron microscopy, alongside the presence of numerous nano-sized pores on the membrane owing to the intrusion of micro(nano)plastics particles [29]. The direct injury to microbes was primarily dependent on the size and morphology features of micro(nano)plastics [30]. Generally, more serious toxic effects on microbes are induced by micro(nano)plastics with smaller sizes, which are more effective at penetrating cell walls and inducing more oxidative stress [31]. Morphological features, such as pointed edges or irregular shapes, can physically damage microbial cells upon contact [32]. Additionally, the granule size of the sludge trends to decrease with an increase in the concentration of micro(nano)plastics [33].
Figure 1
 Figure 1. Schematic diagram of the toxic effects of micro(nano)plastics on microbes. Modified with permission [27]. Copyright 2023, Elsevier.
Figure 1. Schematic diagram of the toxic effects of micro(nano)plastics on microbes. Modified with permission [27]. Copyright 2023, Elsevier.2.2.2 Toxic effects induced by leachates and polymeric monomers
Various chemical additives, including plasticizers, stabilizers, and flame retardants, are added during the manufacturing process to confer new characteristics and prolong the service life of plastic products [34]. However, due to weak hydrogen bonding and van der Waals forces, most additives are gradually released from the micro(nano)plastics under the effects of sunlight, wind, and physical friction [35]. The toxic chemical additives released were eventually accumulated in the WAS and detection levels ranged from mg/kg TS to more than one g/kg TS in WAS, which was reported to be highly related to the biotoxicity effect of microplastic [36,37]. It was reported that chemical additives can directly disrupt microbial cells, induce oxidative stress and impair microbial functions [32]. However, the generated reactive oxygen species (ROS) could accelerate micro(nano)plastics degradation and aging, thereby leading to more toxic additives being leached and ultimately resulting in a vicious circle [38]. The leached antioxidants, such as the 2, 2′-methylenebis (6-tert-butyl-4-methylphenol), can interact with amino acid components of enzymes, causing notable changes in enzyme structures and consequently suppressing enzyme functions [39]. Due to the persistence and bioaccumulation effects of plastics additives, the community structure was observed to eventually shift towards conditions less favorable for methanation under the long-term exposure [40,41]. It is notable that the effects of plastics additives on microbes are species-dependent, and different additives showed disparate effects on the functional microbes due to the diversity of species in the complex sludge AD systems [36]. Except for the chemical additives, polymeric monomers (e.g., caprolactam) might also be leached from micro(nano)plastics [42,43]. The polymeric monomers can occupy the active sites of functional enzymes, which inhibit the enzyme activity [42].
2.2.3 Adsorbing other pollutants from the environment
Owing to the high surface area, hydrophobic nature and charged groups, micro(nano)plastics can serve as vectors for other pollutants, such as heavy metals, antibiotics and polycyclic aromatics, via hydrophobic interactions, partitioning, electrostatic interactions, and other non-covalent interactions during the wastewater treatment process [10]. It should be noted that multiple adsorption effects occur simultaneously in general, and their relative contributions are affected by various factors physicochemical characteristics of micro(nano)plastics (e.g., particle size, crystallinity, and functional groups), and environmental properties (e.g., pH, temperature, and ionic strength), as well as the characteristics of co-pollutants (e.g., types, functional groups, concentration, and hydrophobicity) [44,45]. The adsorption and desorption process of micro(nano)plastics change the environmental behavior of these pollutants, which generally prolongs the presence of pollutants and affects their biotoxicity as well as bioavailability [46,47]. Specifically, co-pollutants might induce more severe toxic effects on microbes than the inhibition induced by sole micro(nano)plastics, including higher oxidative stress, lower enzyme activity, reduced metabolism, and even resulting in cell apoptosis [48,49]. Moreover, surface cracks and higher roughness were observed on the aged micro(nano)plastics, resulting in a greater capacity to adsorb pollutants [50]. Besides, it is notable that owing to the colonization of micro(nano)plastics on microorganisms, the adsorbed pollutants may more readily come into contact with microorganisms, potentially leading to more severe ecological damage [44].
3. Performances and mechanisms of micro(nano)plastics affecting microbial behaviors
3.1 Disturbed microbial diversity and more complex co-occurrence networks
Extensive research has explored the impacts of micro(nano)plastics on microbial composition and diversity in AD systems, and most of the previous results suggested that micro(nano)plastics could reduce the diversity and alter the abundance of the functional microbes within digesters [22,51]. The alpha diversity indices, including Shannon, Chao1, Ace, and PD tree indices, were observed to decrease under micro(nano)plastics exposure, indicating that the homogeneity and variety of functional microbes were decreased [39,52]. Besides, apparent clustering between sludge samples under similar micro(nano)plastics exposure was observed in the principal coordinates analysis plot, suggesting that significantly different beta diversity and micro(nano)plastics shifted the microbial community structure [53]. These alterations were attributed to the selective enrichment of specific bacterial on the surface of micro(nano)plastics [26].
As shown in Table 1, the presence of micro(nano)plastics obviously decreased the abundance of various hydrolytic-acidifying bacteria (e.g., Proteiniborus, Rhodobacter and Cloacamonaceae_W22) in most cases, which hindered the organics transfer from solid sludge to the aqueous phase. Besides, the abundance of aceticlastic methanogens (e.g., Methanosaeta) was greatly declined, which was in accordance with the decline in methane yield observed. The phenomenon might be due to the varying resistance of different microorganisms to microplastic, i.e., the hydrogenotrophic methanogens (e.g., Methanosarcina) with the ability to withstand harsh conditions were enriched, while the aceticlastic methanogens (e.g., Methanosaeta) declined due to their sensitivity to toxicity [52].
Notably, although the micro(nano)plastics decreased the community diversity and the relative abundance of the functional microbes, more nodes and higher average degree in the microbial co-occurrence networks were observed, indicating that micro(nano)plastics increased the complexity and modularity of digestion sludge microbiome networks [54,55]. And the normalized stochasticity values of the digested sludge were lower than 50%, indicating that the governing roles of micro(nano)plastics in determining the assembly processes in the community [54]. The tighter associations among the microbes were considered as a self-protection response to withstand the toxicity effects induced by micro(nano)plastics [54]. Moreover, new positive correlations between aceticlastic methanogens (e.g., Methanosaeta and Methanosarcina) and various hydrolytic-acidifying bacteria (e.g., Candidatus Competibacter, Rhodobacter, and Desulfobulbus) were established, which were helpful in assisting the energy and substrate transfer to defend against micro(nano)plastics stress [53].
3.2 Induce more extracellular polymeric substances secretion
Extracellular polymeric substances (EPS) are high-molecular-weight polymers secreted by microbes, serving vital functions in protecting the cell structure and crucially affecting the stability of AD systems [40]. The existence of micro(nano)plastics was firstly agglomerated on the outer layer of the EPS, combining with EPS through electrostatic adsorption [56]. The side chains of lipids, amino acids and functional groups (e.g., carbonyl and amine groups) in the EPS play an essential role in defensing against micro(nano)plastics toxicity, which was gradually consumed, accompanied with the observation of the decreased content of loosely bound EPS and the increasingly ruptured structure [39,57].
However, as shown in Fig. 2, the functional genes encoding EPS biosynthesis (e.g., Pel and Psl) were activated and improved, which resulted in more extracellular polymers to adapt to adverse conditions, as observed by the increased tightly bound EPS [39,58]. And the more secreted EPS can improve nutrient capture from the extracellular environment and promote cells to adhere to each other, thereby enhancing the ability to defend against microplastic stress [59,60]. And the newly secreted EPS might promote microbes to adsorb on the micro(nano)plastics surface in the form of a biofilm, which could provide a special ecological niche that is less affected by the external environment [38,61]. Meanwhile, the indices related to protein secondary structure, such as α-helix and β-sheet, were increased after exposure to micro(nano)plastics, indicating that the EPS structure became more compact to resist the toxicity [62]. However, the excessive micro(nano)plastics could penetrate EPS and transferred into inner layers of sludge, posing damage to the microbial cell membrane due to their nano-specific properties and strongly hypothesized abilities [62]. Besides, the overhigh micro(nano)plastics pressure could restrain the expression of genes responsible for EPS generation, and the high level of toxic leachates (e.g., sodium dodecyl sulfate) might dissolve EPS, finally resulting in low EPS content [33,48,59].
Figure 2

3.3 Decreased enzyme activity and increased oxidative stress
The activities of biochemical enzymes associated with the AD process (e.g., protease, α-glucoside, acetate kinase, and coenzyme F420) and antioxidant enzymes (e.g., superoxide dismutase, superoxide reductase, catalase, and peroxidase) were observed to significantly decrease under the stress of micro(nano)plastics and their leachates [63]. Specifically, multiple studies have confirmed that the acetate kinase, and coenzyme F420 were greatly inhibited by the micro(nano)plastics stress, contributing to the decreased methane yield observed under higher micro(nano)plastics exposure [31,39]. Besides, one of the common chemical leachates, sodium dodecyl sulfate, was observed to inhibit the activities of antioxidant enzymes (e.g., superoxide dismutase and catalase), which decreases the resistance to oxidative stress caused by micro(nano)plastics [59].
As shown in Fig. 3, the reductive groups on the surface of micro(nano)plastics can react with the residual sub-micromolar concentrations of oxygen in the AD system, leading to the generation of various ROS at nanomolar concentrations, including superoxide radicals (•O2−), hydrogen peroxide (H2O2), and hydroxyl radicals (•OH) [35,64]. It should be noted that once the ROS exceed the regulation capability of antioxidant enzymes, it may result in lysosomal damage, cell membrane disruption, metabolic dysregulation, and even cell death [31,48,65].
Figure 3
 Figure 3. Mechanism diagram of the effects of polycarbonate micro(nano)plastics on the enzyme activity and ROS in WAS anaerobic digestion. Modified with permission [35]. Copyright 2023, Elsevier.
Figure 3. Mechanism diagram of the effects of polycarbonate micro(nano)plastics on the enzyme activity and ROS in WAS anaerobic digestion. Modified with permission [35]. Copyright 2023, Elsevier.3.4 Reduced functional genes expression and altered metabolic pathway
The attached nanoparticle on the cell membrane could change the membrane permeability, enter the cell and further disrupt the metabolic pathway by inducing excessive oxidative stress, impeding DNA synthesis [29]. Specifically, the functional genes encoding energy metabolism, amino acid metabolism and carbohydrate metabolism, were observed to decrease under higher micro(nano)plastics exposure, suggesting that micro(nano)plastics inhibited microbial metabolism [39]. Moreover, genes omcB and mtrC, responsible for mediating electron transfer to extracellular electron acceptors, were reported to decrease under micro(nano)plastics exposure, suggesting that micro(nano)plastics inhibited the extracellular electron transfer process [33,66]. Genes FTHFS, accA, accC and accD, encoding the biosynthesis of formyltetrahydrofolate in the acetyl-CoA pathway of homoacetogens, were observed to be inhibited, indicating that micro(nano)plastics could affect the acetogenesis pathways, accounting for the lower volatile fatty acid yield [67]. Except for affecting the genes involved in acidogenesis and homo-acetogenesis process, the existence of micro(nano)plastics also disrupted the expression of genes encoding methanogenic pathways. For example, the counts of methanogen-specific gene mcrA, were also reported to decrease [68]. Besides, previous studies also reported that the micro(nano)plastics reduced the number of functional genes associated with the nitrogen, phosphorus, and sulfur cycles. Specifically, 50 nm polystyrene nanoplastics obviously decreased the counts of genes lig, naglu, xylA, gcd, and phnk, which encoding lignin peroxidase, α-N-acetylglucosaminidase, xylose isomerase, quinoprotein glucose dehydrogenase, and putative phosphonate transport, respectively [22].
3.5 Promoted the horizontal transfer of antibiotic resistance genes
Antibiotic resistance genes (ARGs) are one of the emerging contaminants, which pose a significant threat to public health owing to their capability of horizontal gene transfer among different species [69]. The existence of micro(nano)plastics significantly increased the abundance of ARGs and mobile genetic elements, indicating that the existence of micro(nano)plastics could act as a reservoir to promote the dissemination of antibiotic resistance in the AD of WAS process [55]. Besides, micro(nano)plastics could specifically enrich the specific ARGs in the digester. Previous studies observed that the abundances of most ARGs type improved in the presence of micro(nano)plastics, while sulfonamide-resistant gene slightly decreased [63]. The reason for this inconsistent impact is complicated, possibly due to the complexity of antibiotic resistance bacteria community in different digester [63]. Each type of ARGs have its specific host microbes, and the ARGs abundance is closely related to the bacterial community, while the relationship between host bacterial community and ARGs abundance variation still unknown and needs further investigation.
The hydrophobic surface of micro(nano)plastics provided a conducive environment for biofilm development and plastisphere formation, which could selectively enrich potential hosts of antibiotic resistant bacteria, thereby promoting the intensive vertical and horizontal transfer of ARGs [38,55]. For example, the ARGs host bacteria Thermoanaerobacter and Caldicoprobacter were enriched by 5.20% and 28.03%, respectively, under the exposure of 200 particles/g-TS polyethylene microplastics [55]. Besides, micro(nano)plastics stress may induce hermetic effects as a microbial self-protection response to defend against and adapt to external stress, which may lead to the accelerated bacterial growth and proliferation [38]. And the improved bacterial proliferation could further promote ARGs vertical transfer [70]. Besides, the direct contact with micro(nano)plastics and the constant chemical additives released could induce severe oxidative stress and increased cell membrane permeability, which facilitated the ARGs to freely entering the cell and promote the horizontal gene flow processes, finally resulting in the acquisition and dissemination of ARGs between different bacteria within the plastisphere [38,71]. For instance, compared with the control, the counts of genes related to cell membrane permeability (e.g., YajC, and Mdla) increased to 1.13-folds in the digester with polyvinyl chloride addition, and the abundance of sulfonamide, beta-lactam, and tetracycline resistance genes also improved by 23.6% [63]. Moreover, the presence of micro(nano)plastics could improve EPS secretion, and the higher EPS content was conducive to genes retention and cell cohesion, thereby promoting the ARGs horizontal transfer [38]. Besides, as displayed in Fig. 4, the type Ⅳ secretion system, responsible for mediating DNA release into the milieu extracellular environment, was up-regulated under the exposure of micro(nano)plastics, which enhanced the active release of extracellular ARGs into the sludge AD system, accounting for the improved ARGs horizontal transfer process [72].
Figure 4
 Figure 4. Potential mechanisms of the enhanced horizontal flow of ARGs induced by micro(nano)plastics in anaerobic sludge digestion. Modified with permission [72]. Copyright 2022, Elsevier.
Figure 4. Potential mechanisms of the enhanced horizontal flow of ARGs induced by micro(nano)plastics in anaerobic sludge digestion. Modified with permission [72]. Copyright 2022, Elsevier.4. Uncovering the effects of micro(nano)plastics characteristics on microbes
4.1 Hormesis-like effects on microbial activities
Micro(nano)plastics exhibited dose-dependent hormesis-like effects on microbial activities in AD system, i.e., key functional enzyme activities were improved under low micro(nano)plastics exposure, while they were greatly inhibited after exposing to excess micro(nano)plastics [35,73]. The dosage-dependent effects of micro(nano)plastics were attributed to their leachates released from plastics, such as bisphenol A, di-N-butyl phthalate. Due to that the weak bond between chemical additives and plastics, the chemical additives might be easily released and displayed as the positive correlation with the increased concentration [74,75].
Although ROS were increased under the exposure of micro(nano)plastics, low concentrations of micro(nano)plastics and their leachates could stimulate the activities of key enzymes to resist oxidative stress. And the low oxidative stress levels improved biomass activity to enhance the methane production [39]. However, a high dosage of micro(nano)plastics and their leachates induced severe intracellular ROS and aggravated oxidative stress, which inhibited enzyme activity, and methanogenesis metabolic reaction [35,76].
Besides, similar effects were also observed for the polymeric monomers (e.g., caprolactam) released from micro(nano)plastics [42,43]. Previous studies also found that low-concentration caprolactam can serve as an activator, altering the active sites of the enzyme, thereby augmenting their affinity for the substrate and the catalytic activity [77]. Moreover, caprolactam as an activator binds to the substrate before associating with the enzyme molecule, thereby modifying the electron density or molecular tension of the substrate, facilitating product formation and enhancing enzyme catalysis [78,79]. However, the elevated caprolactam concentrations might be detrimental to enzymes responsible for hydrolysis, acidogenesis and methanogenesis [42]. Moreover, high concentrations of caprolactam may occupy the active sites of functional enzymes, consequently diminishing the promotive effect [42]. Besides, a strong positive linear correlation was observed between the ARGs abundance and micro(nano)plastics concentration, highlighting the essential role of micro(nano)plastics dosage in promoting ARGs dissemination [55].
4.2 Increased toxicity with decreasing particle size
The particle size of micro(nano)plastics was defined as ranging from micro to nanometer scales, which directly affects their potential contact area with microbes and influences the microbial activity [31]. Most of the previous studies reported that the inhibition effects of micro(nano)plastics were size-depended, i.e., the lower the micro(nano)plastics size, the higher the inhibition effects [39]. The phenomenon might be attributed to the fact that the smaller-sized micro(nano)plastics exhibited higher specific surface area, which exposed more active groups and facilitated the generating of ROS [67]. Except for the damage induced by direct contact with micro(nano)plastics, micro(nano)plastics are more likely to enter into the microbial cell by endocytosis and internalization, which could produce much higher reactive oxygen species and trigger the activation of signaling modules responsible for inflammation and apoptosis [31,52,67]. Moreover, the smaller-sized micro(nano)plastics possess larger surface areas and more adsorption sites, enabling them to adsorb more toxins and exhibit greater inhibition on microbes [80]. Besides, it should be pointed out that the micro(nano)plastics of smaller size are less stable than larger particles, which easily agglomerate into aggregates, thus reducing their adsorption capacity on other pollutants [81]. Compared with the smaller micro(nano)plastics, larger particles displayed stronger dispersion characteristics, which increased their contact with the sludge, ultimately leading to a decrease in biological activity [82]. However, no statistically significant difference in inhibition effects on the microbial activities was observed under the exposure of micro(nano)plastics with a larger size (100–1000 µm), while the corresponding high specific surface area was helpful to microbial colonization and promoted mass transfer [82-84]. Although less toxic effects were observed on the micro(nano)plastics with a large size, it should be noted that they may undergo mechanical forces such as mixing, impaction, and compression within these systems, potentially resulting in their disintegration into smaller fragments [82].
4.3 More severe biotoxicity induced by surface functional groups
The micro(nano)plastics undergo various physicochemical and biological actions, such as ultraviolet radiation, mechanical abrasion and microbial decomposition, which leads to the formation of differentially charged groups generated on the surface of micro(nano)plastics [32,85]. For example, surface oxidation and acquisition of functionalities during the weathering can endow micro(nano)plastics with carbonyl groups, while hydrolyzation of polymer may introduce an amino modification [9]. And X-ray photoelectron spectroscopy has found that functionalized micro(nano)plastics greatly disrupted the side chains of lipids or amino acids in EPS, indicating that the functionalized micro(nano)plastics penetrate the EPS matrix and affect microbes via pore tunnels of sludge [62]. And the increased abundances of aggregated strands, β-sheets and α-helices were observed in the EPS, indicating that the tight structure of EPS acts a self-protection response to functionalized micro(nano)plastics stress [62]. Moreover, except for the inhibition on individual microbes, the microbial community composition was also significantly altered. The abundance of several syntrophic bacteria (e.g., Syntrophobacter, Saccharofermentans, Treponema, Lactivibrio, and Paludibacter) and methanogen (e.g., Methanothrix and Methanomassiliicoccus) was greatly decreased [62]. Correspondingly, the counts of functional genes mcrA and ACA, which encode key enzymes in methanogenesis, were also decreased under the exposure of functionalized micro(nano)plastics [62].
4.4 Varying toxicity with morphology characteristics and aggregation status
Micro(nano)plastics displayed various morphology characteristics and aggregation states, which could affect their toxicity by altering their physical contact with microbes and adsorption with other pollutants [86]. The characteristics of micro(nano)plastics shapes greatly affect their surface roughness degrees [86]. And the increased effective surface area of the micro(nano)plastics was observed with the increased surface roughness and the appearance of cracks, which potentially strengthening their interactions with biological cells [87]. Moreover, the increased surface roughness of micro(nano)plastics was beneficial to improve the pollutants adsorption capacity, which could improve toxicity [88]. Notably, the expanding cracks in micro(nano)plastics might accelerate fragmentation and shredding via increasing the environmental contact surface with other physicochemical factors and accelerating their aging [86]. Besides, compared with the separate micro(nano)plastics, the presence of aggregates might affect the surface area and influencing the micro(nano)plastics aging, such as decelerate internal micro(nano)plastics aging but accelerate surface micro(nano)plastics aging, which might promote the adsorption and transport of pollutants [89,90]. Besides, it should be pointed out that evaluating the toxicity associated with specific shapes or types of micro(nano)plastics is challenging due to the limited availability of data and the difficulties in maintaining consistency across other variables.
5. Strategies for mitigating inhibition effects of micro(nano)plastics on microbes
5.1 Activated carbon addition
Adding activated carbon has been proved to offset the inhibition effects of micro(nano)plastics on sludge anaerobic digestion and further improve methane production [25]. In contrast to the decreased microbial microbial richness and diversity observed in digesters exposed to micro(nano)plastics, higher phylogenetic distance, Shannon index, and Chao1 indices were observed in the digester fed with granular activated carbon, suggesting that granular activated carbon amendment could drive a more diverse microbial community [25]. Moreover, granular activated carbon amendment significantly enriched functional microbes and changed the community composition involved in the AD system. Specifically, the abundances of several electroactive fermentation bacteria, including Clostridium and Bacteroides, were observed to increase, indicating the potential role of granular activated carbon in facilitating direct interspecies electron transfer [25]. Correspondingly, the relative abundance of DIET-active methanogens, including Methanosaeta and Methanobacterium, was increased in the granular activated carbon-amended reactors compared with that in the control [25]. Besides, granular activated carbon was observed to obviously decrease the abundance of ARGs (e.g., tetW, tetA, tetB, tetM, sul1, sul2, blaTEM, ermC) and mobile genetic elements (intl1 and intl2), indicating the great potential of granular activated carbon in reducing horizontal gene transfer of ARGs [25].
The above phenomenon might be attributed to the fact that micro(nano)plastics were adsorbed on the internal pores of granular activated carbon, which minimized the exposure of microbes to them and alleviated the toxic effects [25]. And the divalent ions (e.g., Ca2+, Mg2+) existing in the WAS is expected to generate additional adsorption sites on the granular activated carbon via bridging effects, resulting in a higher adsorption efficiency of micro(nano)plastics and the leached harmful agents (e.g., sodium dodecyl sulfate), and eliminating the oxidative stress on microbes [91]. The reduction in ARGs abundances after granular activated carbon addition might be attributed to its capacity to enhance bacterial activity and limit microbial movement by offering abundant sites for adherence [92]. Moreover, the adsorption of ARGs on the surface and pores of granular activated carbon could decrease the opportunity for ARGs to evolve and proliferate on micro(nano)plastics [25].
5.2 Thermal hydrolysis pretreatment
Thermal hydrolysis is a widely implemented sludge pretreatment method in full-scale anaerobic digestion facilities, offering significant benefits such as enhanced biogas recovery through improved hydrolysis, efficient removal of solids and pathogens, and enhanced dewatering capability of biosolids [93,94]. Recent study reported that thermal hydrolysis assisted AD could partially counteract the inhibition effects of micro(nano)plastics, relieve oxidative stress on microbes, and reduce ARGs propagation [28,95]. Despite the biodegradation resilience of plastic, the plastic polymer becomes more susceptible to hydrolytic degradation through the cleavage of ester linkages under the high-temperature of thermal hydrolysis (e.g., 160 ℃), which is more easily further degraded in subsequent AD process [96]. Meanwhile, cross-sectional layers and increased hydrophilicity were observed on the surface of micro(nano)plastics after thermal hydrolysis pretreatment, suggesting that micro(nano)plastics were more susceptible to microbial degradation [30,96]. Moreover, the relative crystallinity, hydrophilicity, and carbonyl index of micro(nano)plastics significantly declined after thermal hydrolysis, indicating that the adsorption to other pollutants was greatly decreased [96].
Except for promoting micro(nano)plastics degradation, thermal hydrolysis could also promote the microbial proliferation. Thermal hydrolysis enhanced the breakdown of large particulate organic matter, providing more available soluble organic matter for microbial utilization [97,98]. Higher Chao1, OTUs, and Shannon indices were observed in the digested system coupled with thermal hydrolysis pretreatment, indicating a more diverse digester microbiome [28]. And an increased abundance of acetoclastic Methanosaeta was observed after low-temperature thermal hydrolysis (e.g., 80 ℃), which was beneficial for the methane production. However, high-temperature thermal hydrolysis (e.g., 160 ℃) obviously decreased the abundance of Methanosaeta and Methanosarcina, which could be attributed to the formation of refractory and toxic compounds under high-temperature [28]. Besides, a considerable decrease in the number of ARGs (e.g., blaOXA, blaTEM, ermB and ermC) and integron (intl1 and intl2) was observed in the digester with thermal hydrolysis pretreatment, indicating that thermal hydrolysis could decrease the risks of ARG transmission through digestate biosolids [28,99].
5.3 Cationic polyacrylamide addition
Cationic polyacrylamide is a common flocculant, which has been reported to enmesh and capture the micro(nano)plastics during drinking water treatment [100]. Recent studies found that cationic polyacrylamide decreased the interaction of anaerobes and micro(nano)plastics via intensive ionic and van der Waals interactions with sludge and micro(nano)plastics, respectively [101,102]. The addition of cationic polyacrylamide improved the formation of sludge flocs, impeded the direct contact of micro(nano)plastics with oxygen, thereby affecting the ROS production and reducing the oxidative stress intensity [103]. Moreover, the existence of cationic polyacrylamide reduces the releasing of plastic additives and the degree of reduction raises with the increase of cationic polyacrylamide content. Thus, the addition of cationic polyacrylamide mitigated the negative effects of micro(nano)plastics on microbial community, and even increased the richness and diversity to a certain extent [64]. Correspondingly, the existence of cationic polyacrylamide restored the relative abundance of key acidogens (e.g., Longilinea and Christensenellaceae_R-7_group) and methanogens (e.g., Methanobacterium and Methanosaeta), accounting for the improved digestion performance.
6. Fate, degradation and transport of micro(nano)plastics in digestion system
The fate and transport of micro(nano)plastics in sludge AD system involve three main pathways: transfer, transformation, and removal. First, micro(nano)plastics are transferred from the liquid phase of wastewater to the solid phase of sludge through adsorption [104]. Second, some micro(nano)plastics undergo transformation during anaerobic digestion, particularly under extreme conditions such as high temperatures or alkaline environments. For example, certain biodegradable polymers like polyhydroxybutyrate and polylactic acid can be broken down by microorganisms during AD [10,105]. However, most micro(nano)plastics are resistant to microbial degradation due to their durable chemical structures, limiting their breakdown in AD systems [106]. In addition to the limited degradation of micro(nano)plastics, harmful byproducts can leach during anaerobic digestion, inhibiting microbial activity and negatively affecting the process, with their release not directly proportional to the concentration of micro(nano)plastics [10].
During anaerobic digestion, microplastics like polyethylene terephthalate undergo various changes, particularly in their chemical structure and surface morphology. Fourier transform infrared spectroscopy analysis reveals that both untreated and pretreated polyethylene terephthalate show a decrease in absorption intensity, indicating alterations in the chemical bonds as a result of both pretreatment and microbial activity [96]. Surface morphology changes are especially prominent in pretreated polyethylene terephthalate, where scanning electron microscope imaging shows surface compression, thinning, and in some cases blistering [96]. These physical changes make the material more susceptible to microbial degradation. However, the crystallinity of polyethylene terephthalate remains largely unaffected by AD, although pretreatment with alkaline thermal hydrolysis slightly increases it by about 13%, likely due to polymer chain reorganization [96].
Given that most micro(nano)plastics resist degradation, a significant proportion remains in the sludge even after AD process, posing a major environmental concern when applied to land [107]. Studies have indicated that the annual amount of micro(nano)plastics released into the terrestrial environment via digested sludge application is far greater than the amount entering oceans and freshwater systems, making sludge treatment a critical pathway for micro(nano)plastics to enter the environment [107].
7. Future perspectives
7.1 Comprehensive analysis of the combined effects of various micro(nano)plastics
To date, most previous studies have focused on the impacts of single types of micro(nano)plastics on microbes within AD systems. However, these studies are still a simplification of what micro(nano)plastics are like in the real world, which is typically replete with a heterogeneous mixture of various micro(nano)plastics [12,108]. The co-exposure effects of the micro(nano)plastics on microbes cannot be merely aggregated from the individual component effects. When different types of micro(nano)plastics aggregate, their interactions may result in either antagonistic or synergistic effects that alter their collective environmental dynamics [109]. For instance, the formation of clusters by entangled fibrous micro(nano)plastics can obscure other forms of micro(nano)plastics embedded within these aggregates [110]. Moreover, the degradation of diverse micro(nano)plastics can lead to the release of various additives, which may interact chemically, yielding complex reactions that could influence both the degradation pathways and the toxicity profiles of these micro(nano)plastics [111]. To accurately assess the environmental risks associated with microplastic pollution, it is crucial to adopt a holistic perspective that integrates the multidimensional impacts of microplastic mixtures.
To achieve this objective, a comprehensive evaluation of the complete micro(nano)plastics will be conducted to assess their ecotoxicological risks, rather than limiting the research to a single polymer, shape, or size. By adjusting the relative proportions of different types of micro(nano)plastics at a fixed concentration and integrating machine learning, a concentration-composition-effect model can be developed, which can identify which characteristics of microplastics pose higher risks. Furthermore, by integrating external environmental parameters and human impacts, we can identify external drivers of microplastic risks, thereby aiding in the formulation of potential mitigation strategies.
7.2 Revealing deep microbial inhibition mechanisms via multi-omics analysis
Current studies on the inhibition of micro(nano)plastics on microbial communities in sludge anaerobic digestion systems primarily focus on exploring the impacts of micro(nano)plastics on microbial diversity and the abundance of specific microbes, often overlooking the in-depth exploration of microbial community ecological functions and their interactions. The complex anaerobic digestion process involves the synergistic actions of multiple microbes. These functional microbes play a variety of crucial roles in sludge anaerobic digestion, including element cycling, information exchange, and electron transfer, through cooperation and competition [112]. The introduction of micro(nano)plastics may disrupt these established cooperative and competitive interactions, leading to an imbalance in microbial community functions and triggering extensive effects across different ecological levels [113]. However, current single-gene sequencing analysis methods may be insufficient to fully reveal the multi-functional impacts of micro(nano)plastics on microorganisms, including electron transfer pathways, functional microbes, and metabolic functions.
Therefore, an integrated approach that combines genomics, transcriptomics, proteomics, and metabolomics is essential to enhance our understanding of the underlying mechanisms of micro(nano)plastics inhibiting microbial activity. The application of multi-omics technology could not only reveal the direct impacts of micro(nano)plastics on microbial genes expression, protein production, and metabolic functions, but also provide insights into how these microbial functions interact within the microbial ecosystem under micro(nano)plastics pressure. Moreover, utilizing these advanced high-resolution techniques enables researchers to pinpoint specific genes and pathways that are either activated or inhibited by micro(nano)plastics. Furthermore, integrating these diverse data streams through systems biology approach and machine learning models will enable the construction of comprehensive models that predict microbial behavior under the stress of micro(nano)plastics, which aids in crafting targeted interventions to mitigate the inhibition effects effectively.
7.3 Alleviating micro(nano)plastic inhibition through bioaugmentation technology
The inhibition effects of micro(nano)plastics on microbes have been widely confirmed. Considering that the refractory characteristics of micro(nano)plastics and the core roles of functional microorganisms in the anaerobic digestion system, it is essential to strengthen the microbial metabolic activity and resistance to micro(nano)plastics. Thus, bioaugmentation of natural or engineered anaerobic microbial consortia capable of degrading plastics is considered to facilitate the decomposition of micro(nano)plastics in anaerobic sludge digesters [114]. To date, several anaerobes, such as Geobacter spp., Holomonas spp., Cloacamonales, and Thermotogales have shown significant potential for microplastic degradation, but their application in anaerobic digesters has yet to be explored [115]. Therefore, it is essential to screen for anaerobic microorganisms that can efficiently degrade micro(nano)plastics. From the perspective of micro(nano)plastics degradation, the limitation of monoculture bioaugmentation is that the introduced bacterial species may only effectively degrade one or a few types of micro(nano)plastics. Using a microbial consortium could degrade a broader range of micro(nano)plastics, and the immobilization of these bacteria could further enhance biodegradation efficiency. Moreover, bioaugmentation with immobilized methods for degrading micro(nano)plastics also holds great potential to promote biodegradation of micro(nano)plastics and enhance methane production.
In addition to screening natural microbes, genetically engineered microbes have recently been proposed as promising candidates for bioaugmentation of AD, though further studies on this application are still needed [115]. Genetic engineering provides the benefit of incorporating genes that code for enzymes specific to the degradation of certain micro(nano)plastics into a host anaerobe, such as integrating genes coding for specific microplastic-degrading enzymes into Geobacter spp., thereby facilitating anaerobic degradation of micro(nano)plastics [116]. Moreover, when Geobacter metallireducens is co-cultured with Methanosaeta harundinacea and ethanol is added, there is an increase in hydrogen production [117]. This indicated that the potential synergistic benefit of some recombinant Geobacter species in improving biogas production. Further studies have found that Clostridium thermocellum, a thermophilic anaerobe, genetically engineered to incorporate a thermophilic cutinase gene from a plant compost metagenome, was able to degrade over 60% of polyethylene terephthalate films by mass after a 14-day incubation at 60 ℃, significantly exceeding previously reported biodegradation rates [118]. These successful cases indicate that there is considerable potential to genetically engineering microbes engaged in AD system for simultaneous microplastic biodegradation and generate biogas.
8. Conclusions
This study systematically discusses the negative impacts of micro(nano)plastics on microbial activity in AD systems. The inhibition mechanisms include cell damage through direct contact, leaching toxic additives, and adsorption of other pollutants. Micro(nano)plastics reduce microbial diversity, increase oxidative stress, and inhibit key enzyme activities, while also disrupting methanogenesis-related functional genes and enhancing ARGs proliferation. The size and dosage of micro(nano)plastics are critical factors in microbial inhibition. Effective mitigation strategies, such as activated carbon addition, sludge thermal hydrolysis, and cationic polyacrylamide addition, have been proposed to alleviate these effects. Most current studies oversimplify by focusing on single types of micro(nano)plastics, overlook the in-depth exploration of microbial community ecological functions, and lack research on alleviating the inhibitory effects of micro(nano)plastics. Therefore, future research should focus on three main areas, including comprehensive analysis of how different types of micro(nano)plastics interact and affect AD systems, using multi-omics analysis to reveal the detailed mechanisms of microbial inhibition, and exploring bioaugmentation to introduce beneficial microbes that can reduce the negative effects of micro(nano)plastics and improve AD system performance.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
CRediT authorship contribution statement
Weixin Zhao: Writing – review & editing, Writing – original draft, Methodology, Conceptualization. Shuocheng Shao: Writing – review & editing, Writing – original draft. Hao Ma: Writing – review & editing. Jianyuan Zhen: Writing – review & editing. Shufei He: Writing – review & editing. Chuandong Wu: Writing – review & editing. Liangliang Wei: Writing – review & editing, Supervision, Funding acquisition.
Acknowledgments
This work was financially sponsored by the funding from the National Key Research and Development Program of China (No. 2023YFC3207404), the National Nature Science Foundation of China (No. 52270034), and State Key Laboratory of Urban Water Resource and Environment (No. 2023TS16).
-
-
[1]
E.D. Okoffo, E. Donner, S.P. McGrath, et al., Water Res. 201 (2021) 117367. doi: 10.1016/j.watres.2021.117367
-
[2]
Y. Zhou, X. Ren, T. Tsui, et al., J. Hazard. Mater. 445 (2023) 130596. doi: 10.1016/j.jhazmat.2022.130596
-
[3]
Y. Sun, W. Ding, Y. Wang, et al., Chin. Chem. Lett. 36 (2025) 109941. doi: 10.1016/j.cclet.2024.109941
-
[4]
X. Shi, Z. Chen, L. Wu, et al., Curr. Opin. Environ. Sci. Health 31 (2023) 100433. doi: 10.1016/j.coesh.2022.100433
-
[5]
M.K. Nguyen, M. Hadi, C. Lin, et al., Chemosphere 308 (2022) 136455. doi: 10.1016/j.chemosphere.2022.136455
-
[6]
M. Nguyen, C. Lin, H. Nguyen, et al., J. Environ. Manag. 346 (2023) 118982. doi: 10.1016/j.jenvman.2023.118982
-
[7]
Y. Zhang, W. Wei, Q. Huang, et al., Water Res. 179 (2020) 115898. doi: 10.1016/j.watres.2020.115898
-
[8]
C. Wang, Z. Zhang, X. Yi, Chin. Chem. Lett. 34 (2023) 108182. doi: 10.1016/j.cclet.2023.108182
-
[9]
S. Liu, C. Su, Y. Lu, et al., Sci. Total Environ. 902 (2023) 166033. doi: 10.1016/j.scitotenv.2023.166033
-
[10]
Z. He, W. Yang, Y. Ren, et al., Bioresour. Technol. 331 (2021) 125035. doi: 10.1016/j.biortech.2021.125035
-
[11]
X. Li, L. Chen, Q. Mei, et al., Water Res. 142 (2018) 75–85. doi: 10.1016/j.watres.2018.05.034
-
[12]
S. Mohammad Mirsoleimani Azizi, F.I. Hai, W. Lu, et al., Bioresour. Technol. 329 (2021) 124894. doi: 10.1016/j.biortech.2021.124894
-
[13]
H. Sun, J. Hu, Y. Wu, et al., J. Hazard. Mater. 465 (2024) 133144. doi: 10.1016/j.jhazmat.2023.133144
-
[14]
W. Zhao, X. Chen, H. Ma, et al., Bioresour. Technol. 400 (2024) 130678. doi: 10.1016/j.biortech.2024.130678
-
[15]
W. Zhao, J. You, S. Yin, et al., Bioresour. Technol. 367 (2023) 128273. doi: 10.1016/j.biortech.2022.128273
-
[16]
W. Zhao, T. Hu, H. Ma, et al., Bioresour. Technol. 391 (2024) 129929. doi: 10.1016/j.biortech.2023.129929
-
[17]
X. Liu, Q. Lu, M. Du, et al., Environ. Sci. Technol. 56 (2022) 11277–11287. doi: 10.1021/acs.est.2c00062
-
[18]
L. Chen, J.J. Huang, B. Hua, et al., Chemosphere 257 (2020) 127291. doi: 10.1016/j.chemosphere.2020.127291
-
[19]
W. Wei, X. Chen, B. Ni, Environ. Sci. Technol. 55 (2021) 11274–11283. doi: 10.1021/acs.est.1c02300
-
[20]
Y. Zhang, W. Wei, J. Sun, et al., Environ. Sci. Technol. 54 (2020) 9662–9671. doi: 10.1021/acs.est.0c02433
-
[21]
S. Zhang, X. Huang, W. Dong, et al., Sci. Total Environ. 939 (2024) 173518. doi: 10.1016/j.scitotenv.2024.173518
-
[22]
J. Wang, D. Ma, K. Feng, et al., Water Res. 219 (2022) 118606. doi: 10.1016/j.watres.2022.118606
-
[23]
M. Nguyen, M.R.J. Rakib, C. Lin, et al., TrAC Trends Anal. Chem. 168 (2023) 117294. doi: 10.1016/j.trac.2023.117294
-
[24]
B. Gao, F. Gao, X. Zhang, et al., Sci. Total Environ. 933 (2024) 173100. doi: 10.1016/j.scitotenv.2024.173100
-
[25]
S. Mohammad Mirsoleimani Azizi, B.S. Zakaria, N. Haffiez, et al., Bioresour. Technol. 377 (2023) 128938. doi: 10.1016/j.biortech.2023.128938
-
[26]
Y. Xiang, W. Xiong, Z. Yang, et al., Chem. Eng. J. 455 (2023) 140754. doi: 10.1016/j.cej.2022.140754
-
[27]
M.K. Manu, L. Luo, R. Kumar, et al., Environ. Pollut. 325 (2023) 121426. doi: 10.1016/j.envpol.2023.121426
-
[28]
S.M.M. Azizi, N. Haffiez, B.S. Zakaria, et al., ACS ES&T Eng. 2 (2022) 1306–1315. doi: 10.1021/acsestengg.1c00460
-
[29]
S. Fu, J. Ding, Y. Zhang, et al., Sci. Total Environ. 625 (2018) 64–70. doi: 10.1016/j.scitotenv.2017.12.158
-
[30]
W. Kong, M. Jalalah, S.A. Alsareii, et al., J. Environ. Chem. Eng. 11 (2023) 111408. doi: 10.1016/j.jece.2023.111408
-
[31]
T. Zhao, L. Tan, X. Zhu, et al., Mar. Pollut. Bull. 154 (2020) 111074. doi: 10.1016/j.marpolbul.2020.111074
-
[32]
X. Liu, Q. Deng, Y. Zheng, et al., Water Res. 221 (2022) 118780. doi: 10.1016/j.watres.2022.118780
-
[33]
S. Wang, X. Wang, M. Fessler, et al., Water Res. 221 (2022) 118844. doi: 10.1016/j.watres.2022.118844
-
[34]
J.N. Hahladakis, C.A. Velis, R. Weber, et al., J. Hazard. Mater. 344 (2018) 179–199. doi: 10.1016/j.jhazmat.2017.10.014
-
[35]
H. Chen, Z. Zou, M. Tang, et al., J. Hazard. Mater. 443 (2023) 130158. doi: 10.1016/j.jhazmat.2022.130158
-
[36]
Y. Chen, Y. Zhang, Z. Zhang, Environ. Pollut. 287 (2021) 117568. doi: 10.1016/j.envpol.2021.117568
-
[37]
R. Xu, J. Cao, T. Ye, et al., Water Res. 223 (2022) 118975. doi: 10.1016/j.watres.2022.118975
-
[38]
T. Luo, X. Dai, W. Wei, et al., Environ. Sci. Technol. 57 (2023) 14611–14621. doi: 10.1021/acs.est.3c02815
-
[39]
W. Zhao, T. Hu, H. Ma, et al., Sci. Total Environ. 901 (2023) 166551. doi: 10.1016/j.scitotenv.2023.166551
-
[40]
W. Zhao, J. You, S. Yin, et al., Environ. Sci. Ecotechnol. 13 (2023) 100212. doi: 10.1016/j.ese.2022.100212
-
[41]
S. Sethulekshmi, A. Shriwastav, Water Res. 250 (2024) 121029. doi: 10.1016/j.watres.2023.121029
-
[42]
H. Chen, M. Tang, X. Yang, et al., Chem. Eng. J. 408 (2021) 127251. doi: 10.1016/j.cej.2020.127251
-
[43]
X. Zhao, L. Zhan, B. Xie, et al., Chemosphere 207 (2018) 742–752. doi: 10.1016/j.chemosphere.2018.05.156
-
[44]
N. Rafa, B. Ahmed, F. Zohora, et al., Environ. Pollut. 343 (2024) 123190. doi: 10.1016/j.envpol.2023.123190
-
[45]
D. Wang, Q. Pan, J. Yang, et al., Environ. Sci. Technol. 58 (2024) 2598–2614. doi: 10.1021/acs.est.3c09239
-
[46]
S. Zhuang, J. Wang, Sci. Total Environ. 897 (2023) 165414. doi: 10.1016/j.scitotenv.2023.165414
-
[47]
Y. Zhang, X. Su, N.F.Y. Tam, et al., Chin. Chem. Lett. 33 (2022) 5213–5217. doi: 10.3390/polym14235213
-
[48]
X. Zheng, L. Zhu, Z. Xu, et al., Sci. Total Environ. 799 (2021) 149394. doi: 10.1016/j.scitotenv.2021.149394
-
[49]
W. Yan, N. Hamid, S. Deng, et al., J. Hazard. Mater. 397 (2020) 122795. doi: 10.1016/j.jhazmat.2020.122795
-
[50]
Q. Liu, H. Wu, J. Chen, et al., Environ. Res. 214 (2022) 113777. doi: 10.1016/j.envres.2022.113777
-
[51]
P. Wang, Y. Guo, M. Yu, et al., Biochem. Eng. J. 198 (2023) 109012. doi: 10.1016/j.bej.2023.109012
-
[52]
M. Sun, K. Xiao, Y. Zhu, et al., Environ. Res. 214 (2022) 114032. doi: 10.1016/j.envres.2022.114032
-
[53]
Z. Zhen, J. Luo, Y. Su, et al., Environ. Sci. Pollut. Res. 30 (2023) 121584–121598. doi: 10.1007/s11356-023-30935-5
-
[54]
J. Liu, G. Xu, S. Zhao, et al., mLife 2 (2023) 378–388. doi: 10.1002/mlf2.12090
-
[55]
S. Wang, D. Zeng, B. Jin, et al., Chem. Eng. J. 452 (2023) 139520. doi: 10.1016/j.cej.2022.139520
-
[56]
C. Hung, C. Huang, S. Hsieh, et al., Chemosphere 297 (2022) 134113. doi: 10.1016/j.chemosphere.2022.134113
-
[57]
W. Wei, Q. Huang, J. Sun, et al., Environ. Sci. Technol. 53 (2019) 2509–2517. doi: 10.1021/acs.est.8b07069
-
[58]
X. Wang, Y. Zhang, Y. Zhao, et al., J. Hazard. Mater. 446 (2023) 130717. doi: 10.1016/j.jhazmat.2022.130717
-
[59]
W. Wei, Q. Hao, Z. Chen, et al., Water Res. 182 (2020) 116041. doi: 10.1016/j.watres.2020.116041
-
[60]
T. Luo, X. Dai, Z. Chen, et al., Water Res. 228 (2023) 119356. doi: 10.1016/j.watres.2022.119356
-
[61]
Y. Xiang, W. Xiong, Z. Yang, et al., Chem. Eng. J. 486 (2024) 149979. doi: 10.1016/j.cej.2024.149979
-
[62]
Y. Feng, L. Feng, S. Liu, et al., Environ. Sci. Water Res. Technol. 4 (2018) 1902–1911. doi: 10.1039/c8ew00535d
-
[63]
Z. Shang, R. Wang, X. Zhang, et al., Sci. Total Environ. 875 (2023) 162674. doi: 10.1016/j.scitotenv.2023.162674
-
[64]
X. Tang, B. Zhang, C. Fan, et al., Chem. Eng. J. 427 (2022) 131442. doi: 10.1016/j.cej.2021.131442
-
[65]
C. Jeong, E. Won, H. Kang, et al., Environ. Sci. Technol. 50 (2016) 8849–8857. doi: 10.1021/acs.est.6b01441
-
[66]
H. Wang, Q. Zhou, J. Clean. Prod. 450 (2024) 141931. doi: 10.1016/j.jclepro.2024.141931
-
[67]
Y. Li, X. Li, P. Wang, et al., J. Hazard. Mater. 438 (2022) 129493. doi: 10.1016/j.jhazmat.2022.129493
-
[68]
Z. Zhang, Y. Chen, Chem. Eng. J. 382 (2020) 122955. doi: 10.1016/j.cej.2019.122955
-
[69]
Y. Li, W. Zhong, Z. Ning, et al., Bioresour. Technol. 364 (2022) 128052. doi: 10.1016/j.biortech.2022.128052
-
[70]
X. Wang, G. Huang, Q. Chen, et al., Environ. Res. 252 (2024) 119061. doi: 10.1016/j.envres.2024.119061
-
[71]
X. Tang, M. Zhou, G. Zeng, et al., Chem. Eng. J. 435 (2022) 134734. doi: 10.1016/j.cej.2022.134734
-
[72]
L. Zhang, J. Sun, Z. Zhang, et al., Water Res. 217 (2022) 118426. doi: 10.1016/j.watres.2022.118426
-
[73]
Y. Cheng, X. Hu, Y. Gao, et al., Bull. Environ. Contam. Toxicol. 112 (2023) 5.
-
[74]
A. Pires, A. Cuccaro, M. Sole, et al., Environ. Res. 214 (2022) 113642. doi: 10.1016/j.envres.2022.113642
-
[75]
X. He, Y. Xiang, R. Xu, et al., Environ. Sci. Pollut. Res. 31 (2024) 19635–19648. doi: 10.1007/s11356-024-32422-x
-
[76]
E.R.M. Kelly, J.E. Trujillo, A. Setiawan, et al., Mar. Pollut. Bull. 203 (2024) 116438. doi: 10.1016/j.marpolbul.2024.116438
-
[77]
A. Arsalan, H. Younus, Int. J. Biol. Macromol. 118 (2018) 1833–1847. doi: 10.1016/j.ijbiomac.2018.07.030
-
[78]
K. Holmberg, Colloids Surf. B Biointerfaces 168 (2018) 169–177. doi: 10.1016/j.colsurfb.2017.12.002
-
[79]
D. Wang, L. Yan, X. Ma, et al., Int. J. Biol. Macromol. 119 (2018) 453–461. doi: 10.1016/j.ijbiomac.2018.07.133
-
[80]
H.E.G. Akbay, C. Akarsu, Z. Isik, et al., Biochem. Eng. J. 187 (2022) 108619. doi: 10.1016/j.bej.2022.108619
-
[81]
L. Fu, J. Li, G. Wang, et al., Ecotoxicol. Environ. Saf. 217 (2021) 112207. doi: 10.1016/j.ecoenv.2021.112207
-
[82]
Z. Gao, H. Qian, T. Cui, et al., Chem. Eng. J. 484 (2024) 149703. doi: 10.1016/j.cej.2024.149703
-
[83]
H. Liu, X. Zhou, W. Ding, et al., ACS Sustain. Chem. Eng. 7 (2019) 20097–20101. doi: 10.1021/acssuschemeng.9b05960
-
[84]
C. Mallikarjuna, R.R. Dash, J. Environ. Chem. Eng. 8 (2020) 104233. doi: 10.1016/j.jece.2020.104233
-
[85]
H. Liu, H. Xu, C. Ma, et al., Rev. Environ. Sci. Biotechnol. 21 (2022) 469–482. doi: 10.1007/s11157-022-09614-3
-
[86]
X. Yang, Y. Zhou, R. Xia, et al., J. Hazard. Mater. 463 (2024) 132900. doi: 10.1016/j.jhazmat.2023.132900
-
[87]
S. Wieland, A. Balmes, J. Bender, et al., J. Hazard. Mater. 428 (2022) 128151. doi: 10.1016/j.jhazmat.2021.128151
-
[88]
Q. Guo, M. Wang, S. Jin, et al., Sci. Total Environ. 919 (2024) 170824. doi: 10.1016/j.scitotenv.2024.170824
-
[89]
M. Li, Z. Li, F. Chen, et al., J. Hazard. Mater. 470 (2024) 134286. doi: 10.1016/j.jhazmat.2024.134286
-
[90]
Q. Wang, Q. Wang, T. Wang, et al., Sci. Total Environ. 917 (2024) 170541. doi: 10.1016/j.scitotenv.2024.170541
-
[91]
Z. Chen, X. Liu, W. Wei, et al., Water Res. 221 (2022) 118820. doi: 10.1016/j.watres.2022.118820
-
[92]
J. Zhang, F. Mao, K. Loh, et al., Bioresour. Technol. 249 (2018) 729–736. doi: 10.1016/j.biortech.2017.10.082
-
[93]
V. Toutian, M. Barjenbruch, C. Loderer, et al., Water Res. 202 (2021) 117465. doi: 10.1016/j.watres.2021.117465
-
[94]
T. Luo, H. Huang, Z. Mei, et al., Chin. Chem. Lett. 30 (2019) 1219–1223. doi: 10.1016/j.cclet.2019.03.018
-
[95]
Y. Zeng, X. Tang, C. Fan, et al., ACS ES&T Water 2 (2022) 117–127. doi: 10.1021/acsestwater.1c00294
-
[96]
M.D. Hatinoglu, F. Dilek Sanin, Waste Manag. 153 (2022) 376–385. doi: 10.1016/j.wasman.2022.09.016
-
[97]
S.M.M. Azizi, B.R. Dhar, Chemosphere 353 (2024) 141516. doi: 10.1016/j.chemosphere.2024.141516
-
[98]
S.M. Mirsoleimani Azizi, N. Haffiez, A. Mostafa, et al., Renew. Sustain. Energy Rev. 200 (2024) 114453. doi: 10.1016/j.rser.2024.114453
-
[99]
S.M. Mirsoleimani Azizi, B.S. Zakaria, N. Haffiez, et al., ACS Sustain. Chem. Eng. 11 (2023) 7253–7262. doi: 10.1021/acssuschemeng.3c01349
-
[100]
B. Ma, W. Xue, C. Hu, et al., Chem. Eng. J. 359 (2019) 159–167. doi: 10.1016/j.cej.2018.11.155
-
[101]
H.P.H. Arp, H. Knutsen, Environ. Sci. Technol. 54 (2020) 3–5. doi: 10.1021/acs.est.9b07089
-
[102]
M. Lapointe, J.M. Farner, L.M. Hernandez, et al., Environ. Sci. Technol. 54 (2020) 8719–8727. doi: 10.1021/acs.est.0c00712
-
[103]
B. Zhang, X. Tang, C. Fan, et al., Water Res. 205 (2021) 117651. doi: 10.1016/j.watres.2021.117651
-
[104]
Y. Wang, X. Liu, W. Han, et al., Bioresour. Technol. 400 (2024) 130649. doi: 10.1016/j.biortech.2024.130649
-
[105]
L. Hou, D. Kumar, C.G. Yoo, et al., Chem. Eng. J. 406 (2021) 126715. doi: 10.1016/j.cej.2020.126715
-
[106]
E.F. Gómez, F.C. Michel, Polym. Degrad. Stab. 98 (2013) 2583–2591. doi: 10.1016/j.polymdegradstab.2013.09.018
-
[107]
A. Cydzik-Kwiatkowska, N. Milojevic, P. Jachimowicz, Sci. Total Environ. 848 (2022) 157466. doi: 10.1016/j.scitotenv.2022.157466
-
[108]
W. Yang, P. Gao, Z. Ye, et al., Sci. Total Environ. 934 (2024) 173218. doi: 10.1016/j.scitotenv.2024.173218
-
[109]
W. Wei, C. Wang, X. Shi, et al., Water Res. 222 (2022) 118895. doi: 10.1016/j.watres.2022.118895
-
[110]
J. Li, M. Dagnew, M.B. Ray, J. Environ. Manag. 346 (2023) 118792. doi: 10.1016/j.jenvman.2023.118792
-
[111]
J.H. Bridson, R. Abbel, D.A. Smith, et al., J. Hazard. Mater. 459 (2023) 132303. doi: 10.1016/j.jhazmat.2023.132303
-
[112]
N. Kim, S. Lee, Y. Kim, et al., Bioresour. Technol. 344 (2022) 126350. doi: 10.1016/j.biortech.2021.126350
-
[113]
X. Ren, M. Mao, M. Feng, et al., Water Res. 251 (2024) 121121. doi: 10.1016/j.watres.2024.121121
-
[114]
K.H.D. Tang, T. Hadibarata, Bioprocess Biosyst. Eng. 45 (2022) 1865– 1878. doi: 10.1007/s00449-022-02793-x
-
[115]
K.H.D. Tang, J. Clean. Prod. 387 (2023) 135864. doi: 10.1016/j.jclepro.2023.135864
-
[116]
K. Rani, P. Singh, R. Agarwal, et al., Chapter 3–Concomitant degradation of petroleum products and microplastics in industrial wastewater using genetically modified microorganisms. in: M.H. El-Naas, A. Banerjee (Eds.), Petroleum Industry Wastewater, Elsevier, 2022, pp. 19–34.
-
[117]
A. Mostafa, S. Im, Y. Song, et al., Processes 8 (2020) 424. doi: 10.3390/pr8040424
-
[118]
F. Yan, R. Wei, Q. Cui, et al., Microb. Biotechnol. 14 (2021) 374–385. doi: 10.1111/1751-7915.13580
-
[119]
W. Wei, Q. Huang, J. Sun, et al., Environ. Sci. Technol. 53 (2019) 9604–9613. doi: 10.1021/acs.est.9b02971
-
[120]
J. Shi, Q. Dang, C. Zhang, et al., Environ. Pollut. 310 (2022) 119859. doi: 10.1016/j.envpol.2022.119859
-
[1]
-
Figure 1 Schematic diagram of the toxic effects of micro(nano)plastics on microbes. Modified with permission [27]. Copyright 2023, Elsevier.
Figure 3 Mechanism diagram of the effects of polycarbonate micro(nano)plastics on the enzyme activity and ROS in WAS anaerobic digestion. Modified with permission [35]. Copyright 2023, Elsevier.
Figure 4 Potential mechanisms of the enhanced horizontal flow of ARGs induced by micro(nano)plastics in anaerobic sludge digestion. Modified with permission [72]. Copyright 2022, Elsevier.
Table 1. Impacts of micro(nano)plastics on methane production performance and microbial community variation in AD system.
Micro(nano)plastics type Dosage Size Substrate Digestion mode Methane production performance Bacteria variation Methanogen variation Ref. Polystyrene 150 µg/L 50 nm WAS Batch, 37 ℃, 300 rpm Methane yield decreased by 29.34% Cloacamonaceae_W22 decreased by 14.6% Methanosaeta decreased by 16% [28] Polyvinyl Chloride 60 particles/g TS 1 mm WAS Batch, 37 ± 1 ℃ Methane yield decreased by 24.2% Proteiniborus decreased by 25% Methanosaeta decreased by 16.5% [57] Polyethylene 200 particles/g TS 40 ± 2 µm WAS Batch, 37 ± 1 ℃ Methane yield decreased by 27.5% Rhodobacter decreased by 15.2% Methanosaeta decreased by 16% [119] Polystyrene 160 particles/g TS 100 µm WAS Batch, 37 ℃, 150 rpm Methane yield decreased by 11.04% Leptolinea decreased by 1.99% Methanobacterium decreased by 5.31% [39] Polyvinyl chloride 2.4 g/g VS 3000 µm WAS Semi-continuous, OLR = 15 g VS/L/d Methane yield improved by 34.9% Acidobacteriota improved by 17.7% Methanosaeta decreased by 53.6% [52] Polystyrene 150 µg/L 50 nm WAS Batch, 37 ± 2 ℃, 300 rpm Methane yield decreased by 32.3% Treponema completely disappeared Methanosaeta decreased by 14% [91] Polystyrene 50 mg/g TS 50 nm WAS Batch, 35 ± 2 ℃ Methane yield decreased by 15.5% Sulfurovum decreased by 36.6% Candidatus Methanofastidiosum
decreased by 61%[22] Polyethylene 100 mg/g TS 180 µm WAS Batch, 37 ℃, 120 rpm Methane yield decreased by 13.8% Atopostipes increased by 2.1% Methanosaeta decreased by 3.4% [120] Polycarbonate 30 particles/g TS 40 µm WAS Batch, 37 ± 1 ℃, 300 rpm Methane yield improved by 24.7% Longilinea increased by 1.34% Methanosaeta decreased by 8.4% [35] Polyvinyl chloride 30 mg/g TS 0.35 mm WAS Batch, 35 ± 1 ℃, 140 rpm Methane yield decreased by 15.62% Actinobacteria decreased by 12% Methanosaeta decreased by 1.1% [64]  下载: 导出CSV
下载: 导出CSV
-









 扫一扫看文章
扫一扫看文章
计量
- PDF下载量: 0
- 文章访问数: 150
- HTML全文浏览量: 10
 下载:
下载:
