Table 1.
PfA-M1 and PfA-M17 inhibitory activities of compounds 7–18.
Citation:
Anil Kumar Marapaka, Priyanka Sankoju, Guozhen Zhang, Yongzheng Ding, Chunhua Ma, Vijaykumar Pillalamarri, Renu Sudhakar, Bharati Reddi, Puran Singh Sijwali, Yingjie Zhang, Anthony Addlagatta. Development of peptidomimetic hydroxamates as PfA-M1 and PfA-M17 dual inhibitors: Biological evaluation and structural characterization by cocrystallization[J]. Chinese Chemical Letters,
2022, 33(5): 2550-2554.
doi:
10.1016/j.cclet.2021.09.102

Development of peptidomimetic hydroxamates as PfA-M1 and PfA-M17 dual inhibitors: Biological evaluation and structural characterization by cocrystallization
English
Development of peptidomimetic hydroxamates as PfA-M1 and PfA-M17 dual inhibitors: Biological evaluation and structural characterization by cocrystallization
Abstract:
Plasmodium parasites causing malaria have developed resistance to most of the antimalarials in use, including the artemisinin-based combinations, which are the last line of defense against malaria. This necessitates the discovery of new targets and the development of novel antimalarials. Plasmodium falciparum alanyl aminopeptidase (PfA-M1) and leucyl aminopeptidase (PfA-M17) belong to the M1 and M17 family of metalloproteases respectively and play critical roles in the asexual erythrocytic stage of development. These enzymes have been suggested as potential antimalarial drug targets. Herein we describe the development of peptidomimetic hydroxamates as PfA-M1 and PfA-M17 dual inhibitors. Most of the compounds described in this study display inhibition at sub-micromolar range against the recombinant PfA-M1 and PfA-M17. More importantly, compound 26 not only exhibits potent malarial aminopeptidases inhibitory activities (PfA-M1 Ki = 0.11 ± 0.0002 µmol/L, PfA-M17 Ki = 0.05 ± 0.005 µmol/L), but also possesses remarkable selectivity over the mammalian counterpart (pAPN Ki = 17.24 ± 0.08 µmol/L), which endows 26 with strong inhibition of the malarial parasite growth and negligible cytotoxicity on human cell lines. Crystal structures of PfA-M1 at atomic resolution in complex with four different compounds including compound 26 establish the structural basis for their inhibitory activities. Notably, the terminal ureidobenzyl group of 26 explores the S2′ region where differences between the malarial and mammalian enzymes are apparent, which rationalizes the selectivity of 26. Together, our data provide important insights for the rational and structure-based design of selective and dual inhibitors of malarial aminopeptidases that will likely lead to novel chemotherapeutics for the treatment of malaria.
-
Key words:
- Aminopeptidase
- / Dual inhibitor
- / Antimalaria
- / Peptidomimetic
- / Plasmodium falciparum
-
Malaria, the world's most prevalent parasitic disease, is caused by the protozoan parasite called Plasmodium [1]. According to the World Malaria Report 2020, the number of malaria cases exceeded 220 million and around 400,000 deaths from malaria occurred in 2019, mostly children in Africa [2]. Traditional chemotherapeutic strategies for malaria control can be categorized mainly into three types: quinoline-related compounds (quinine, quinidine, chloroquine, mefloquine, etc.), antifolate compounds (proguanil, pyrimethamine, trimethoprim, etc.) and artemisinin derivatives (artemisinin, dihydroartemisinin, artesunate, artemether, arteether, etc.) [3]. Unfortunately, the emergence and widespread of resistant Plasmodium falciparum to most of the currently available antimalarials is alarming and necessitates development of novel anti-malarial drugs [1, 3].
Hemoglobin digestion is an essential metabolic process in the intraerythrocytic stage of the Plasmodium life cycle. During this process, malaria parasites digest host cell hemoglobin to get amino acids required for parasite protein synthesis [4, 5]. Among kinds of metalloami-nopeptidases involved in hemoglobin digestion in Plasmodium falciparum, alanyl aminopeptidase (PfA-M1) and leucyl aminopeptidase (PfA-M17) are two important contributors of single amino acid generation from shorter peptides generated by endopeptidases [6]. PfA-M1 has been demonstrated to cleave hydrophobic and positively charged amino acids with best preference to leucine, alanine, arginine and phenylalanine [7]. On the other hand, PfA-M17 has much narrower substrate specificity exhibiting a strong preference for N-terminal hydrophobic residues, in particular leucine and tryptophan [7]. It is worth noting that gene knockout and specific inhibition of PfA-M1 and PfA-M17 from Plasmodium falciparum were lethal, indicating they are ideal drug targets [8, 9]. Given that both PfA-M1 and PfA-M17 are potential drug targets, several attempts were made to target these two metalloaminopeptidases individually or together by various types of small molecules, which could be mainly categorized into hydroxamates [10-15], phosphates/phosphinates [16-19], amino-benzosuberones [20, 21] and bestatin analogs [9, 22]. Generally, inhibitors that consist of hydroxamate moiety as the zinc-binding group (ZBG) exhibited potent aminopeptidases inhibitory activities and were capable of reaching the site of action within infected red blood cells to exert antimalarial effects [10-15]. For example, compounds 1, 2 and 3 were reported to be potent PfA-M1 inhibitors, among which 1 and 2 exhibited moderate in vitro antimalarial activities [10, 11]. Compounds 4 and 5 could inhibit PfA-M1 and PfA-M17 simultaneously and showed promising in vitro antimalarial activities [12, 13]. Moreover, another PfA-M1 and PfA-M17 dual inhibitor 6 showed oral efficacy against murine malaria [14] (Fig. S1 in Supporting information).
In our previous work, we developed a series of leucine-derived hydroxamates (Fig. S2 in Supporting information) with potent inhibitory activities against mammalian aminopeptidase N (APN) [23, 24]. Considering the structural similarity between the leucine-derived hydroxamates and the reported PfA-M1 inhibitor 3 (Fig. S2), we speculated that these leucine-derived hydroxamates could also inhibit PfA-M1 and even PfA-M17, hence used as antimalarial lead compounds. In the present work, twelve leucine-derived hydroxamates were progressed to in vitro malarial aminopeptidases inhibition assay, which confirmed their potent PfA-M1 and PfA-M17 inhibitory activities. Structural characterization by cocrystallization of three compounds (8, 11 and 18) revealed their binding modes in PfA-M1. Compounds 8 and 18 adopted similar binding modes in the active site of PfA-M1, with their benzyl and isobutyl groups occupying the S1 and S1′ pockets, respectively. Surprisingly, compound 11 bound in PfA-M1 with its isobutyl group in the S1 pocket, its benzyl group reaching into the substrate/product access channel (indicated as the S2′ region), while leaving the S1′ pocket unoccupied. Based on the different binding modes, we suggested that compounds capable of interacting with S1, S1′ pockets and S2′ region simultaneously should exhibit improved malarial aminopeptidases inhibitory activity and selectivity. Therefore, a series of novel peptidomimetic hydroxamates were designed, synthesized and biologically evaluated. Satisfyingly, compound 26 stood out as a potent PfA-M1 and PfA-M17 dual inhibitor with in vitro antimalarial activities. Notably, compound 26 showed significant selectivity over the mammalian counterpart porcine aminopeptidase N (pAPN), which could be rationalized by cocrystal structure analysis.
The leucine-derived hydroxamates 7–18 were previously reported as potent mammalian APN inhibitors with half inhibition concentration (IC50) values against pAPN in the low micromolar to nanomolar range [23, 24]. In this study, the malarial aminopeptidases inhibitory activities of compounds 7–18 were evaluated against both PfA-M1 and PfA-M17 with bestatin as the positive control. The results listed in Table 1 showed that all leucine-derived hydroxamates possessed moderate to potent inhibitory activities against both malarial aminopeptidases. Overall, most compounds showed preference against PfA-M1 over PfA-M17. For example, compounds 7, 11, 13, 15 and 16 inhibited PfA-M1 with over 10-fold greater affinity than PfA-M17. Since the main purpose of current research was identification of PfA-M1 and PfA-M17 dual inhibitors as antimalarial agents, we focused our attention on compounds 8, 17 and 18, which displayed potent and balanced PfA-M1 and PfA-M17 dual inhibition. It is worth noting that the malarial aminopeptidases inhibitory activities of compounds 8, 17 and 18 were more potent than bestatin, therefore, they were progressed to cocrystallization study to provide the basis for rational design of more potent malarial aminopeptidases inhibitors.
Table 1
We have successfully determined the crystal structures of PfA-M1 in complex with compounds 8 and 18 at atomic resolution (Figs. 1A and B and Table S1 in Supporting information). Our attempts to crystallize the PfA-M17 in complex with inhibitors did not yield diffraction quality crystals. As shown in Figs. 1A and B, compounds 8 and 18 adopt similar binding modes in the active site of PfA-M1, where the hydroxamate group bidentately coordinates the catalytic Zn2+, and the benzyl and isobutyl groups interact with S1 and S1′ pockets, respectively. Moreover, both compounds 8 and 18 can form two hydrogen bonds with Y580 via the carbonyl O of hydroxamate group and the ureido NH that is close to the isobutyl group. Note that the binding poses of benzyl groups of compounds 8 and 18 in the S1 pocket are different. To be specific, the chlorobenzyl group of 8 is sandwiched between Y575 and M1034, forming π-π stacking interaction with Y575, hydrogen bond with A320, and hydrophobic interactions with V459, M1034, E572 and E319 (Fig. 1A). In contrast, the dimethylbenzyl group of 18 rotates dramatically and forms π-π T-shaped interaction with Y575, probably to avoid steric clashes resulting from the dimethyl substituents (Fig. 1B). The rotation of dimethylbenzyl group leads to a flip of the side chain of M1034, which might account for the slightly decreased potency of 18 compared with 8. The isobutyl group of both compounds 8 and 18 reaches into the S1′ pocket outlined by V459, G460, V493, H496 and Y580. Although not fully occupying the S1′ pocket, the isobutyl group can form hydrophobic interactions with V459, G460 and Y580 (Figs. 1A and B).
Figure 1

The cocrystal structure of compound 11, a meta-chloro-substituted isomer of 8 was also resolved (Fig. 1C and Table S1). Surprisingly, the binding mode of compound 11 in PfA-M1 is quite different from that of compound 8. Comparing Figs. 1A and C, it is found that the isobutyl and benzyl groups of 11 rotate about 180° around the C-Cα bond relative to compound 8. As a result, the isobutyl group of 11 moves out of S1′ pocket into the S1 pocket, wherein it points to the backwall of S1 pocket (M462, E319 and Q317) and forms hydrophobic interactions with V459 and Y575. However, the chlorobenzyl group of 11 could not occupy the S1′ pocket due to its steric bulk. Instead, the chlorobenzyl group reaches beyond the S1′ pocket and into the substrate/product access channel (S2′ region). It is worth noting that the ureido moiety of compound 11 forms three hydrogen bonds with G460 and A461, which are not observed in compounds 8 and 18. However, the PfA-M1 inhibitory activity of 11 (Ki = 0.23 ± 0.001 µmol/L) is only marginally increased relative to 8 (Ki = 0.41 ± 0.0001 µmol/L) and 18 (Ki = 0.57 ± 0.0002 µmol/L), which could be ascribed to its lack of interaction with the S1′ pocket.
Although the malarial aminopeptidases inhibitory potency of our previous leucine-derived hydroxamates have been confirmed, there are two problem s of these compounds. First, they show limited selectivity over mammalian aminopeptidase. Secondly, their malarial aminopeptidases inhibitory potency, especially against PfA-M17, needs to be further improved. Structural characterization by cocrystallization revealed that these compounds use hydroxamate group to chelate the catalytic Zn2+, which is crucial to their potency. It is worth noting that all these compounds can only occupy two of the three pockets/regions (S1, S1′ and S2′) in PfA-M1 due to their simple structures (Table 2). We wondered if compounds capable of occupying S1, S1′ pockets and S2′ region simultaneously could exhibit improved malarial aminopeptidases inhibitory activity and selectivity. Therefore, a novel series of peptidomimetic hydroxamates were designed, hoping their three branched side chains could occupy the S1, S1′ pockets and the S2′ region simultaneously (Table 2).
Table 2
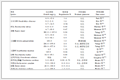 Table 2. Design strategy of novel peptidomimetic hydroxamates 19–26 and their inhibition data against PfA-M1, PfA-M17 and pAPN.
Table 2. Design strategy of novel peptidomimetic hydroxamates 19–26 and their inhibition data against PfA-M1, PfA-M17 and pAPN. DownLoad:
CSV
DownLoad:
CSV

Compounds 19–24 were prepared following the procedures described in Scheme S1 (Supporting information). The amino acid methyl ester hydrochlorides A1–3 were condensed with pyrazinecarboxylic acid to give B1–3, which were hydrolyzed to give C1–3, respectively. Condensation of Boc-protected amino acid E1–3 with l-leucine methyl ester hydrochloride gave intermediate F1–3, which upon deprotection afforded G1–3, respectively. Condensation of C1–3 with l-leucine methyl ester hydrochloride, and condensation of G1–3 with pyrazinecarboxylic acid, gave the common intermediates D1–6, which were reacted with NH2OK in methanol to give target compounds 19–24, respectively.
Compounds 25 and 26 were prepared following the procedures described in Scheme S2 (Supporting information). Condensation of C2 and l-phenylalanine methyl ester hydrochloride led to D7, which was converted into compound 25 with NH2OK in methanol. Intermediate G4 was obtained from Boc-l-leucine and d-phenylglycine methyl ester hydrochloride using the similar method of synthesizing G1–3. Then G4 was connected with benzylamine in the presence of triphosgene to give H, which was further reacted with NH2OK in methanol to give target compound 26.
The newly synthesized compounds 19–26 were progressed to malarial aminopeptidases inhibition assay with bestatin as the positive control. Unsatisfactorily, compounds 19–25 showed dramatically decreased inhibitory activities against both malarial and mammalian enzymes compared to 7–18 (Table 2). Among analogs 19–25, only compound 22 with R1 being H exhibited moderate PfA-M17 inhibitory activity (Ki = 6.96 µmol/L). We speculated that these compounds could not fit well in the enzyme active sites due to steric hindrance resulting from their unsuitable geometry. In contrast, compound 26 with ureido-attached benzyl group, showed remarkable malarial aminopeptidases dual inhibition (PfA-M1 Ki = 0.11 µmol/L, PfA-M17 Ki = 0.05 µmol/L), which were more potent than bestatin and all parent leucine-derived hydroxamates (Table 1). It is worth noting that compound 26 displayed significantly improved PfA-M17 activity compared to most leucine-derived hydroxamates. More importantly, compound 26 showed about 340-fold selectivity towards PfA-M17, 150-fold selectivity towards PfA-M1 over their mammalian counterpart pAPN. The positive control bestatin is a pan-aminopeptidases inhibitor with some preference to pAPN.
The crystal structure of PfA-M1–26 complex was determined at 1.96 Å (Fig. 2 and Table S1). As expected, compound 26 also coordinates the catalytic Zn2+ through the hydroxamate group. The hydrogen bond with Y580 is reserved. More importantly, it can interact with S1, S1′ pockets and S2′ region simultaneously (Fig. 2). To be specific, the phenyl group adjacent to the hydroxamate group is sandwiched between Y575 and V459 in S1 pocket, forming π-π stacking interaction with Y575 and π-σ interaction with V459. The amide O forms dual hydrogen bonds with G460 and A461, which facilitates the hydrophobic isobutyl group to occupy the S1′ pocket, wherein it interacts with the π electrons of H496 and forms hydrophobic interactions with V493 and T492. Remarkably, the ureido-attached benzyl moiety of 26 effectively explores the S2′ region, forming amide-π stacking interaction with the amide moiety connecting T576 and T577.
Figure 2
 Figure 2. The binding mode of compound 26 (green, PDB: 5XM7) in the active site of PfA-M1 (gray cartoon and sticks). The hydrogen bonds are indicated by black dashed lines. The zinc ion coordination is omitted for clarity.
Figure 2. The binding mode of compound 26 (green, PDB: 5XM7) in the active site of PfA-M1 (gray cartoon and sticks). The hydrogen bonds are indicated by black dashed lines. The zinc ion coordination is omitted for clarity.Since we did not get the cocrystallization of the PfA-M17 and compound 26, molecular docking was used to investigate the binding mode of compound 26 in PfA-M17. As shown in Fig. S3A (Supporting information), compound 26 could fit well in the active site of PfA-M17, with the hydroxamate group chelating the catalytic and regulatory zinc ions simultaneously. The coordination between the hydroxamate group and the two zinc ions are clearly presented in Fig. S3B (Supporting information). Similar to its binding mode in PfA-M1, compound 26 could interact with the S1, S1′ pockets and the S2′ region of PfA-M17 via its phenyl, isobutyl and benzyl groups, respectively. To be specific, the phenyl group of 26 is sandwiched between M396 and G489 in the S1 pocket, forming hydrophobic interactions with F398, L492 and A577. The isobutyl group partially occupies the S1′ pocket bordered by A460, N457, I547 and S554. The ureido-attached benzyl group is projected to the solvent-exposed S2′ region, interacting with the loop consist of N384, L385, K386, A387 and A388 (Fig. S3A). Remarkably, compound 26 could form multiple hydrogen bonds with PfA-M17. As shown in Fig. S3B, the hydroxamate group can not only coordinate the two zinc ions, but also form four hydrogen bonds with L487, L374, D459 and L386, respectively. Moreover, three additional hydrogen bonds are observed between the amide moiety and G489, and between the ureido moiety and D459. These hydrogen bonds network together with geometric fitness in the active site could well explain the high affinity of compound 26 towards PfA-M17 (Ki = 0.05 ± 0.005 µmol/L).
To preliminarily investigate the structural basis for malarial aminopeptidases selectivity of 26, we aligned the PfA-M1–26 complex (PDB: 5XM7) on the human APN (hAPN) structure (PDB: 4FYQ) using PyMol (http://www.pymol.org/). As shown in Fig. S4A (Supporting information), compound 26 fits well in the active site of PfA-M1, with the benzyl group reaching to the S2′ region and close to the residue Y575 (also see Fig. 2). In contrast, the analogous residue of Y575 (PfA-M1) is F472 in hAPN which has a collision with the benzyl group of 26 as shown in Fig. S4B (Supporting information). The similar collision is also observed in pAPN, which has F467 as the analogous residue (data not shown). Considering the similar volumes of phenylalanine and tyrosine, it is suggested that the different geometry of S2′ regions between PfA-M1 and mammalian APNs might be the dominant factor of inhibitor selectivity. These results might in part rationalize the malarial aminopeptidases selectivity of 26, moreover, they support our compound design strategy that inhibitors occupying S1, S1′ pockets and S2′ region simultaneously could exhibit improved malarial aminopeptidases inhibitory activity and selectivity.
To evaluate their anti-malarial activities, some compounds were selected and tested against the drug-sensitive (3D7) and drug-resistant (Dd2) strains of Plasmodium falciparum. The results in Table S2 (Supporting information) showed that the most potent PfA-M1 and PfA-M17 dual inhibitor 26 also displayed the best anti-malarial activities against both parasite strains (3D7 IC50 = 3.83 µmol/L, Dd2 IC50 = 6.38 µmol/L). The second best one among these peptidomimetic hydroxamates is compound 17, which is also a potent PfA-M1 and PfA-M17 dual inhibitor. In contrast, the weak malarial aminopeptidases inhibitor 21 only exhibited marginal anti-malarial activity. Overall, the anti-malarial activities of these compounds were in line with their malarial aminopeptidases inhibitory activity. Moreover, the PfA-M1 and PfA-M17 dual inhibitors (17 and 26) possessed superior anti-malarial activities relative to the PfA-M1 selective inhibitor (15).
Considering its significant malarial aminopeptidases selectivity and anti-malarial activity, compound 26 was progressed to in vitro cytotoxicity evaluation against one cancer cell line A549 (adenocarcinomic human alveolar basal epithelial cells) and one normal cell line MRC-5 (human embryonic lung fibroblasts). Satisfyingly, compound 26 inhibited A549 cells growth with IC50 value over 500 µmol/L, which was less cytotoxic than bestatin (IC50 = 487 ± 38 µmol/L) (Table S3 in Supporting information). More importantly, no significant cytotoxicity of 26 was observed against the normal cell line MRC-5 at the concentration of 500 µmol/L (Table S3), indicating the promising safety of 26. In contrast, the positive control doxorubicin exhibited significant cytotoxicity against both A549 (IC50 = 14.5 ± 2.6 µmol/L) and MCR-5 cells (IC50 = 22.1 ± 4.2 µmol/L) (Table S3).
To preliminarily evaluate the metabolic stability of 26, the mouse liver microsome assay was carried out. The reduced nicotinamide adenine dinucleotide phosphate (NADPH) was used as cofactor. The results in Table S4 (Supporting information) showed that the remaining of 26 after 60 min incubation was 67%, indicating a favorable metabolic stability of 26.
PfA-M1 and PfA-M17 are promising anti-malaria targets. In the present study, previously reported APN inhibitors were repositioned as PfA-M1 and PfA-M17 inhibitors. Cocrystal structure analysis revealed the binding modes of several representative compounds in PfA-M1, which guided the rational design of more potent and selective malarial aminopeptidase inhibitors. Among the newly synthesized compounds in this study, compound 26 was demonstrated to be a potent PfA-M1 and PfA-M17 dual inhibitor with significant selectivity over mammalian counterpart. Structural and modeling studies rationalized the inhibitory potency and selectivity against malarial aminopepetidases of 26. Moreover, the potent in vitro antimalarial potency, low host cytotoxicity and favorable mouse liver microsome stability of 26 were also confirmed. To the best of knowledge, compound 26 is the first reported PfA-M1 and PfA-M17 dual inhibitor with significant selectivity over mammalian APN, which provides basis for further development of selective malarial aminopepetidases inhibitors as antimalarial agents.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
Anil Kumar Marapaka and Priyanka Sankoju are the recipients of a fellowship from Council of Scientific and Industrial Research (CSIR), New Delhi, India. Vijaykumar Pillalamarri is supported by University Grants Commission (UGC), New Delhi, India and Bharati Reddi by Department of Science and Technology, New Delhi, India for their research fellowships. CSIR-IICT manuscript number is IICT/Pubs./2018/299. Anthony Addlagatta thanks Science and Engineering Research Board (SERB), New Delhi, India for research grants (Nos. EMR/2015/000461 and CRG/2019/006013). Yingjie Zhang thanks Natural Science Foundation of Shandong Province (No. ZR2018QH007, China), Key Research and Development Program of Shandong Province (No. 2017CXGC1401, China) and Young Scholars Program of Shandong University (No. YSPSDU, 2016WLJH33, China) for research fellowships. Renu Sudhakar is the recipient of the fellowship from the Department of Biotechnology (India). Puran Singh Sijwali lab is supported with funds from the Department of Biotechnology, India (Nos. SR/SO/BB/−0124/2012 and BT/COE/34/SP15138/2015) and the Council of Scientific & Industrial Research, India.
Supplementary materials
Supplementary material associated with this article can be found, in the online version, at doi:10.1016/j.cclet.2021.09.102.
-
-
[1]
B. Mills, R.E. Isaac, R. Foster, J. Med. Chem. 64 (2021) 1763-1785. doi: 10.1021/acs.jmedchem.0c01721
-
[2]
World Health Organization. World Malaria Report 2020; Geneva, 2020. https://www.who.int/teams/global-malaria-programme/reports/worldmalaria-report-2020/.
-
[3]
Z. Kifle, Infect. Drug Resist. 13 (2020) 4047-4060. doi: 10.2147/IDR.S279433
-
[4]
D.E. Goldberg, A.F. Slater, A. Cerami, G.B. Henderson, Proc. Natl. Acad. Sci. U. S. A. 87 (1990) 2931-2935. doi: 10.1073/pnas.87.8.2931
-
[5]
J. Liu, E.S. Istvan, I.Y. Gluzman, J. Gross, D.E. Goldberg, Proc. Natl. Acad. Sci. U. S. A. 103 (2006) 8840-8845. doi: 10.1073/pnas.0601876103
-
[6]
S. McGowan, Curr. Opin. Struct. Biol. 23 (2013) 828-835. doi: 10.1016/j.sbi.2013.07.015
-
[7]
S.M. Marcin Poreba, T.S. Skinner-Adams, K.R. Trenholme, et al., PLoS One 7 (2012) e31938. doi: 10.1371/journal.pone.0031938
-
[8]
S. Dalal, M. Klemba, J. Biol. Chem. 282 (2007) 35978-35987. doi: 10.1074/jbc.M703643200
-
[9]
M.B. Harbut, G. Velmourougane, S. Dalal, et al., Proc. Natl. Acad. Sci. U. S. A. 108 (2011) E526-E534.
-
[10]
R. Deprez-Poulain, M. Flipo, C. Piveteau, et al., J. Med. Chem. 55 (2012) 10909-10917. doi: 10.1021/jm301506h
-
[11]
M. Flipo, T. Beghyn, V. Leroux, et al., J. Med. Chem. 50 (2007) 1322-1334. doi: 10.1021/jm061169b
-
[12]
N.B. Vinh, N. Drinkwater, T.R. Malcolm, et al., J. Med. Chem. 62 (2019) 622-640. doi: 10.1021/acs.jmedchem.8b01310
-
[13]
S.N. Mistry, N. Drinkwater, C. Ruggeri, et al., J. Med. Chem. 57 (2014) 9168-9183. doi: 10.1021/jm501323a
-
[14]
T.S. Skinner-Adams, C.L. Peatey, K. Anderson, et al., Antimicrob. Agents Chemother. 56 (2012) 3244-3249. doi: 10.1128/AAC.06245-11
-
[15]
N. Drinkwater, N.B. Vinh, S.N. Mistry, et al., Eur. J. Med. Chem. 110 (2016) 43-64. doi: 10.1016/j.ejmech.2016.01.015
-
[16]
C. Ruggeri, N. Drinkwater, K.K. Sivaraman, et al., PLoS One 10 (2015) e0138957. doi: 10.1371/journal.pone.0138957
-
[17]
K.K. Sivaraman, A. Paiardini, M. Sienczyk, et al., J. Med. Chem. 56 (2013) 5213-5217. doi: 10.1021/jm4005972
-
[18]
E. Cunningham, M. Drag, P. Kafarski, A. Bell, Antimicrob. Agents Chemother. 52 (2008) 3221-3228. doi: 10.1128/AAC.01327-07
-
[19]
T.S. Skinner-Adams, J. Lowther, F. Teuscher, et al., J. Med. Chem. 50 (2007) 6024-6031. doi: 10.1021/jm070733v
-
[20]
E. Salomon, M. Schmitt, E. Mouray, et al., Bioorg. Chem. 98 (2020) 103750. doi: 10.1016/j.bioorg.2020.103750
-
[21]
L. Bounaadja, M. Schmitt, S. Albrecht, et al., Malar. J. 16 (2017) 382. doi: 10.1186/s12936-017-2032-4
-
[22]
G. Velmourougane, M.B. Harbut, S. Dalal, et al., J. Med. Chem. 54 (2011) 1655-1666. doi: 10.1021/jm101227t
-
[23]
C. Ma, J. Cao, X. Liang, et al., Eur. J. Med. Chem. 108 (2016) 21-27. doi: 10.1016/j.ejmech.2015.11.021
-
[24]
C. Ma, K. Jin, J. Cao, et al., Bioorg. Med. Chem. 21 (2013) 1621-1627. doi: 10.1016/j.bmc.2013.01.068
-
[1]
-
Figure 2 The binding mode of compound 26 (green, PDB: 5XM7) in the active site of PfA-M1 (gray cartoon and sticks). The hydrogen bonds are indicated by black dashed lines. The zinc ion coordination is omitted for clarity.
Table 2. Design strategy of novel peptidomimetic hydroxamates 19–26 and their inhibition data against PfA-M1, PfA-M17 and pAPN.
 下载: 导出CSV
下载: 导出CSV
-






 扫一扫看文章
扫一扫看文章
计量
- PDF下载量: 13
- 文章访问数: 990
- HTML全文浏览量: 79
 下载:
下载:
