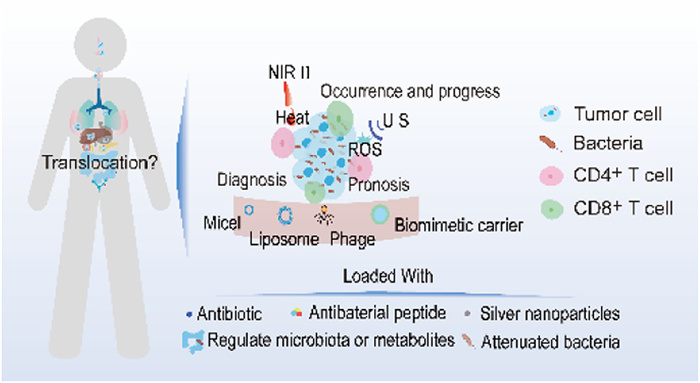
Figure 1.
The impact and treatment of intratumoral bacteria on tumors.
Recent progress on drug delivery systems of regulating intratumoral bacteria for tumor therapy
Jing Feng , Yanhong Liu , Liming Gong , Chenfei Liu , Congcong Xiao , Liqing Chen , Mingji Jin , Zhonggao Gao , Wei Huang , Yubo Li

The human microbiota exists on the surface of our skin and in our cavities. More and more studies have shown that the microbiota of healthy and diseased populations is different. Especially, bacteria in the tumor microenvironment have an inseparable relationship with the occurrence and development of cancer. Traditionally sterile tumor tissues have also shown unique microbial characteristics. Since 100 years ago, the presence of intratumoral microorganisms was been discovered, however, due to low content, the accuracy of detection was difficult to ensure. In recent years, researchers have studied and proposed solutions to the possible problems in the determination and provided analysis methods of intratumoral microorganisms, making the detection level of microorganisms more accurate and valuable, the presence of low-biomass bacteria in tumors has gradually been verified and accepted by the public which promoting the research progress of intratumoral bacteria [1-3]. During the same period, researchers measured the microorganisms in 1526 tumors and their adjacent tissues, they found that each tumor had its specific microbial types [4,5], due to the different bacteria having their own best survival conditions, there are differences in the type and abundance in different tumors of different systems, in addition, the microorganisms inside the tumors existed within the tumor cells [6]. Researchers have pointed out that microorganisms on the mucosa are an important source of microorganisms within tumors, while microorganisms in other parts of tumors are transferred and colonized from mucosa through blood and other methods [7]. Literature research has shown that the existence of microorganisms in tumors is closely related to the presence of microorganisms in the digestive tract, it has been confirmed that the transfer and colonization of bacteria through blood to other traditionally sterile tissues has also been confirmed [8]. Researchers have found that the pathogenic mechanisms of microorganisms within tumors include inducing inflammation, altering the cell cycle, causing rapid cell proliferation, DNA double-strand damage, or even breakage, thereby promoting tumor progression [9-13]. Not only that, tumor microorganisms also can lead to resistance by inhibiting the immune microenvironment in the tumor, reducing immune killing ability and affecting the stability of chemotherapy drugs [7,13,14]. Since tumor microorganisms are biomarkers for tumor development, there also be some correlation between tumor prognosis and diagnosis with tumor microorganisms. The types, abundance, metabolites and alpha-diversity of intratumoral bacteria can effectively predict and diagnose the level, size of tumors and patient survival [15-17]. There are many tumor treatments for intratumoral bacteria, except traditional antibiotic treatment, the nano delivery systems, near-infrared (NIR) thermogenesis, regulation of gut microbiota and the use of attenuated intratumoral bacteria or metabolites, all of these can achieve the goal of reducing drug resistance, increasing drug targeting, reducing systemic toxicity and increasing patient compliance [18-20]. So far, most clinical applications of intratumoral bacteria have focused on the diagnosis and prognosis of patients. It is believed that with the continuous advancement of scientific research, research on intratumoral bacteria can play an important role in the clinical treatment of cancer.
This review emphasizes the key role of intratumoral bacteria in occurrence and progression, including promoting tumor development, accelerating tumor metastasis and promoting tumor resistance. In addition, this article summarizes the application of intratumoral bacteria in tumor diagnosis and prognosis. Aiming to understand the role of intratumoral bacteria at various stages of tumors. Moreover, this review also summarizes the therapeutic strategies of intratumoral bacteria, including non-nanodelivery system and nanodelivery systems, aiming to provide current research ideas and help researchers to design better anti-tumor strategies. The highlights of this review are summarized in Fig. 1.
We already know that bacteria colonize in various types of tumors and there is a significant difference in bacteria diversity between healthy tissues and cancer cells [21-23]. We usually believe that sterile tissues, such as the breast and pancreas, the microbiota in these tissues may transferred from the gastrointestinal tract through biofilms, this is related to the permeability of the gastrointestinal mucosa [17,24,25].
Esophageal cancer is one of the eight common cancers in the world and the sixth leading cause of death [26]. Esophageal squamous cell carcinoma develops rapidly with poor prognosis and the incidence rate increases year by year [27].
After research, there was an increased abundance of Fusobacterium, Porphyromonas and Fusobacterium nucleatum (F. nucleatum) in patients with esophageal squamous cell carcinoma and the increase of this abundance had significant differences [28-30].
Colon cancer is the main cause of death related to gastrointestinal diseases [31,32]. Research indicated that the composition of intratumoral microbiota was different in colon cancer patients. Colorectal cancer was related to F. nucleatum and other microbiota, such as Faecalibaterium and other microbiota-colonized microniches. In these bacterial species, there were more F. nucleatum in colon cancer patients than the healthy people and there were less Faecalibaterium than healthy people [21,22].
There are some studies about liver cancer. The authors indicated that microbiota in tumor cells was associated with hepatocellular carcinoma. Patescibacteria, Proteobacteria, Bacteroidota, Firmicutes and Actinobacterita were enriched in hepatocellular carcinoma cells, the abundance of the intratumor bacteria was related to clinicopathological features. The authors also found that the intratumoral bacteria aggregation in the cytoplasm rather than the nucleus, was consistent with the results of other cancers [33,34]. Besides, the bacteria also existed around the tumor tissue, which indicated that the tissue around the tumor was similar to the tumor tissue [35]. Another article also came up with a similar conclusion, there was a significant difference in the intratumoral microbiota between the hepatocellular carcinoma tumor tissue and the normal tissue around the tumor, the microbiota includes Actinobacteria, Proteobacteria and Firmicutes, the diversity of bacteria in tumors was lower than that in normal liver tissue, these intratumoral bacteria may have migrated from the gut microbiota [17].
Pancreatic cancer is known as the "king of cancer" in the medical community because of its characteristics of occult onset, low diagnosis rate, rapid progress and poor prognosis [36]. There is a paper indicating that bacteria could transfer from the intestine to the pancreas and this phenomenon then lead to pancreatic cancer. Compared to healthy people and mice, bacteria increased about 3000 times than the normal pancreatic tissue and Malassezia spp. enriched in pancreatic cancer cells.
A. sydowii was a symbiotic bacterium in the lungs, due to the detection of Aspergillus nidulans (A. sydowii) in the oral cavity, A. sydowii likely transferred from the oral cavity to lung [37]. The bacterial load in lung cancer cells is higher than in immune cells and the microbiota load gradually decreases from the respiratory tract to surrounding tumors and then tumor cells, this indicates the transfer pathways of bacteria [38].
The infection and pathogenic bacteria of oral cancer and colon cancer are similar, the bacteria that can lead to cancer in the oral cavity also can colonize in the colon through the gastrointestinal tract, but there are > 600 bacterial types are endemic in oral squamous cell carcinoma, by contrast, there are not so many types of bacteria in colon cancer, this may be due to there are more opportunities for oral to contact with outside [39].
The study found there were 16.4 species of bacteria in breast cancer tissue on average while there were no more than 9 types of bacteria in other cancer cells [6].
The presence of microbiota in melanoma has been found in many studies [40,41]. Such as a study indicates that the intratumoral bacteria in melanoma affect the immune microenvironment of the tumor based on the different types of bacteria. Lachnoclostridium genus is positively correlated with the quantity of CD8+ T cells, Lachnoclostridium genus also can improve patients' survivability [42].
We already know that bacteria colonize in various types of tumors and there is a significant difference in bacteria diversity between healthy tissues and cancer cells [21,22,33,43]. We usually believe that microbiota in sterile tissues may transferred from the gastrointestinal tract through biofilms, this is related to the permeability of the gastrointestinal mucosa [17,24,25]. Bacteria inside tumors can affect the tumor immune microenvironment and inflammatory microenvironment [21,22,25,28-30,44-46]. Not only that, the intratumoral bacteria can associated with gene mutation and DNA double-strand breakage, which is the most serious species of DNA damage [25]. These factors can promote the occurrence and development of tumors. The summary of intratumoral bacteria promotes tumor occurrence is shown in Table S1 (Supporting information).
Intratumoral bacteria reshape the tumor's immune and inflammatory microenvironment is the main reason for promoting tumor occurrence and progress. Fusobacterium and Porphyromonas in the tumors of esophageal cancer patients could increase the level of programmed death receptor 1 (PD-1) which decreases the immune ability of body. Porphyromonas could stimulate Toll-like receptors (TLR) receptors in oral epithelial cells, activate the interleukin-6 (IL-6)/signal transducer and activator of transcription 3 (STAT3) pathway and promote the growth of squamous cell carcinoma [28,29]. Furthermore, Fusobacterium had a similar ability to promote the process of colon cancer and oral squamous cell carcinoma [21,22,44,45]. Another research indicates F. nucleatum in esophageal squamous cell carcinoma tumor cells could promote cancer progression. The reason was that it could reduce the immune response of PD-1 inhibitors and inhibit T cell proliferation, in addition, its toxicity factors Fusobacterium nucleatum DNA starvation/stable phase protection protein (Fn-Dps) could promote the survival of F. nucleatum in tumor cells [30].
Studies show that F. nucleatum in colorectal tumor cells could promote cancer progression, the reason was that tumor suppressor p53 downregulation, CD4+ and CD8+ T cells also downregulated in tumor tissue, at the same time, cytotoxic T-lymphocyte-associated antigen 4 (CTLA-4) and arginase 1 (ARG1) overexpressed in bacterial positive tumor tissue. Not only that, the infected tumor cell showed obvious regulation of signaling pathways including tumor necrosis factor-α (TNF-α), inflammation, hypoxia and the p53 pathway which could lead to inflammation and promote cancer progression [22].
In liver cancer, the family Streptococcaceae and genus Lactococcus were related to the hepatocellular carcinoma patient who had cirrhosis, the reason was that immune environment altered the function, metabolism and composition of microorganisms [35].
Similarly, intratumoral microbiota A. sydowii in lung cancer could activate and amplify myeloid-derived suppressor cells through IL-1β-mediated, the pathway is that β-glucan/dectin-1/caspase recruitment domain-containing protein 9 (CARD9). This phenomenon would lead to T lymphocyte cell activity being inhibited [37].
From the other references, it can be concluded that PD-1 overexpressed in Oral squamous cell carcinoma tumors and the bacteria also positive in this area, this phenomenon could lead to the T cell markers downregulation, such as CD3, CD8, CD4, CD27 and CD44, as a consequence, the bacteria in tumor tissue could inhibit the activity of T cells [22]. F. nucleatum could create an immune microenvironment which led to cancer cell deterioration, the reason was F. nucleatum could induce cancer cells to produce more lactic acid, which promoted the cancer process [44]. A similar study also found that bacteria Sphingomonas yanoikuyae in healthy breast tissue had a special ability to stimulate immune cells to maintain a normal breast immune environment. This result indicates that dysbacteriosis is related to breast cancer [47].
Human leukocyte antigen class Ⅰ (HLA-I) and HLA-Ⅱ peptides derived from bacteria within tumor cells could be presented by melanoma cells and recurrent peptides could trigger immune responses in tumors. Intratumoral microbiota in melanoma include Bifidobacterium longum, Collinsella aerofaciens and Enterococcus faecium. These intratumoral microbiota could affect the magnitude of spontaneous antitumor T cell responses of patients and animal models, it also could reduce the effectiveness of immunotherapy, if we use anti-PD-1 therapy in sterile mice, the T cell response will be enhanced [48,49].
Intratumoral bacteria can also alter the metabolism of host tumor cells, thereby promoting tumor development. A study found there was a significant difference in microbiota between hepatocellular carcinoma tissue and normal liver tissue, which resulted in differences in metabolic patterns between cancer tissue and normal liver tissue. Not only that, but it also altered the host's genetic transcription [50].
The special is that smoking-induced lung cancer patients have unique microbiota in their lung cancer cells, this type of tumor contains tumor protein 53 (TP53) mutations, but TP53 is a tumor suppressor gene in organisms, which means that the function of the inhibiting cancer has been damaged. Indicating that the intratumoral microbiota could associated with gene mutation [51].
The increase of anaerobic bacteria could also cause dysbiosis of the microbial community, thus inducing to inflammatory reactions, increasing mutation rate, altering immune microenvironment, which is related to the process of hepatocellular carcinoma, drug resistance and prognostic effects [25].
Bacteria in tumors is closely related to tumor metastasis [52-54]. Although the intracellular content of the bacteria is low, it also plays a crucial role in tumor metastasis [55]. A brief summary of the promotion of tumor metastasis by intratumoral bacteria is shown in Table S2 (Supporting information). Research has found that the exosomes produced by tumors can help the metastasis of bacteria in tumors [56]. In addition, intratumoral bacteria also can enhance the adhesion between tumor cells and epithelial cells, which is crucial for tumor growth and metastasis [57]. The metastasis of colon cancer cells is closely related to the permeability of the gastrointestinal tract, some bacteria can reduce the permeability of the gastrointestinal mucosa, which can help the metastasis of intratumoral bacteria [58]. In addition, toxins produced by intratumoral bacteria can assist in the metastasis of cancer cells. Of course, the metastasis of cancer cells is closely related to changes in the immune environment caused by intratumoral bacteria [22,59].
Bacteria exists in colon cancer cells could activate the transcription factor and create the same genetic characteristics of metastasis. The migration speed of colon cancer cells which infected by F. nucleatum was 15 times faster than the cancer cells not been infected by F. nucleatum. This conclusion indicated that F. nucleatum be could associated with colon cancer cell metastasis [22].
The exosomes produced by cancer tumors also could help the metastasis of colon cancer. F. nucleatum can promote the production of exosomes. The colon cells infected by F. nucleatum had higher level of miR-1246/92b-3p/27a-3p and C-X-C motif chemokine 16/Ras homolog family member A/IL-8 (CXCL16/RhoA/IL-8), the exosomes which promoted by F. nucleatum of colon cells would carry miR-1246/92b-3p/27a-3p-rich and CXCL16/RhoA/IL-8 to the colon cells which not been infected by F. nucleatum, furthermore, miRNAs transported by exosomes could help bacteria survive in cancer cells. This situation can increase the metastasis ability of colon cells higher than before [56].
F. nucleatum could increase the adhesion between colon cancer cells and epithelial cells to assist in tumor cell metastasis. F. nucleatum abundance had a positive correlation with abundance of alpha-protein kinase 1 (ALPK1) and intercellular adhesion molecule-1 (ICAM1). Through this mechanism, the level of ICAM1 increased and enhanced adhesion between colon cancer cells and epithelial cells, the adhesion between colon cancer cells and epithelial cells was the key step to cancer cell colonization, for these reasons, colon cancer cells metastasis ability will be enhanced [57].
Another report indicates that bacterial metastasis in colon cancer was related to the permeability of the gastrointestinal tract. The gut vascular barrier was injured by the virulence factor from Escherichia coli (E. coli) then the intratumoral bacteria could transfer to liver, the level of plasmalemma vesicle-associated protein 1 (PV-1) was a marker of intestinal damage, the metastasis of colon cancer to the liver was associated with an increase in the content of PV-1 which confirms the hypothesis of researchers [58].
The researchers found that bacteria in breast cancer also play a key role in metastasis. Due to the survival of bacteria within tumor cells, bacteria migrate with the metastasis of tumor cells during the metastasis process. The content of bacteria in metastatic issues is higher than in primary tumors, which means that metastatic tumor tissue is more suitable for bacterial growth. The intracellular bacteria can reshape the cell cytoskeleton and enhance cell vitality under the mechanical force of blood, which leads to the promotion of cell metastasis [8]. The metastasis path and mechanism of breast cancer showed in Fig. S1 (Supporting information).
Intratumoral bacteria can cause phenotypic changes in cancer cells and the metastasis of cancer cells is related to the toxins produced by bacteria. The breast cancer cells infected by Fragili have a mesenchymal-like phenotype which has a significant change in cell morphology, besides, this phenotype enhanced migration ability. The potential ability of stem cells has been discovered in breast cancer cells which are treated by Fragili toxins and stem cells are crucial for cell migration. The toxin produced by Fragili also can activate the Wnt-β-catenin pathway which is also related to the metastasis of breast cancer cells [60].
The metastasis of intratumoral bacteria is related to changes in the immune environment. Another report indicated that F. nucleatum-infected breast cancer cells could inhibit the accumulation of T cells containing CD4+ and CD8+ at the tumor site and promote the metastasis of breast cancer [59]. Intratumor bacteria in oral squamous cell carcinoma by recruiting bone marrow cells to increase the ability of cancer cells to metastasize, while recruiting bone marrow cells is a kind of cells that could promote the process of oral squamous cell carcinoma [22].
The mechanisms of drug resistance include genetic changes, DNA damage, dysregulation of cell apoptosis and changes in the tumor environment [61]. Drug resistance caused by intratumoral bacteria can lead to chemotherapy failure [62-64]. The drug resistance caused by intratumoral bacteria can be divided into the following reasons: Firstly, intratumoral bacteria can reshape the microenvironment of immune and oxygen environment [65,66]. Secondly, intratumoral bacteria can downregulate the efficacy of chemotherapy drugs, including some bacteria, such as E. coli can combine with chemotherapeutics, so that can increase the ability of pathogenic bacteria to promote cancer progression [67]. Thirdly, intratumoral bacteria also can affect the stability of chemotherapy drugs [68]. Last but not least, l-lactate produced by Lactobacillus can change signal pathway of l-lactate and metabolic pathways of tumors [69]. All of these reasons can lead to resistance of chemotherapy and radiation therapy, even shorten the lifetime of patients. A Brief Summary of intratumoral bacteria tumor that promotes tumor cell resistance is listed in Table S3 (Supporting information).
According to the reference, intratumoral bacteria is closely related to tumor resistance due to it can reshape the microenvironment of immune and oxygen environment. The literature suggests that resistance to colon cancer was related to five types of bacteria. These five kinds of bacteria could cause immune inhibition and hypoxia at the tumor site, which could promote the progress of tumor [65]. Pancreatic cancer is also related to the specific strains of intratumoral bacteria. The intratumoral bacteria can induce an immunosuppressive environment so that it can cause resistance to chemotherapy. When the researchers used antibiotics to kill intracellular bacteria, macrophages transitioned from M2-like to M1-like, CD4+ and CD8+ T cells infiltration significantly. This meant that the immune system was activated and drug resistance was relieved by antibiotics [66].
The intratumoral bacteria also could reduce the efficacy of chemotherapeutics. The experiment indicated that 5-fluorouracil lost 95% of its ability to inhibit the growth of colon cancer epithelial cells after 24 h exposure to E. coli, which increased the ability of F. nucleatum to promote the colon cancer process. Therefore, leading to the resistance of treatment [67]. Besides, the intratumoral bacteria also could affect the stability of chemotherapy drugs. Another study also pointed out that intratumoral bacteria Mycoplasma hyorhinis (M. hyorhinis) could lead to resistance of gemcitabine, the reason was that intratumoral M. hyorhinis could metabolize gemcitabine into metabolite 2′-2′-difluorodeoxyuridine without pharmacological activity [13].
The l-lactate produced by Lactobacillus could change the signal pathway of l-lactate and metabolic pathways of tumors. It was worth noting that l-lactate increases significantly in tumors after irradiation, therefore, cancer cells could utilize more l-lactate to promote the citric acid cycle, cancer cells consume a large amount of energy and rapidly proliferate, thus exhibiting drug resistance in cancer cells [69].
The species and the abundance of intratumoral bacteria can be biomarkers for diagnosis [51,70-73], besides, gene mutations in tumor cells caused by intratumoral bacteria can also serve as biomarkers for cancer diagnosis [51], last but not least, the labeled bacteria also can be used to analyze tumor metastasis trajectories [74]. A Brief application summary of intratumoral bacteria in tumor diagnosis is listed in Table S4 (Supporting information).
Study showed that the tumor bacteria and the bacteria in the bronchoalveolar fluid were closely related, bronchoalveolar fluid was easy to get and no-invasive, thus we could detective the abundance of bacteria in the bronchoalveolar fluid to evaluate the severity of non-small cell lung cancer [70]. The process of papillary thyroid cancer is related to the abundance of bacteria in tumor cells, the alpha-diversity of bacteria in late-stage patients is higher than early stage patients, besides, the researchers found that the eight tumor bacteria taxa can predict the stage of tumor progression, thus both the alpha-diversity and the species of intratumoral bacteria could be a biomarker for tumor progression [71]. Another study indicated that it is also related to bacteria DNA loaded in tumors. The bacteria DNA load in breast tumor tissue is lower than the healthy breast tissue, which indicates that the bacteria DNA load may become a diagnostic indicator of breast cancer. It is worth noting that intratumoral bacteria can cause DNA breakage [72]. The bacteria from breast cancer tissue were different from the healthy breast tissue, the bacteria of Bacillus, Enterobacteriaceae and Staphylococcus enriched in breast cancer tissue, the Enterobacteriaceae and Staphylococcus which isolated from cancer tissue could cause DNA double-strand breakage in HeLa cell [73].
The literature suggested that in the tumor with TP53 mutations, Acidovorax sp. was enriched in this kind of lung cancer, the loss of TP53 could damage epithelial cells and stimulate the pathway of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) resulting in the rapid progression of cancer, thus Acidovorax sp. could be a biomarker of early diagnosis [51].
Apart from using the types and abundance of bacteria we have known to evaluate tumor progression, the labeled bacteria can be used to analyze tumor metastasis trajectories. In this study, the researchers used green fluorescent protein (GFP)-Erm to decorate the Staphylococcus xylosus (S. xylosus) and track the metastasis trajectories of mouse mammary tumor virus promoter-polyoma virus middle T antigen (MMTV-PyMT) tumor-bearing mice. The labeled S. xylosus could colon in tumor cells and metastasis with tumor cells, so that researchers could track the trajectory of cancer cells, this method could detect early cancer metastasis [74].
Researchers found that intratumoral bacteria could affect the prognosis of patients with various types of cancer, the types of intratumoral bacteria could be a good method of predicting the patient's state, including the level, size, site of tumors and survival term of patients [15,17,45,75-79]. Many researchers use the type of tumor bacteria to divide the tumors into different subtypes, different subtypes and patients' disease-free survival, and the survival rate is significantly related. Meanwhile, probiotics could protect the tumor immune environment and inhibit tumor growth [75,80]. The application summary of intratumoral bacteria on prognosis is shown in Table S5 (Supporting information).
The study indicated that F. nucleatum and Granulicatella adiacens were associated with a positive correlation with disease-free survival. On the contrary, if patients had intratumoral bacteria containing Faecalibacterium prausnitzii and Ruminococcus gnavus, they would have shorter disease-free survival [81].
Different species of intratumoral bacteria are associated with different prognoses and clinical features of breast cancer, most of them are associated with a reshaped immune environment. For example, there is research found that Anaerococcus, Caulobacter and Streptococcus were the main bacteria in benign breast tissue and it did not exist in malignant breast tissue. The high abundance of Propionibacterium and Staphylococcus were related to a worse immune environment. The researchers found that the higher abundance was associated with lower survival, in addition, the increased DNA of F. nucleatum was significantly associated with poor prognosis in patients. The mechanism of that was the bacteria could stimulate the chemokines such as chemokine (C—C motif) ligand 20 (CCL20) which could promote the process of tumor development and metastasis [82], CCL20 also could be a hinder of Treg lymphocytes migration, and the accumulation of Treg lymphocytes was significantly correlated with the patient's survival [83,84]. Besides, some researchers also found that A. baumannii, P. namnetense and P. stutzeri were related to poorer prognosis, which could be a biomarker of prognosis [34]. The different types of bacteria are also associated with the patient's mesenchymal traits subpopulation and immune microenvironment. The colon cells with a high abundance of Fusobacteriales and the patients with mesenchymal traits subpopulation would have poorer prognosis. The paper indicated that Fusobacterium can induce an activation of the NF-κB-TNF-α-IL-6 state and then activate the inflammatory factor, inhibiting the immune environment. Finally, it results in the proliferation of colon cancer cells [85]. Intratumoral microbiota can also affect the prognosis of liver cancer. The patients with a higher abundance of Akkermansia and Methylobacterium would have better overall survival and recurrence-free survival, on the contrary, the patients with lower alpha-diversity and higher abundance of Proteobacteria and Actinobacteria would have poorer prognosis. The mechanism was that the unbalance of tumor microorganisms can affect the tumor microenvironment. In addition, patients with Proteobacteria in gut would have shorter survival terms and a high risk of cancer malignancy, because the bacteria could colonize the liver via the pathway of the gut-liver axis [17]. The reason for this phenomenon was that the intratumoral microbiota may lead to a proinflammatory microenvironment, such as the increased level of CD8+ T cells in tumors, but the mechanism of this effect should be certified in the near future [15]. There is a paper indicating that the patients with high F. nucleatum load would have longer overall survival and disease-free survival, thus F. nucleatum load could be a predictor of anal squamous cell carcinoma prognosis. In this study, the researchers found that F. nucleatum could lead to the reshaping of the immune microenvironment, such as the lower level of M2 macrophages, CD4 lymphocytes, and ligand of TLR4, OX40, which indicated that F. nucleatum could result in a good prognosis. So the level of F. nucleatum could be a good marker of prognosis [77]. The major bacteria in nasopharyngeal carcinoma tumor cells are Corynebacterium and Staphylococcus. The major bacteria in nasopharyngeal carcinoma come from the nasopharynx and a portion comes from the mouth and intestines. The patients with a higher abundance of intratumoral bacteria load would have shorter disease-free survival, the reason was that it could inhibit the immune environment and reduce the T-lymphocyte infiltration which was consistent with previous research, besides, the patients who had suffered relapses also had higher alpha-diversity [76]. There are also some opposite examples, the F. nucleatum is related to the prognosis of oral squamous cell carcinoma. The reason is that F. nucleatum can reshape the local immune environment in tumors. The tumor with F. nucleatum infected is always related to higher age, drinking, smoking and less lymph node metastasis. The patients who are infected by F. nucleatum will have a lower probability of recurrence than the patients who have not been infected by F. nucleatum. In addition, the tumor which infected by F. nucleatum has lower M2 macrophages than the tumor that is not infected by F. nucleatum. These consequences do not conform with the former laws of pathogenesis and metastasis [45]. On the other hand, the high abundance of Streptococcus and Propionibacterium were related to more active T cells. In addition, the metabolites produced by intratumoral bacteria were also associated with the immune environment, for instance, cadaverine produced by Streptococcus genus could inhibit the process of tumor development and the transition of epithelial to mesenchymal [75]. Researchers analyzed the different intratumoral microbiota species in patients with long-term and short-term survival, the patients with long-term survival have a higher intratumoral microbiota alpha-diversity, the patients with Pseudoxanthomonas, Streptomyces, Saccharopolyspora, Bacillus clausii in tumor would have longer survival term [15].
On the contrary, some probiotics can slow down the progression of cancer, such as Leptotrichia, if the abundance of Leptotrichia is over the median, the patients will have a higher survival rate. The abundance of Leptotrichia in early patients is higher than that in late-stage patients, which indicates that it can protect the microenvironment and slow down the progression of cancer [68].
There was another report also indicated that the high alpha diversity of intratumoral microbiota had a significant correlation with survival. Proteobacteria, Firmicutes and Bacteroidetes were the main microbiota in long-term survival patients and Sphingomonas, Megasphaera were also enriched in tumor cells. The reason for this phenomenon was that intratumoral microbiota could reshape the metabolic pathway of tumors. Some bacteria also could produce metabolites that could inhibit tumor growth, such as Megasphaera can produce butyrate which has the ability to inhibit tumor growth [80].
The intratumoral bacteria of adrenocortical carcinoma is associated with the overall survival of patients. The regulatory mechanism is the genetic mutations in adrenocortical carcinoma tumor cells, including the upregulation of cell cycle and p53 signatures pathway are caused by the intratumoral bacteria and it can lead to a bad prognosis. The combination of the clinical stage of adrenocortical carcinoma and intratumoral bacteria can better predict the prognosis of patients [79].
Some researchers found that 107 genera signatures could affect lipid metabolism, which could lead to improvement of fatty acid metabolism and tumor cell cycle or division. However, Klebsiella has a higher abundance in patients with good prognosis. The improvement of fatty acid metabolism was enriched in better prognosis patients. In this literature, the ability of intratumoral bacteria on prognosis exceeded the clinical staging and patient age patients [16].
Researchers classified colon cancer into three species based on the strain of intratumoral bacteria, survival rate, and mutation signatures. Among the patients of stages Ⅱ, the intratumoral bacteria contained Fusobacterium, Escherichia, Pseudescherichia, and Shigella were related to lower survival rates. This would provide a criterion for judging the prognosis of colon cancer, besides, if we delivered drugs targeting pathogenic bacteria, such as metronidazole or bacteriophages, survival rates of patients could increase significantly [86]. Another report provided a similar consequence, they divided breast cancer into four species which were based on different intratumoral bacteria, different species of breast cancer have different prognoses. The level, size and the site of the tumor were closely related to the subtypes of breast cancer. The gram-negative bacteria had been detected in breast cancer subtypes, especially Fusobacterium had been detected with high abundance, which indicated that these subtypes would have a worse prognosis [87]. Ovarian cancer is known as a "silent killer" because it is often diagnosed in the late stage and the onset characteristics of ovarian cancer are blurry [88,89]. Recent studies have shown the bacteria in ovarian tumor cells conclude five species, Acinetobacter seifertii could inhibit the transfer of macrophages, which meant that many intracellular bacteria could not be engulfed by macrophages, leading to rapid cancer progression. This study also found that ovarian cancer could be divided into two types based on different types of bacteria: immune enriched and immune deficient types. Immune enriched type had a better prognosis [78].
There are many treatments for intratumoral bacteria, such as antibiotic therapy, macromolecular therapy, inflammatory factor inhibitor therapy, NIR-Ⅱ photothermal therapy, inorganic antibacterial agents, reactive oxygen species (ROS) therapy and microbes therapy. Microbes therapy include regulating gut microbiota, probiotic secretions, bionic carriers and phage. It is interesting that we also can use bacteriophages and attenuated intratumoral bacteria or extracts of intratumoral bacteria to increase the targeting of chemotherapy drugs and the permeability of tumor cells, thereby helping to improve the efficacy of chemotherapy drugs [20,90]. In terms of mechanism, besides reducing chemotherapy drug resistance and increasing the ability of an immune system, it also can increase the permeability of drugs in tumor cells, bacteria DNA damage and disrupt the oxygen environment in tumors, thereby achieving the goal of sterilization and inhibiting tumor growth [13,91-94]. The summary of the treatment is shown in Fig. 2.
All of these treatments can be divided into two categories, nanodelivery system and non-nanodelivery system. Compared with these two treatment methods, the treatment of non-nanodelivery systems due to lacking target, the efficiency dose is higher than nanodelivery system, so it will produce systemic toxicity. The long-term use of non-nanodelivery system will promote drug resistance. Not only that, the poor water solution of non-nanodelivery system also limited the dosage, obstructing the absorption of drug, thereby reducing biological utilization. Especially, The use of antibiotics can easily lead to drug resistance, while nanodelivery system can effectively alleviate or even solve this problem [18,90-92,95,96].
The summary of the Non-nanodelivery system strategies is listed in Table S6 (Supporting information).
Although non-nanodelivery systems can be easily used, it often exists systemic toxicity, low efficacy, resistance and poor stability in vivo.
The discovery of antibiotics is known as one of the greatest discoveries of the last century [97]. Antibiotics can kill bacteria inside tumors, reducing drug resistance [90], increasing the ability of the immune system [98]. In colon cancer, intratumoral bacteria Gammaproteobacteria can lead to the resistance of gemcitabine depending on secret cytidine deaminase, so the researchers used antibiotic ciprofloxacin to kill the bacteria and enhance the antitumor effect of gemcitabine [13]. Another study also used antibiotics to increase the ability of the immune system. The succinic acid was produced by F. nucleatum which could inhibit the transport of CD8+ T cells, the antibiotic metronidazole could kill the F. nucleatum and enhance the ability of the immune system, thus anti-PD-1 monoclonal antibody could exert anti-tumor effects [98].
In recent years, the development of engineered living materials has received increasing attention. In particular, the inherent targeting and colonization capabilities of bacteria play an important role in maintaining human health. For example, bacteria and their metabolites can stimulate the immune system, reshape the immune microenvironment, thus improving the efficacy of immunotherapy [99-101], in addition, bacterial therapy also has the effect of anti-tumor metastasis [102-104].
(1) Intestinal microbiota
The gut microbiota plays a crucial role in cancer treatment, the changes in the gut microbiota can reshape the immune environment and reduce tumor resistance. Clinical trials have shown that the transplantation or uptake of microorganisms can improve the success rate of treatment in patients [96], Mechanisms used by gut microbiota to modulate anticancer drug efficacy are shown in Fig. S2 (Supporting information). The intestinal microbiota plays an important role in the process of cancer, we also can use intestinal microbiota to slow down the development of cancer. Recently, a study used Ruminococcus gnavus and Blautia producta injection to inhibit the process of the tumor, in terms of mechanism, Ruminococcus gnavus and Blautia producta could degrade lyso-glycerophospholipids, lyso-glycerophospholipids was a compound which could inhibit T cell immune function, thus Ruminococcus gnavus and Blautia producta can activate the immune microenvironment [105]. There is another study that also applies the principle of gut microbiota regulate the immune environment, they found that Streptococcus could significantly increase the level of CD8+ and granzyme B+ T cells thus increasing the response of anti-PD-1 immunotherapy in esophageal squamous cell carcinoma [106]. Furthermore, the researchers found that Sachharopolyspora, Pseudoxanthomonas and Streptomyces are enriched in long survival term of pancreatic cancer patients, the special bacteria of tumor led to different immune microenvironment, such as an increase the level of CD8+ T cells in tumor environment [15].
(2) Probiotic secretions
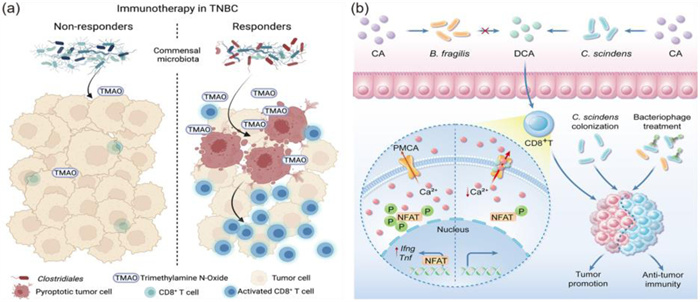
Recent studies have found that bacterial metabolites can affect the progression of cancer. Based on the data available in this case, we can speculate whether the metabolites of the microbial community can stimulate the immune system and improve the therapeutic effect of chemotherapy drugs [107]. There are some articles proposed viewpoints, such as indicating that many strains of Lachnospiraceae family could metabolize sugars to produce butyrate [108], Clostridium butyricum has positive effects for a range of human diseases, such as gut intestinal injury, inflammatory bowel disease and colorectal cancer [109], this was another reason that some microbiotas can be beneficial to the intestines and inhibited the tumor progress. Recently, researchers have used indole-3-aldehyde which is secreted by Lactobacillus reuteri to increase the effect of immune checkpoint inhibitors. The mechanism was that indole-3-aldehyde could stimulate the production of interferon-γ-producing CD8+ T cells, in addition, indole-3-aldehyde was a necessary condition for driving anti-tumor immunity [110]. There is a similar study that researchers used the trimethylamine N-oxide produced by Clostridiales which could stimulate pyroptosis in tumor cells by activating the endoplasmic reticulum stress kinase (PERK), then this process led to cell pyroptosis and achieved the goal of increasing the level of CD8+ T cells, reshaping the immune environment and increasing the efficacy of immunotherapy [111]. The mechanism of microbial metabolite trimethylamine N-oxide promotes antitumor immunity in triple-negative breast cancer in Fig. 2a. In addition, the researchers also used indole-3-acetic acid produced by B. fragilis and B. thetaiotaomicron to increase the effect of multiple chemotherapy drugs, including 5-fluorouracil, irinotecan, and oxaliplatin. The oxide of indole-3-acetic acid combined with 5-fluorouracil, irinotecan and oxaliplatin could lead to ROS accumulation and downregulation of autophagy in tumor cells, which could prevent the proliferation of cancer cells [112].
(3) Phage
The concentration of deoxycholic acid in patients with colorectal cancer is very high, the concentration of deoxymal acid is inversely proportional with the killing function of CD8+ T cells. Specifically, deoxymal acid can promote Ca2+, thereby suppressing the signal transmission of CD8+ T cells, eventually inhibiting the activation of CD8+ T cells. Deoxycholic acid is produced by Clostridium scindens (C. scindens), the researchers use phage to target C. scindens, effectively inhibit the concentration of deoxycholic acid, thereby stimulating the immune micro-environment, suppressing the growth of tumors ultimately [113]. The mechanism of bile acids which modified by the intestinal microbiota promote colorectal cancer growth is shown in Fig. 2b.
(4) Bionic carriers
Because the 5-fluoropromimidine will lose 95% of the efficacy under the exposure of the intestinal microorganism, intratumoral bacteria are resistant to 5-fluoropromimidine, therefore, it can protect the growth of intratumoral bacteria. The 5-fluoropromimidine decorated with intestinal microorganism isolates can improve the stability of 5-fluoropromimidine and reduce resistance. The researchers use E. coli isolates to modify 5-fluoropromimidine, achieving the goal of reducing the epithelial cell toxicity of 5-fluoropromimidine, increasing biological utilization, reducing the problem of chemotherapy resistance [67].
The most serious problem with non-nanodelivery systems treatments is the toxicity caused by systemic distribution in the whole body [114]. Due to the lack of targeting, it can cause harm to probiotics, thus leading to an imbalance in the body microbiota.
Nanodelivery systems have the following three advantages. Firstly, the nanodelivery system has strong targeting ability. Generally speaking, tumor drugs and bactericidal drugs have problems such as high toxicity and long-term use of drug resistance [115-117]. Nano-delivery systems loaded with anti-tumor drugs and anti-bacteria drugs to lesions, increasing the concentration of drugs at the tumor site and improving concentration dependence. The nanodelivery system also reducing the distribution of drugs in normal tissues, at the same time reducing toxicity in whole body, thus improving patients compliance [18]. For instance, natural products-derived nanomedicine has great potential in this regard [118]. Secondly, nanodelivery system have ability to prolong the circulation time of drugs in the body. Nanodrug delivery systems can extend the half-life of drugs by encapsulating drugs, which can significantly improve the bactericidal effect of time-dependent antibiotics [119,120]. Last but not least, it also can reshape the tumor microenvironment. Since intratumoral bacteria can suppress the tumor immune microenvironment, targeted killing of intratumoral bacteria can activate the immune microenvironment and increase immune cell infiltration in the tumors. However, it also has disadvantages, such as it is difficult to expand production and lower stability than expected. The summary of the nanodelivery system strategies is listed in Table S7 (Supporting information).
Here are some articles about nanodelivery of antibiotics. For instance, there is a report linking the antibiotics metronidazole and fluorouridine by disulfide bond, forming amphiphilic molecules metronidazole-fluorouridine, thus nano micelles could be formed in hydrophilic solutions, the high level of glutathione (GSH) in the tumor environment could lead to disulfide bond cleavage, targeting release of metronidazole and fluorouracil in the tumor environment, reshaping the tumor microenvironment and reducing irritation to the gastrointestinal tract [18]. It is worth noting that silver ions is a broad-spectrum antibacterial drug [139,140]. There is a research used silver-tinidazole complex loaded in liposome to clear up the intratumoral bacteria of in situ and liver metastases, the treatment mechanism was that the liposome could stimulate the immune environment and increase the CD8+ T cells, this liposome could increase the long-term survival rate exceeded 70% [14]. In addition, the outer membrane of intratumoral bacteria could be used to stimulate the immune system and improve the targeting effect. Due to the high affinity between protein fibroblast activation protein 2 (Fap-2) of F. nucleatum and D-galactose-β(1–3)-N-acetyl-D-galactosamine, the researchers used the membrane of F. nucleatum to modify liposomes, which liposome was colistin loaded. So this liposome could target colon cancer cells actively [121]. There are other studies which also used metronidazole as the antibiotic, the difference was that the outer membrane of bacteria was coated on the surface of nanoparticles, which can target tumor cells and it also could promote the maturation of dendritic and T cell infiltration [122]. Furthermore, the study use colistin crosslinked gemcitabine micelle and it can released by GSH in tumor microenvironment [123]. Last but not least, another research designed a nanoassembly loaded with lauric acid and oxaliplatin prodrug and the nanoassembly can release drug in an acidic tumor microenvironment [124].
Antibacterial peptides are a kind of peptide composed of 15–50 amino acid residues, so antibacterial peptides are a macromolecular. Antibiotics are facing severe challenges due to the issue of drug resistance. While the advantages of antibacterial peptides include lack of resistance, low toxicity and high solubility [95]. Research designed mesoporous silica nanoparticles loaded with doxorubicin and antibacterial peptide HHC36 (seq: Lys-Arg-Trp-Trp-Lys-Trp-Trp-Arg-Arg), the drug release relay on triggered by GSH and stimulates the immune system which is inhibited by Staphylococcus aureus and Streptococcus pneumoniae bacteria [125]. There is another study also used antibacterial peptides linked to photosensitizer via poly ethylene glycol (PEG) chain, it could form micelles with a diameter of 20 nm. Both antibacterial peptides and photosensitizers can produce antibacterial effects and reduce the resistance [126]. Another research found that lipopolysaccharide (LPS) which from Gram-negative bacteria can downregulate the immune microenvironment, so the study designed an LPS-targeting fusion protein and loaded it with nanoparticle system, this nanodelivery system significantly relieved the immunosuppressive microenvironment and even attenuated colorectal cancer (CRC) liver metastasis [127].
Anti-TNF-α can effectively control the patient's inflammation, it takes effect quickly and has good tolerance. So the researchers can use anti-TNF-α to combine with the intratumoral bacteria and reduce the local inflammation, anti-TNF-α prevents bacterial colonization in the lesion [91]. The researchers used anti-TNF-α loaded in Nb2C/Au nanocomposite to reshape the immune and metabolic environment and then remove the tumor resistance. This method has achieved significant anti-tumor effects [128].
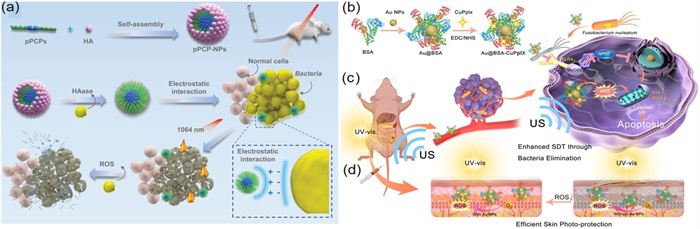
The researchers use NIR-Ⅱ photothermal which can kill the bacteria, the NIR-Ⅱ photothermal can generate mild heat and interfere with cell membrane permeability, even causing DNA damage to bacteria. The researchers designed a structure with photothermal and it could be degraded by oxygen species around infected tissue, so the safety could be guaranteed, the material could form nanoparticles with hyaluronic acid (HA) and modified by quaternary phosphonium cations, thus it could target bacterial membranes. Under infrared light irradiation, the nanoparticles could continuously kill intracellular bacteria and the nanoparticles display 100% sterilization efficiency in vitro [129]. Schematic illustration showing the design of phosphonium-based pseudo conjugated polymers nanoparticles (pPCP-NPs) for antibacterial therapy was shown in Fig. 3a. Similarly, there is a study that designed a nanoparticle loaded with gemcitabine and photothermal agent IR1048, under laser irradiation, the heat IR1048 created eliminated bacteria inside pancreatic cancer cells [130].
The number of deaths caused by antibiotic resistance worldwide is 700,000 every year and this number will continue to rise, so we urgently need to develop a new sterilization method to replace antibiotics. Ag nanoparticles is a kind of inorganic metal, it has a strong bactericidal ability due to their small particle size and large specific surface area. Mechanistically speaking, it is because Ag nanoparticles can produce ROS and can damage membranes [92]. The study used mesoporous silica nanoparticles loaded with Ag nanoparticles (Ag NPs) and the anticancer drug epirubicin, killing bacteria while regulating cellular autophagy pathways to achieve the goal of reducing cell resistance [131].
ROS plays a crucial role in bacterial death, the mechanism of antibiotic-induced bacterial cleaning is the production of ROS [93,94]. Recently, a research designed a nanoparticle that could produce ROS after ultrasound and ROS could eliminate F. nucleatum in tumor cells, reducing the level of cell apoptosis inhibitory proteins, thereby inducing cell apoptosis, at the same time, it also could inhibit lung metastasis [132], the synthesis and therapeutic mechanism of nanoparticles was shown in Figs. 3b–d. Similarly, another researcher also designed a protein-supported nanozyme that could produce ROS and consume GSH, inducing unbalance of intracellular redox reactions and apoptosis of cancer cells, in addition, ROS could remove F. nucleatum and reduce the drug resistance of colon cancer cells [133].
(1) Phage
In the traditional sense, bacteriophages are the only factors that affect host infection. More and more studies have shown that using bacteriophages as carriers for drug delivery has better delivery efficiency and safety [20]. There is a study which researchers used the phage linked with irinotecan-loaded dextran nanoparticles, inhibiting growth of F. nucleatum and increasing the efficacy of chemotherapy drugs [134].
(2) Bionic carriers
Due to the unique accumulation characteristics of bacteria within tumors, bacteria can serve as a unique carrier for anti-cancer drugs. Bacteria also can combine with anti-cancer drugs in various ways to form targeted delivery drugs, which help anti-tumor drugs penetrate the interior of tumors, being absorbed by tumor cells, increasing drug concentration at the lesion site and thus increasing anti-tumor effects [90,118]. Salmonella typhimurium (S. typhimurium) can target and penetrate the interior of tumors, so the researchers used attenuated S. typhimurium carry liposomes loaded with cancer inhibitors, this method improved the targeting and effectiveness of chemotherapy drugs [135]. Another similar study used bacteria which decorated with bacteria lipoteichoic acid antibodies, so the bacteria could accurately target bacteria within tumors and deliver chemotherapy drugs. The effectiveness of this treatment has been validated in a model of colon cancer, lung cancer, and breast cancer [136]. The researchers used F. nucleatum membrane to grained nanoparticles and then target F. nucleatum. This drug system can reshape the micro-environment of the tumor, enhance the therapeutic effect of the programmed death 1 (PD-L1) loaded by nanoparticles and extend the survival of the tumor-bearing mice [137]. In addition, another research designed a capsular polysaccharide enfolded camouflaged gallium-polyphenol metal-organic network, the Ga3+ released from the net could disrupt bacteria iron respiration and the capsular polysaccharide could extend the half-life of nanoparticles [138].
Intratumoral bacteria as an inducer of cancer, a biomarker of the entire cancer process and a criterion for diagnosis and prognosis [15,21,22,71], has received increasing attention from researchers in recent years. It is generally believed that aseptic tumor tissues, such as breast cancer, pancreatic cancer, liver cancer and lung cancer tissues would not be contact with the gastrointestinal tract and can still detect the existence of microorganisms in the tumor. Although there is a literature reports that microorganisms in these tissues may be transmitted and colonized through blood [8], but the detection method of microorganisms still needs to be improved, so we can determine the way of metastasis and colonization of tumor cells, which helps us prevent the metastasis of microorganisms in the tumor and further reduce the risk of tumor metastasis. Current literature indicates that the role of intratumoral bacteria in the occurrence and development of tumors mainly includes promoting tumor occurrence and metastasis, as well as promoting drug resistance in tumor cells. The mechanisms mainly include intratumoral bacteria remold the tumor microenvironment, altering the metabolic pathways of tumor cells, inducing gene mutations in tumor cells [30,50,51,139]. In addition, intratumoral bacteria can increase the adhesion of tumor cells, increase the permeability of the gastrointestinal mucosa, reshape the tumor cell cytoskeleton, and increase the vitality of tumor cells [57,58].
The types and applications of intratumoral bacteria in diagnosis and prognosis are also very extensive. At present, the clinical application of intratumoral microbiota is mainly in the field of diagnosis and prognosis. Specifically, diagnosing the types and abundance of intratumoral bacteria can be used as a biomarker for tumor diagnosis. In addition, the mutation of host genes caused by intratumoral bacteria can also be used as a biomarker for tumor diagnosis [51,70-73]. In clinical applications of patient prognosis field, the types of bacteria in tumors can predict the level of tumors and patients' survival [15,45,75-79]. In addition, the types of intratumoral bacteria can divide tumors into different subtypes, different tumor subtypes are significantly correlated with the patients' disease-free survival and survival rate [78,86,87]. Moreover, with the advancement of analytical methods of bacteria, the types of bacteria in tumors will continue to be studied and identified. Therefore, strategies can be designed according to the specific bacterial species in the tumor, so as to carry out more targeted and effective tumor treatment.
The traditional systemic use of antibiotics in the treatment of intratumoral bacteria has the potential problem of drug resistance. To address this issue, we use nanodelivery system to reduce side effects, such as antibiotic, macromolecular, inflammatory factor inhibitors, NIR-photothermal therapy, inorganic antibacterial agents, reactive species and microbes therapy [18-20,140]. The researchers can load antibiotics and various therapeutic drugs into nanoparticles for combination therapy, for instance, the combination therapy of antibiotics and chemotherapy drugs or the combination therapy of Ag nanoparticles and PD-1 inhibitors can be loaded in the nanodelivery system. We also can use the targeted colonization ability of tumor microbiota to modify nanoparticles and then achieve the goal of improving targeting ability, avoiding antibiotic resistance, reducing the toxicity of systemic medication, increasing the drug concentration of lesions, improving patient compliance and achieving good therapeutic effects. However, at the same time, we should pay attention to issues such as the scalability and industrialization of the formulation process, only the drug that can be used on patients can play value.
In this constantly developing new field, there are still many problems waiting to be solved. There are five main difficulties in the research field of intratumoral bacteria. Firstly, the interaction mechanism between intratumoral bacteria and tumors is not significant. In the future research process, the increasingly clear mechanism will help researchers design more targeted treatment strategies. Secondly, the sensitivity of the analysis method of intratumoral bacteria does not meet with the requirements. Due to the low content of intratumoral bacteria, resulting in high uncertainty in the species of bacteria. In the future research process, researchers in the analysis field will be able to improve the detection sensitivity and solve this problem. Thirdly, it is difficult to ensure aseptic conditions in the intratumoral microbiota experiment. Since it is difficult to ensure aseptic conditions during the sampling, transfer, and determination process, there is a high possibility of environmental contamination, the accuracy of the intratumoral microbiota detection results is difficult to ensure. In addition, safety issues in the use of antibiotics. Overuse of antibiotics can lead to drug resistance. In addition, broad-spectrum antibiotics have a wide range of bactericidal activity and may kill beneficial bacteria. For example, in the treatment of colorectal cancer, beneficial bacteria in the intestine should be protected. Therefore, targeted delivery of antibiotics is a focus to improve the safety of antibiotic use. Last but not least, the heterogeneity of intratumoral bacteria in different tumors increases the difficulty of treatment. Since the types of intratumoral bacteria in different tumors are different and the antibacterial spectra of antibiotics are different, treatment must be adopted for tumors when designing the strategies.
In future research, we should start from the above five aspects to get a deeper understanding of intratumor bacteria and develop personalized treatment plans for patients.
The authors declare no competing interests. It has not been published elsewhere and that it has not been submitted simultaneously for publication elsewhere.
Jing Feng: Writing – review & editing, Writing – original draft, Investigation, Funding acquisition, Formal analysis, Data curation, Conceptualization. Yanhong Liu: Formal analysis. Liming Gong: Formal analysis. Chenfei Liu: Resources. Congcong Xiao: Resources. Liqing Chen: Resources. Mingji Jin: Validation. Zhonggao Gao: Validation, Conceptualization. Wei Huang: Writing – review & editing, Funding acquisition, Formal analysis, Conceptualization. Yubo Li: Software.
This work was supported by CAMS Innovation Fund for Medical Sciences (CIFMS), China (No. 2021-I2M-1–026).
Supplementary material associated with this article can be found, in the online version, at doi:
S.P. Walker, M. Tangney, M.J. Claesson, Front. Oncol. 10 (2020) 179. doi: 10.3389/fonc.2020.00179
C. Xue, Q. Chu, Q. Zheng, et al., Cell Rep. Med. 4 (2023) 100884. doi: 10.1016/j.xcrm.2022.100884
C.J.F. Heymann, J.M. Bard, M.F. Heymann, et al., Cancer Lett. 522 (2021) 63–79. doi: 10.1016/j.canlet.2021.09.009
Y. Fu, J. Li, W. Cai, et al., Acta Pharm. Sin. B 14 (2024) 1560–1591. doi: 10.1016/j.apsb.2023.11.018
L. Yang, A. Li, Y. Wang, Y. Zhang, Signal Transduct. Target. Ther. 8 (2023) 35. doi: 10.1038/s41392-022-01304-4
D. Nejman, G. Fuks, N. Gavert, et al., Science 368 (2020) 973–980. doi: 10.1126/science.aay9189
Y. Xie, F. Xie, X. Zhou, et al., Adv. Sci. 9 (2022) 2200470. doi: 10.1002/advs.202200470
A. Fu, B. Yao, T. Dong, et al., Cell 185 (2022) 1356–1372. doi: 10.1016/j.cell.2022.02.027
M.R. Wilson, Y. Jiang, P.W. Villalta, et al., Science 363 (2019) eaar7785. doi: 10.1126/science.aar7785
Z. He, R.Z. Gharaibeh, R.C. Newsome, et al., Gut 68 (2019) 289–300. doi: 10.1136/gutjnl-2018-317200
Mara R. Rubinstein, X. Wang, W. Liu, et al., Cell Host Microbe. 14 (2013) 195–206. doi: 10.1016/j.chom.2013.07.012
M.T. Abreu, R.M. Peek, Gastroenterology 146 (2014) 1534–1546. doi: 10.1053/j.gastro.2014.01.001
G. Leore T, B.R. Michal, D. Tal, et al., Cancer 357 (2017) 1156–1160.
M. Wang, B. Rousseau, K. Qiu, et al., Nat. Biotechnol. 42 (2023) 1263–1274.
E. Riquelme, Y. Zhang, L. Zhang, et al., Cell 178 (2019) 795–806. doi: 10.1016/j.cell.2019.07.008
F. Pentimalli, M. Krstic-Demonacos, C. Costa, et al., Front. Immunol. 14 (2023) 1129513. doi: 10.3389/fimmu.2023.1129513
L. Sun, X. Ke, A. Guan, et al., Clin. Transl. Med. 13 (2023) 1263–1274. doi: 10.1007/978-981-19-2635-8_90
C. Gao, X. Wang, B. Yang, et al., ACS Nano 17 (2023) 7335–7351. doi: 10.1021/acsnano.2c11305
Y. Goto, S. Iwata, M. Miyahara, E. Miyako, Adv. Sci. 10 (2023) 2301679. doi: 10.1002/advs.202301679
G. Petrov, M. Dymova, V. Richter, Int J. Mol Sci. 23 (2022) 14245. doi: 10.3390/ijms232214245
H. Miao, N. Wu, C. Luan, et al., Acta Microbiol. Sin. 54(10) (2014) 1228–1234.
J.L. Galeano Niño, H. Wu, K.D. LaCourse, et al., Nature 611 (2022) 810–817. doi: 10.1038/s41586-022-05435-0
B. Roje, B. Zhang, E. Mastrorilli, et al., Nature 632 (2024) 1137–1144. doi: 10.1038/s41586-024-07754-w
E.M. Stoffel, R.E. Brand, M. Goggins, Gastroenterology 164 (2023) 752–765. doi: 10.1053/j.gastro.2023.02.012
J. Yang, Q. He, F. Lu, et al., Gut Microbes 15 (2023) 2201159. doi: 10.1080/19490976.2023.2201159
G. Murphy, V. McCormack, B. Abedi-Ardekani, et al., Ann. Oncol. 28 (2017) 2086–2093. doi: 10.1093/annonc/mdx279
H. Liang, J. Fan, Y. Qiao, Cancer Biol. Med. 1 (2016) 33.
Y. Masugi, R. Nishihara, J. Yang, et al., Gut 66 (2017) 1463–1473. doi: 10.1136/gutjnl-2016-311421
K. Mima, Y. Sukawa, R. Nishihara, et al., JAMA Oncol. 1 (2015) 653–661. doi: 10.1001/jamaoncol.2015.1377
Y. Li, S. Xing, F. Chen, et al., Nat. Commun. 14 (2023) 5788. doi: 10.1038/s41467-023-40987-3
J.J. Granados-Romero, A.I. Valderrama-Treviño, E.H. Contreras-Flores, et al., Int. J. Res. Med. Sci. 5 (2017) 4667. doi: 10.18203/2320-6012.ijrms20174914
W.S. Muhammad, C. Edward, Cancer Cell 16 (2010) 196–201.
Y.Q. Lu, Q. Han, X.R. Tan, et al., Trends MicroBiol. 32 (2024) 807–822. doi: 10.1016/j.tim.2024.01.007
G. Sambruni, A.D. Macandog, J. Wirbel, et al., Genome Med. 15 (2023) 32. doi: 10.1186/s13073-023-01180-9
J.H. Huang, J. Wang, X.Q. Chai, et al., MicroBiol. Spectr. 10 (2022) e00983-00922.
L.D. Wood, M.I. Canto, E.M. Jaffee, D.M. Simeone, Gastroenterology 163 (2022) 386–402. doi: 10.1053/j.gastro.2022.03.056
N.N. Liu, C.X. Yi, L.Q. Wei, et al., Cancer Cell 41 (2023) 1927–1944. doi: 10.1016/j.ccell.2023.08.012
A. Wong Rolle, Q. Dong, Y. Zhu, et al., J. Immunother. Cancer 10 (2022) 004698.
A.K. Markopoulos, Open Dent. J. 6 (2012) 126–130. doi: 10.2174/1874210601206010126
S. Kalaora, A. Nagler, D. Nejman, et al., Nature 592 (2021) 138–143. doi: 10.1038/s41586-021-03368-8
K. Vega Escobar, P.O. Armijos, T. Milman, et al., Am. J. Ophthalmol. Case Rep. 30 (2023) 101833. doi: 10.1016/j.ajoc.2023.101833
G. Zhu, H. Su, C.H. Johnson, et al., Eur. J. Cancer 151 (2021) 25–34. doi: 10.1016/j.ejca.2021.03.053
B. Aykut, S. Pushalkar, R. Chen, et al., Nature 574 (2019) 264–267. doi: 10.1038/s41586-019-1608-2
J. Sun, Q. Tang, S. Yu, et al., EBioMedicine 88 (2023) 88.
C. Neuzillet, M. Marchais, S. Vacher, et al., Sci. Rep. 11 (2021) 7870. doi: 10.1038/s41598-021-86816-9
B. h. Xian, J. m. Wei, G. b. Qiao, et al., J. Sun Yat-sen Univ. (Med. Sci. ) 43 (2022) 480–487.
K. Takabe, C. Xuan, J.M. Shamonki, et al., PLoS One 9 (2014) e83744. doi: 10.1371/journal.pone.0083744
M. Vyara, F. Jessica, B. Riyue, et al., Science 359 (2018) 104–108. doi: 10.1126/science.aao3290
G.D. Sepich Poore, H. Carter, R. Knight, Cancer Cell 39 (2021) 601–603. doi: 10.1016/j.ccell.2021.04.008
C. Xue, J. Jia, X. Gu, et al., Signal Transduct. Target. Ther. 7 (2022) 335. doi: 10.1038/s41392-022-01159-9
K.L. Greathouse, J.R. White, A.J. Vargas, et al., Genome Biol. 19 (2018) 123. doi: 10.1186/s13059-018-1501-6
J. Massagué, A.C. Obenauf, Nature 529 (2016) 298–306. doi: 10.1038/nature17038
A. Fu, B. Yao, T. Dong, S. Cai, Trends Cell Biol. 33 (2023) 583–593. doi: 10.1016/j.tcb.2022.11.007
F. Aikun, Y. Bingqing, D. Tingting, C. Shang, Life Medicine 1 (2022) 61–63. doi: 10.1093/lifemedi/lnac009
G. Wang, X. He, Q. Wang, Biochim. Biophys. Acta Rev. Cancer 1878 (2023) 188846. doi: 10.1016/j.bbcan.2022.188846
S. Guo, J. Chen, F. Chen, et al., Gut 70 (2021) 1507–1519. doi: 10.1136/gutjnl-2020-321187
Y. Zhang, L. Zhang, S. Zheng, et al., Gut Microbes 14 (2022) 2038852. doi: 10.1080/19490976.2022.2038852
A. Bertocchi, S. Carloni, P.S. Ravenda, et al., Cancer Cell 39 (2021) 708–724. doi: 10.1016/j.ccell.2021.03.004
L. Parhi, T. Alon-Maimon, A. Sol, et al., Nat. Commun. 11 (2020) 3259. doi: 10.1038/s41467-020-16967-2
S. Parida, S. Wu, S. Siddharth, et al., Cancer Discov. 11 (2021) 1138–1157. doi: 10.1158/2159-8290.cd-20-0537
A. Sevcikova, N. Izoldova, V. Stevurkova, et al., Int. J. Mol Sci. 23 (2022) 488. doi: 10.3390/ijms23010488
A. Tijeras Raballand, M. Hilmi, L. Astorgues-Xerri, et al., Clin. Res. Hepatol Gas 45 (2021) 101589.
S. Lu, J. Lin, J. Jin, et al., Expert Opin. Drug Discov. 17 (2022) 1407–1423. doi: 10.1080/17460441.2023.2157402
S.W. Guan, Q. Lin, H.B. Yu, World J. Gastrointest. Oncol. 15 (2023) 713–730. doi: 10.4251/wjgo.v15.i5.713
X. Huang, C. Chen, W. Xie, et al., Int. J. Radiat. Oncol. Biol. Phys. 117 (2023) 1255–1269. doi: 10.1016/j.ijrobp.2023.06.2515
E. Riquelme, A. Maitra, F. McAllister, Cancer Discov. 8 (2018) 386–388. doi: 10.1158/2159-8290.CD-18-0123
K.D. LaCourse, M. Zepeda-Rivera, A.G. Kempchinsky, et al., Cell Rep. 41 (2022) 111625. doi: 10.1016/j.celrep.2022.111625
M. Hamada, H. Inaba, K. Nishiyama, et al., Int. J. Mol. Sci. 24 (2023) 15456. doi: 10.3390/ijms242015456
L.E. Colbert, M.B. El Alam, R. Wang, et al., Cancer Cell 41 (2023) 1945–1962. doi: 10.1016/j.ccell.2023.09.012
M. Zhang, Y. Zhang, Y. Han, et al., Transl. Cancer Res. 11 (2022) 426–434. doi: 10.21037/tcr-22-92
L. Yuan, P. Yang, G. Wei, et al., Commun. Biol. 5 (2022) 864. doi: 10.1038/s42003-022-03814-x
R.E. Eckart, E.A. Shry, A.P. Burke, et al., J. Am. Coll. Cardiol. 58 (2011) 1254–1261. doi: 10.1016/j.jacc.2011.01.049
C. Urbaniak, G.B. Gloor, M. Brackstone, et al., Appl. Environ. Microbiol. 82 (2016) 5039–5048. doi: 10.1128/AEM.01235-16
T. Dong, A. Fu, S. Cai, STAR Protoc. 3 (2022) 101624. doi: 10.1016/j.xpro.2022.101624
A. Tzeng, N. Sangwan, M. Jia, et al., Genome Med. 13 (2021) 60. doi: 10.1186/s13073-021-00874-2
H. Qiao, X.R. Tan, H. Li, et al., JAMA Oncol. 8 (2022) 1301. doi: 10.1001/jamaoncol.2022.2810
M. Hilmi, C. Neuzillet, J.H. Lefèvre, et al., Cancers 14 (2022) 1606. doi: 10.3390/cancers14071606
D. Sheng, K. Yue, H. Li, et al., MicroBiol. Spectr. 11 (2023) e03549-03522. doi: 10.1128/spectrum.03549-22
Y. Li, D. Zhang, M. Wang, et al., iMeta 2 (2023) e102. doi: 10.1002/imt2.102
Y. Huang, N. Zhu, X. Zheng, et al., Front. Immunol. 13 (2022) 785422. doi: 10.3389/fimmu.2022.785422
J.L. Alexander, J.M. Posma, A. Scott, et al., Microbiome 11 (2023) 100. doi: 10.1186/s40168-023-01518-w
A.S. Campbell, D. Albo, T.F. Kimsey, et al., J. Surg. Res. 123 (2005) 96–101. doi: 10.1016/j.jss.2004.07.013
D. Unutmaz, K.J. Chen, S.Z. Lin, et al., PLoS One 6 (2011) e24671. doi: 10.1371/journal.pone.0024671
Y. Tomohide, Y. Xuexian O, C. Yeonseok, et al., J. Immunol. 181 (2009) 8391–8401.
M. Salvucci, N. Crawford, K. Stott, et al., Gut 71 (2021) 1600–1612.
D. Mouradov, P. Greenfield, S. Li, et al., Gastroenterology 165 (2023) 104–120. doi: 10.1053/j.gastro.2023.03.205
S. Banerjee, Z. Wei, T. Tian, et al., Cell Death Dis. 12 (2021) 831. doi: 10.1038/s41419-021-04092-x
R. Brett M, P. Jennifer B, S. Thomas A, Cancer Biol. Med. 14 (2017) 9–32. doi: 10.20892/j.issn.2095-3941.2016.0084
C. Stewart, C. Ralyea, S. Lockwood, Semin. Oncol. Nurs. 35 (2019) 151–156. doi: 10.1016/j.soncn.2019.02.001
W. Cheng, L. He, W. Ren, et al., Nano TransMed 2 (2023) 100008. doi: 10.1016/j.ntm.2023.100008
J. y. Zhang, M. Shen, Clin. Immunol. 17 (2023) 68–73.
V. Pareek, S. Devineau, S.K. Sivasankaran, et al., Front. Microbiol. 12 (2021) 638640. doi: 10.3389/fmicb.2021.638640
D.J. Dwyer, M.A. Kohanski, B. Hayete, J.J. Collins, Mol. Syst. Biol. 3 (2007) 91. doi: 10.1038/msb4100135
G.D. Wright, Cell 130 (2007) 781–783. doi: 10.1016/j.cell.2007.08.023
Y. Luo, Y. Song, Int. J. Mol. Sci. 22 (2021) 11401. doi: 10.3390/ijms222111401
W.Y. Cheng, C.Y. Wu, J. Yu, Gut 69 (2020) 1867–1876. doi: 10.1136/gutjnl-2020-321153
C. Moser, C.J. Lerche, K. Thomsen, et al., APMIS 127 (2019) 361–371. doi: 10.1111/apm.12951
S.S. Jiang, Y.L. Xie, X.Y. Xiao, et al., Cell Host Microbe 31 (2023) 781–797. doi: 10.3390/f14040781
Z. Cao, J. Liu, Acc. Mater. Res. 5 (2024) 872–883. doi: 10.1021/accountsmr.4c00116
J. Yu, B. Zhou, S. Zhang, et al., Nat. Commun. 13 (2022) 7903. doi: 10.1038/s41467-022-35580-z
L. Wang, Z. Cao, M. Zhang, et al., Adv. Mater. 34 (2021) 2106.
Q. Han, F. Yang, M. Chen, et al., Adv. Sci. 11 (2024) 2407402. doi: 10.1002/advs.202407402
K.F. Xu, S.Y. Wu, Z. Wang, et al., Nat. Commun. 15 (2024) 5147. doi: 10.1038/s41467-024-49156-6
S. Chowdhury, S. Castro, C. Coker, et al., Nat. Med. 25 (2019) 1057–1063. doi: 10.1038/s41591-019-0498-z
X. Zhang, D. Yu, D. Wu, et al., Cell Host Microbe 31 (2023) 418–432. doi: 10.1007/978-3-031-46674-8_29
H. Wu, X. Leng, Q. Liu, et al., Cancer Res. 83 (2023) 3131–3144. doi: 10.1158/0008-5472.can-22-2593
Y. He, L. Fu, Y. Li, et al., Cell Metab. 33 (2021) 988–1000. doi: 10.1016/j.cmet.2021.03.002
P. Louis, H.J. Flint, FEMS MicroBiol. Lett. 294 (2009) 1–8. doi: 10.1111/j.1574-6968.2009.01514.x
M.K. Stoeva, J. Garcia-So, N. Justice, et al., Gut Microbes 13 (2021) 1907272. doi: 10.1080/19490976.2021.1907272
M.J. Bender, A.C. McPherson, C.M. Phelps, et al., Cell 186 (2023) 1846–1862. doi: 10.1016/j.cell.2023.03.011
H. Wang, X. Rong, G. Zhao, et al., Cell Metab. 34 (2022) 581–594. doi: 10.1016/j.cmet.2022.02.010
J. Tintelnot, Y. Xu, T.R. Lesker, et al., Nature 615 (2023) 168–174. doi: 10.1038/s41586-023-05728-y
J. Cong, P. Liu, Z. Han, et al., Immunity 57 (2024) 876–889. doi: 10.1016/j.immuni.2024.02.014
R. Dey, S. Mukherjee, S. Barman, J. Haldar, Macromol. Biosci. 21 (2021) 2100182. doi: 10.1002/mabi.202100182
J. Zou, X. Xing, C. Teng, et al., Acta Pharm. Sin. B 14 (2024) 4509–4525. doi: 10.1016/j.apsb.2024.08.014
W. Chen, K. Shi, Y. Yu, et al., Chin. Chem. Lett. 35 (2024) 109159. doi: 10.1016/j.cclet.2023.109159
L. Yin, X. Jiang, M. Wang, et al., Chin. Chem. Lett. (2024) 110224. doi: 10.1016/j.cclet.2024.110224
S. Kwon, H. ThiThu Ngo, J. Son, et al., Nat. Rev. Clin. Oncol. 21 (2024) 569–589. doi: 10.1038/s41571-024-00908-9
W. Eisenreich, T. Rudel, J. Heesemann, W. Goebel, Front. Cell Infect Microbiol. 12 (2022) 900848. doi: 10.3389/fcimb.2022.900848
M. Zoulikha, Z. Chen, J. Wu, W. He, Chin. Chem. Lett. 36 (2025) 110225. doi: 10.1016/j.cclet.2024.110225
L. Chen, R. Zhao, J. Shen, et al., Adv. Mater. 35 (2023) 2306281. doi: 10.1002/adma.202306281
X. Liu, M. Sun, F. Pu, et al., J. Am. Chem. Soc. 145 (2023) 26296–26307. doi: 10.1021/jacs.3c09472
Q. Qiu, D. Lu, G. Liu, et al., Bioconjug. Chem. 33 (2022) 1944–1952. doi: 10.1021/acs.bioconjchem.2c00407
X. Li, Y. Ma, Y. Xin, et al., ACS Appl. Mater. Interfaces. 15 (2023) 14164–14172.
Z. Ma, H. Wang, Z. Shi, et al., ACS Nano 17 (2023) 5740–5756. doi: 10.1021/acsnano.2c12165
R. Liu, H. Yang, S. Qu, et al., Aggreg 5 (2023) e423.
W. Song, K. Tiruthani, Y. Wang, et al., Adv. Mater. 30 (2018) e1805007. doi: 10.1002/adma.201805007
F. Kong, C. Fang, Y. Zhang, et al., Adv. Sci. 9 (2022) 2105523. doi: 10.1002/advs.202105523
X. Kang, F. Bu, W. Feng, et al., Adv. Mater. 34 (2022) 2206765. doi: 10.1002/adma.202206765
H. Zhou, D. Tang, X. Kang, et al., Adv. Sci. 9 (2022) 2200732. doi: 10.1002/advs.202200732
Q. Chen, Y. Zhu, K. Wang, et al., Nano Today 52 (2023) 101994. doi: 10.1016/j.nantod.2023.101994
X. Qu, F. Yin, M. Pei, et al., ACS Nano 17 (2023) 11466–11480. doi: 10.1021/acsnano.3c01308
X. Wang, Q. Chen, Y. Zhu, et al., Signal Transduct. Target. Ther. 8 (2023) 227. doi: 10.1038/s41392-023-01435-2
D.W. Zheng, X. Dong, P. Pan, et al., Nat. Biomed. Eng. 3 (2019) 717–728. doi: 10.1038/s41551-019-0423-2
T. Hillman, Nano Particle 1 (2019) 1.
W. Song, D. Zheng, S. Zeng, et al., ACS Nano 16 (2022) 17402–17413. doi: 10.1021/acsnano.2c08555
S. Geng, P. Guo, X. Li, et al., ACS Nano 18 (2024) 8971–8987. doi: 10.1021/acsnano.3c12687
Z. Han, Q. Chen, D. Zheng, et al., Adv. Mater. 35 (2023) 2302551. doi: 10.1002/adma.202302551
C. Wang, L. Zhong, J. Xu, et al., Nat. BioMed. Eng. 8 (2024) 561–578. doi: 10.1038/s41551-024-01191-w
Y. Ma, S. Liu, Q. Zhou, et al., Chin. J. Nat. Med. 22 (2024) 699–710.

Figure 2 Killing intratumoural bacteria by non-nanodelivery systems. (a) Microbial metabolite trimethylamine N-oxide enhances CD8+ T cell-mediated antitumor immunity in triple-negative breast cancer (TNBC) by inducing pyroptosis in tumor cells, revealing insights into microbiota-metaboliteimmune crosstalk. Copied with permission [111]. Copyright 2022, Elsevier. (b) Deoxycholic acid suppresses cytotoxic CD8+ T cell effector functions by impairing intracellular calcium accumulation, subsequently promoting colorectal tumor growth. Copied with permission [113]. Copyright 2024, Elsevier.

Figure 3 Killing intratumoural bacteria by nanodelivery systems. (a) Negatively charged HA as the shell of pPCPs formed through electrostatic interaction to form the core–shell nanoparticles pPCP-NPs. After intravenous injection, pPCP-NPs were effectively accumulated in the infected site in the mouse model. HA was degradable under the HAase and then pPCPs were released. pPCPs then anchored to bacterial membrane by electrostatic interaction. Under 1064 nm (NIR-Ⅱ) laser irradiation, pPCPs generated heat, resulting in bacterial cell death. Copied with permission [129]. Copyright 2023, American Chemical Society. (b) Schematic description of the Au@BSA-CuPpIX synthesizing process. (c) Au@BSA-CuPpIX reduces the level of apoptosisinhibiting proteins by inhibiting F. nucleatum to advance the efficacy of sound dynamic therapy (SDT) for orthotopic CRC with lung metastasis restriction. (d) Au@BSA-CuPpIX effectively reduces the phototoxicity of metalloporphyrin accumulated in the skin during tumor treatment, preventing severe inflammation and damage to the skin. Copied with permission [132]. Copyright 2023, American Chemical Society.

 扫一扫看文章
扫一扫看文章

扫一扫关注我们
 DownLoad:
DownLoad:

 下载:
下载:
 下载:
下载: